
Functional Reconstruction of Denervated Muscle by Xenotransplantation of Neural Cells from Porcine to Rat

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
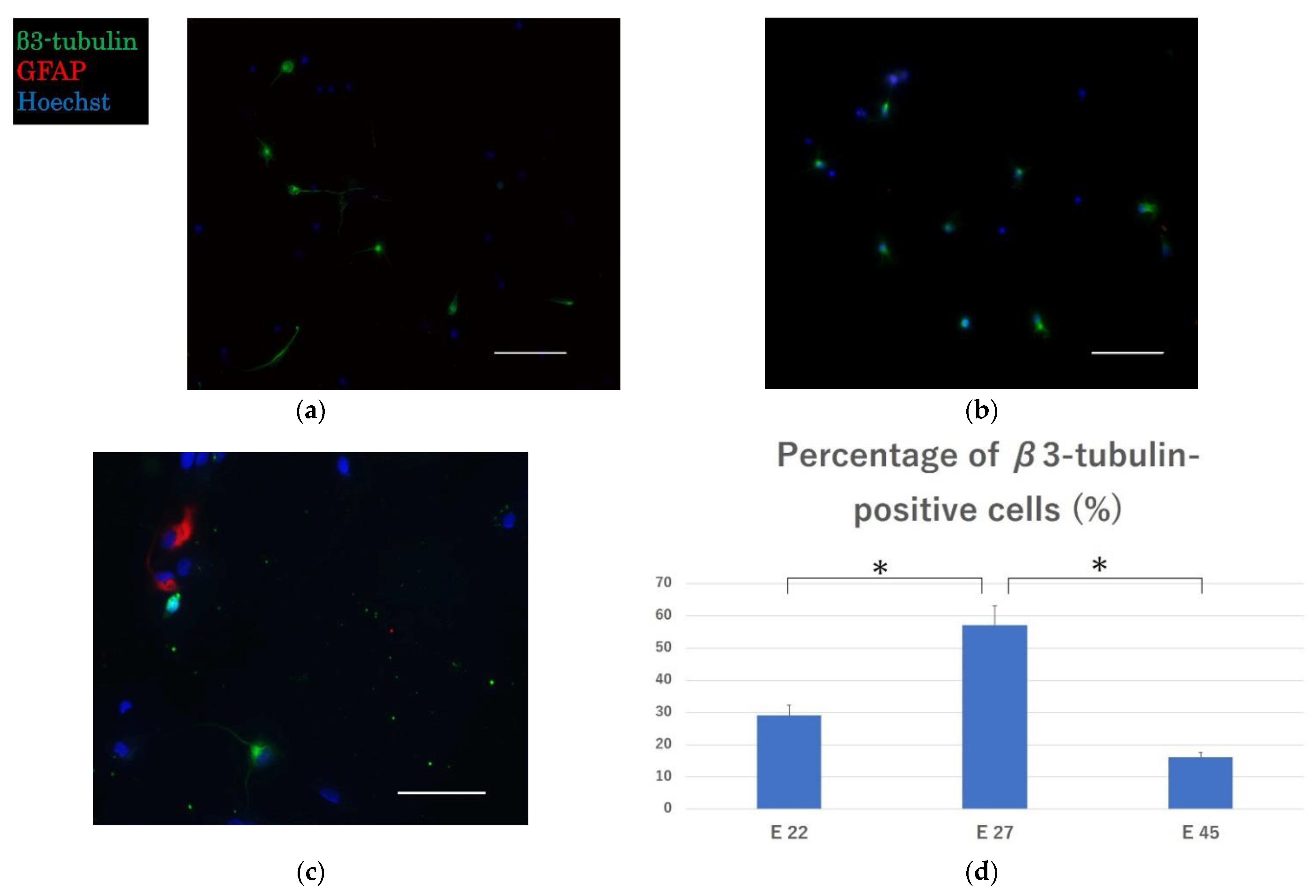
2.1. Effective Engraftment and Neurogenesis Post-Transplantation
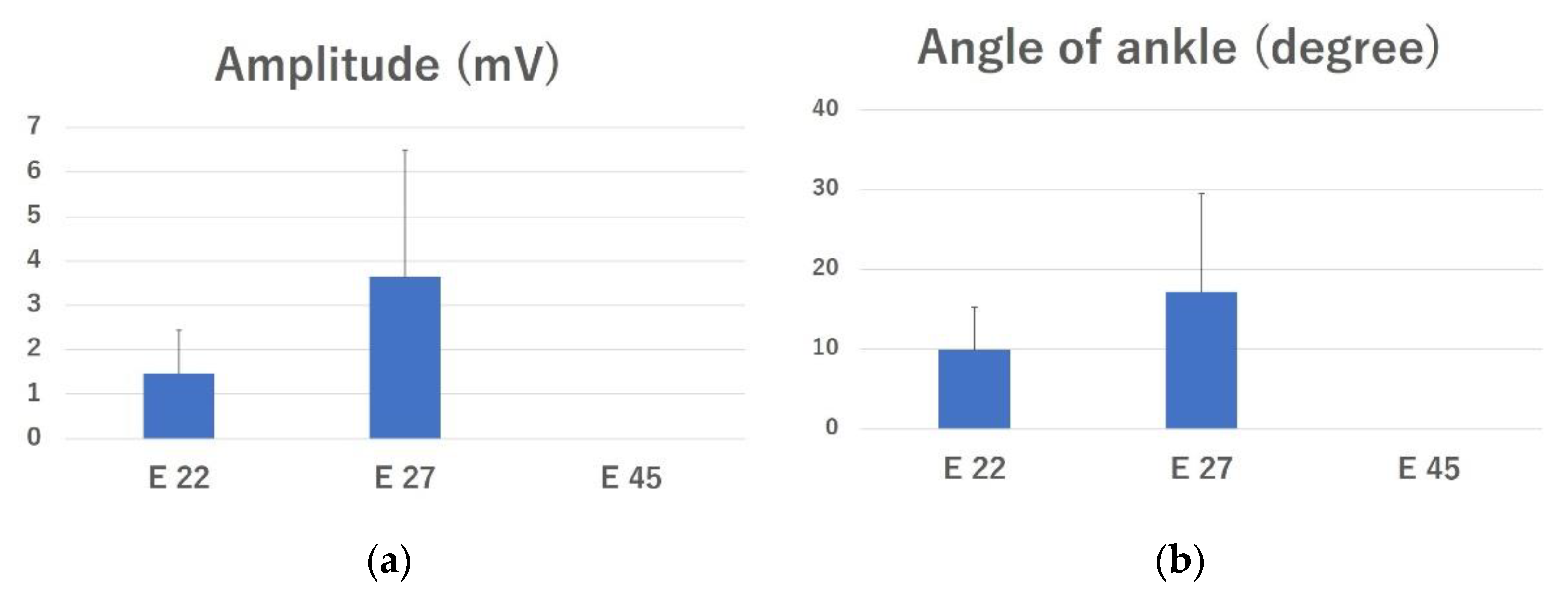
2.2. Electrophysiological Evaluation
2.3. Tissue Analysis
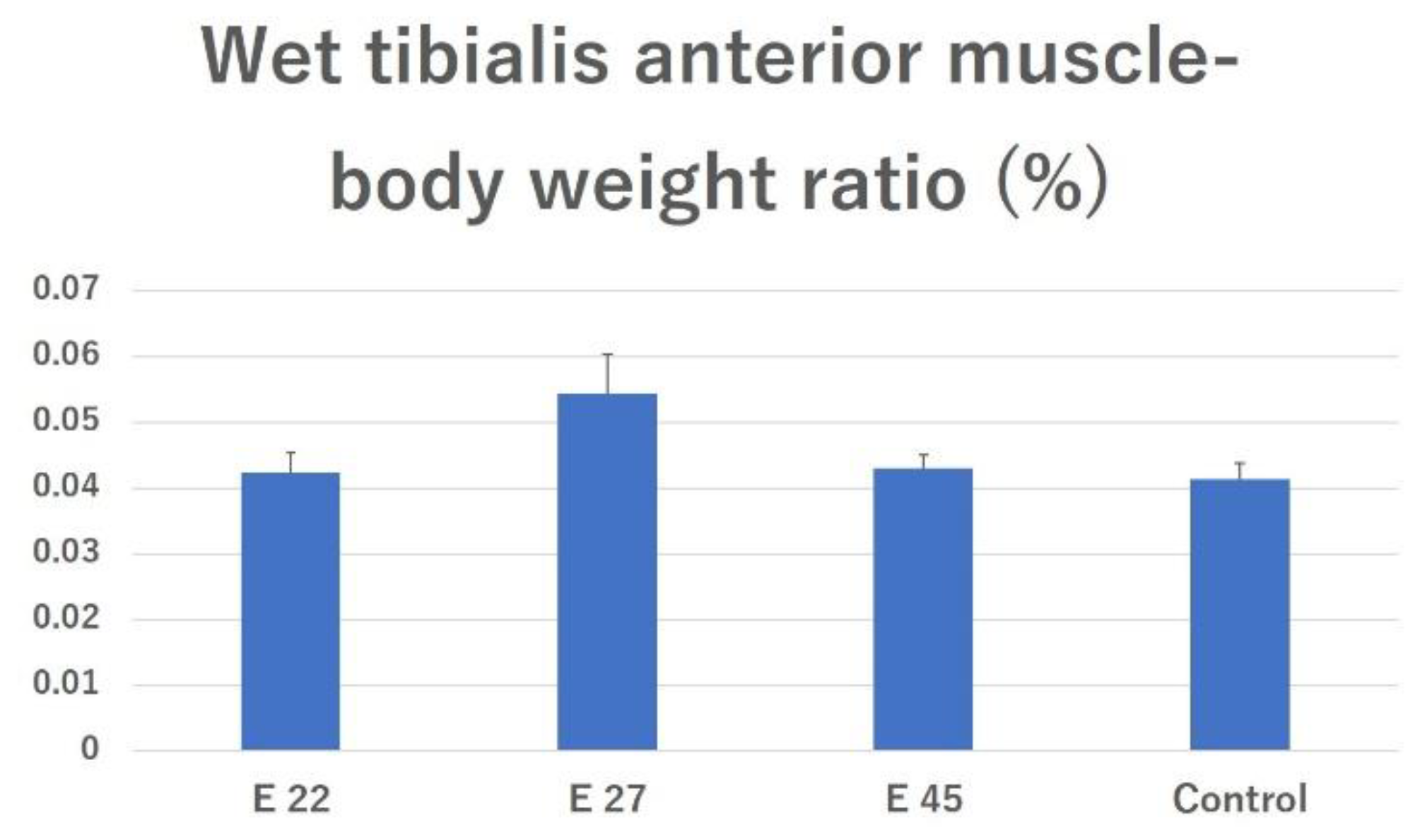
2.3.1. Effects of Preventing Muscle Atrophy
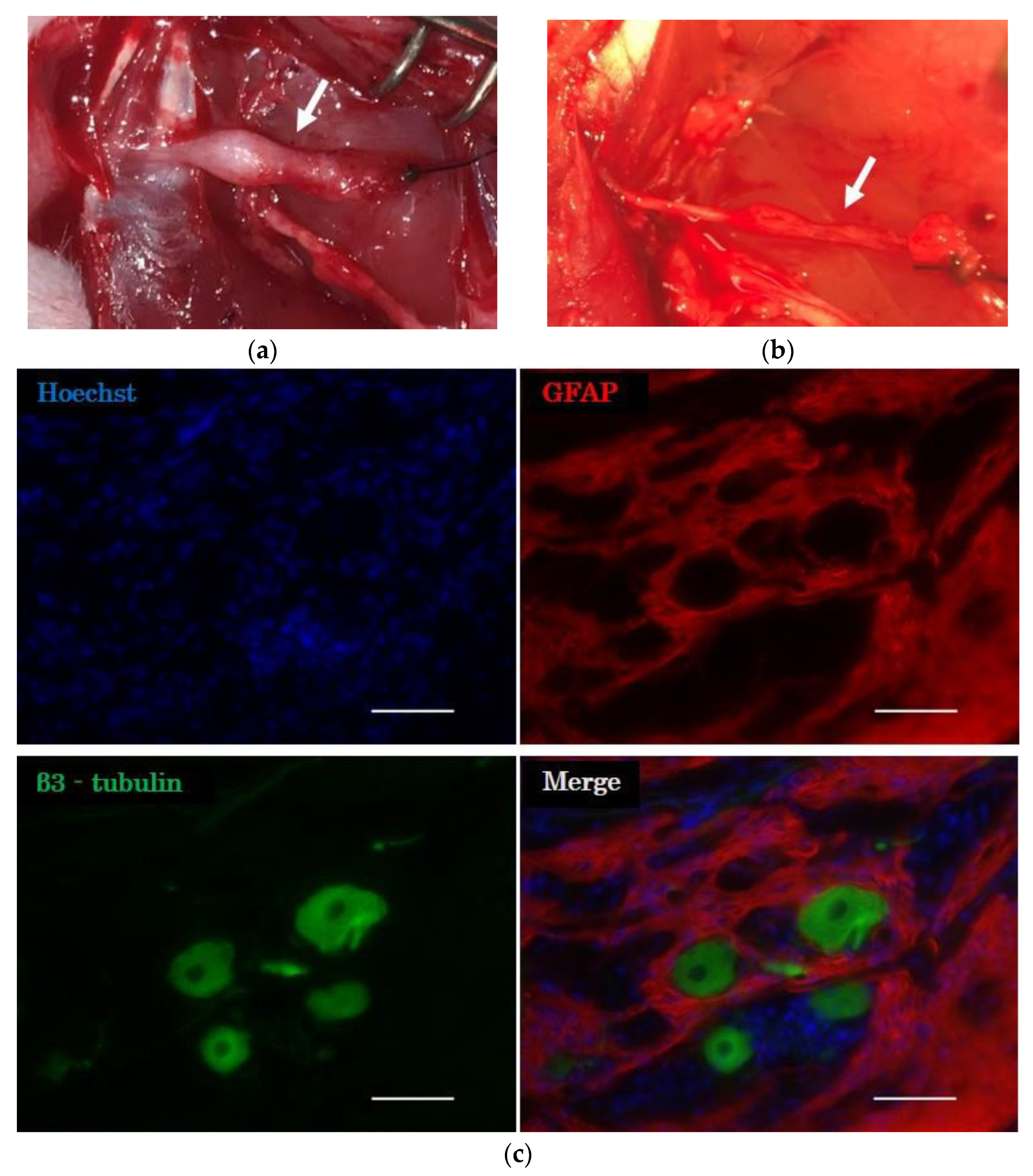
2.3.2. Histological Evaluation of the Distal Nerve Stump
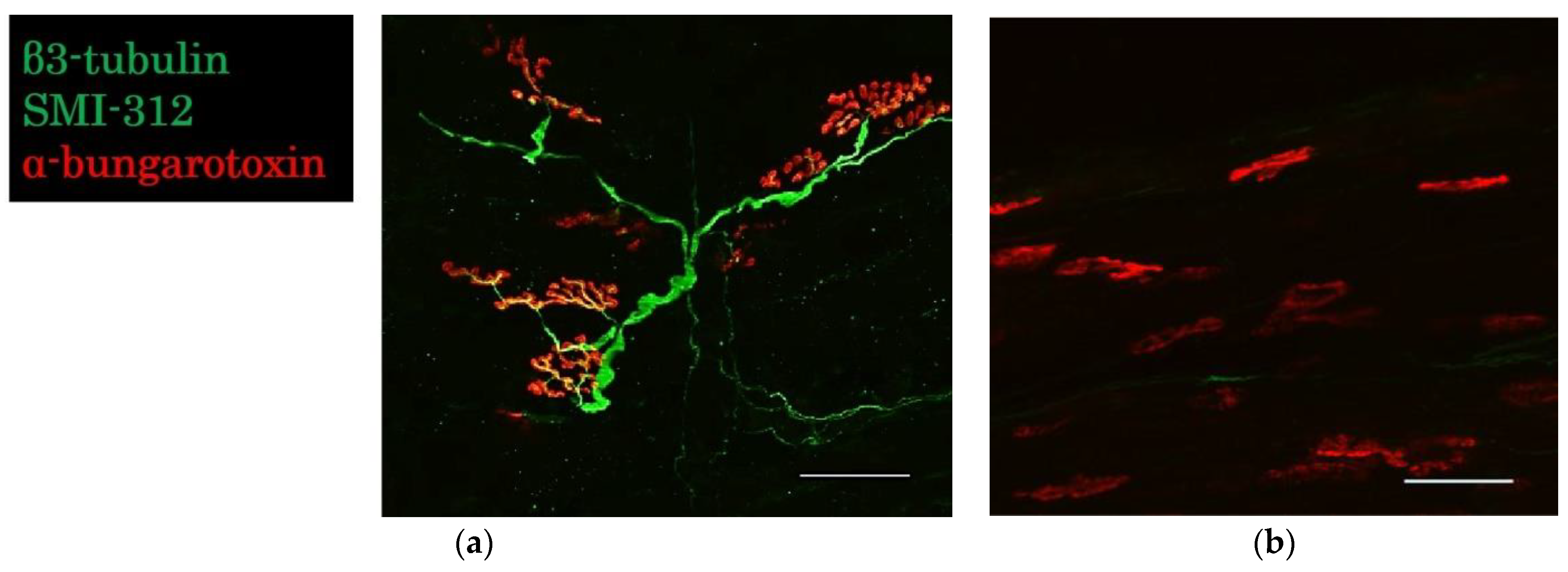
2.3.3. Evaluation of the Neuromuscular Junction
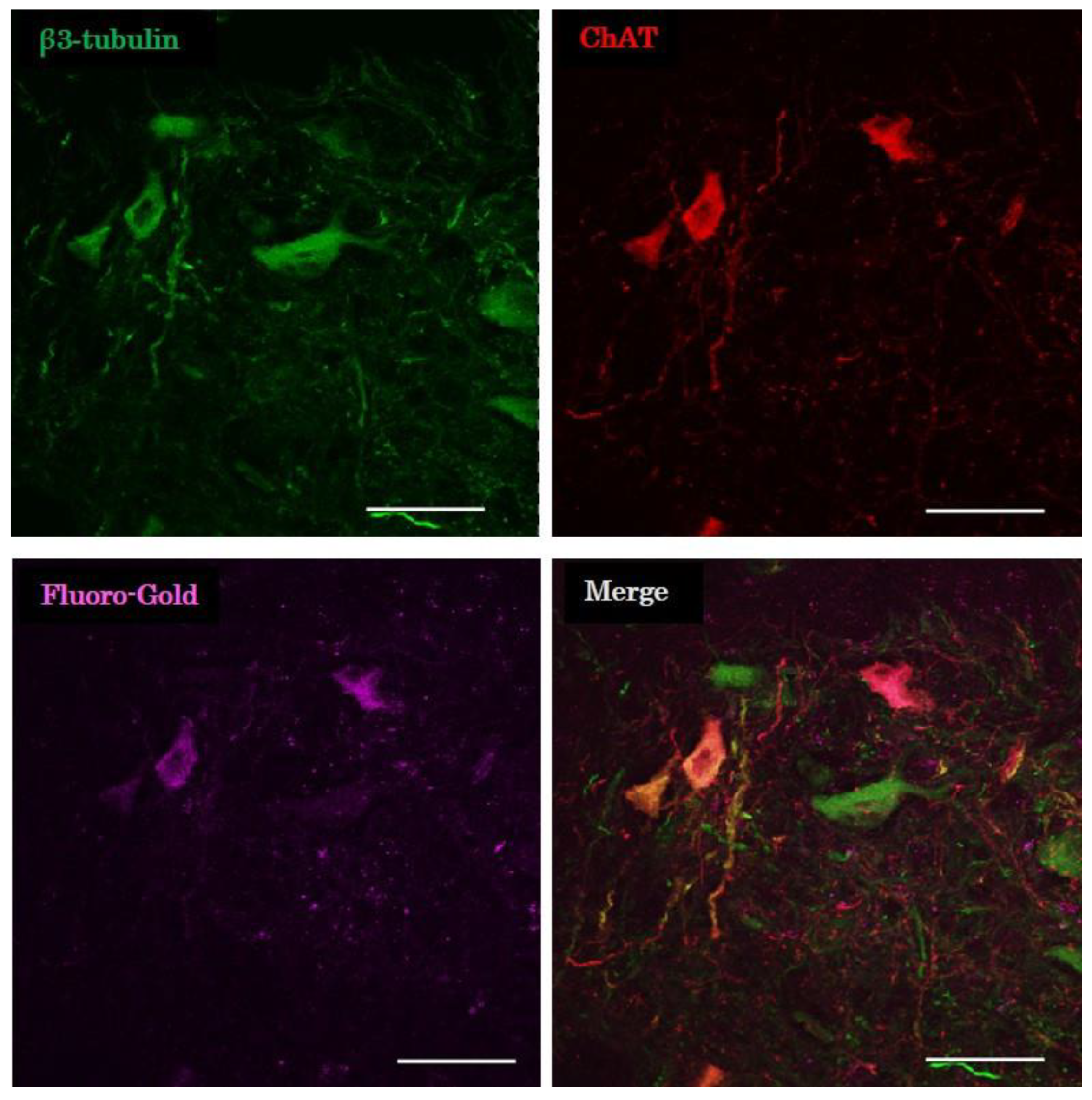
2.3.4. Evaluation of Retrograde Tracing
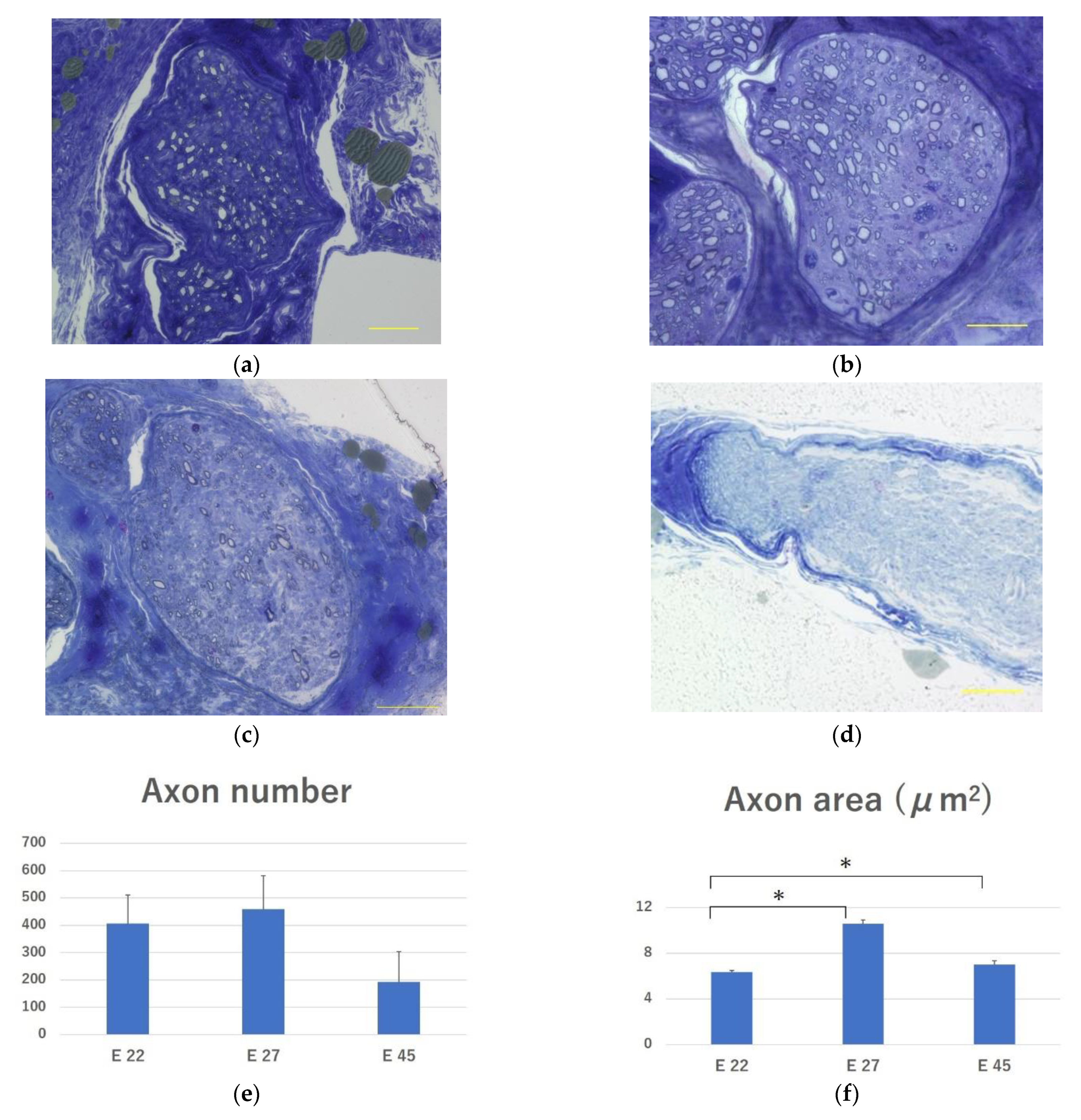
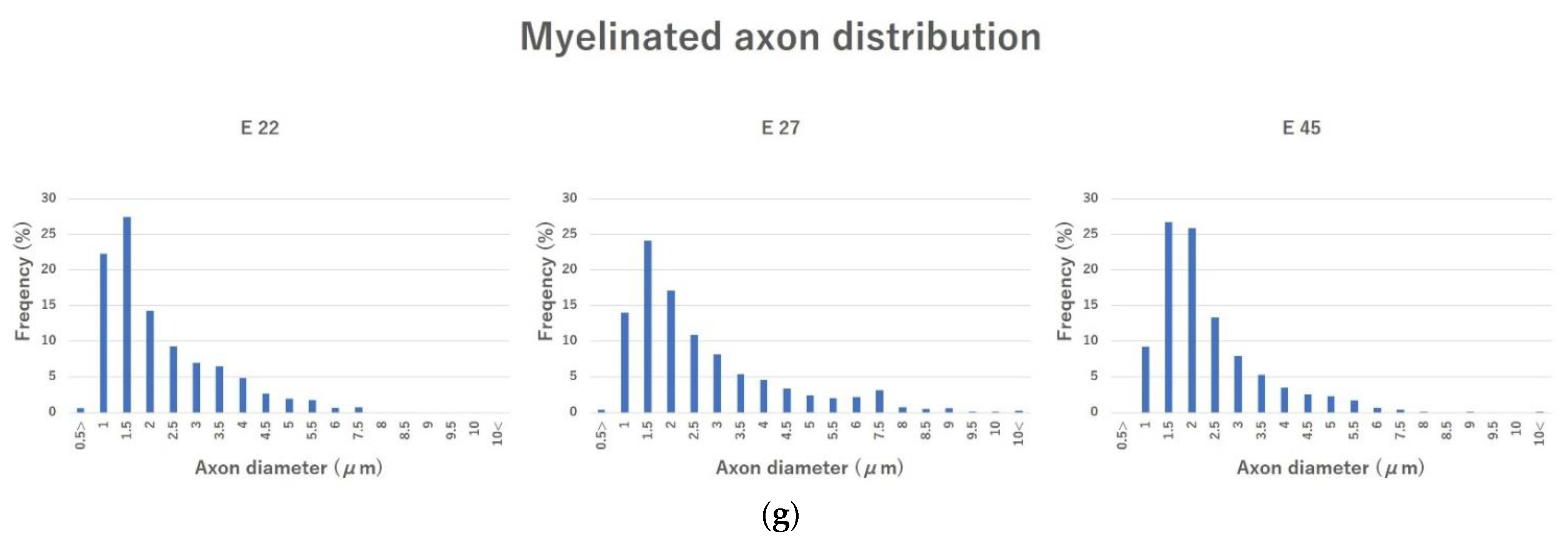
2.3.5. Evaluation of Regenerated Myelinated Axons
3. Discussion
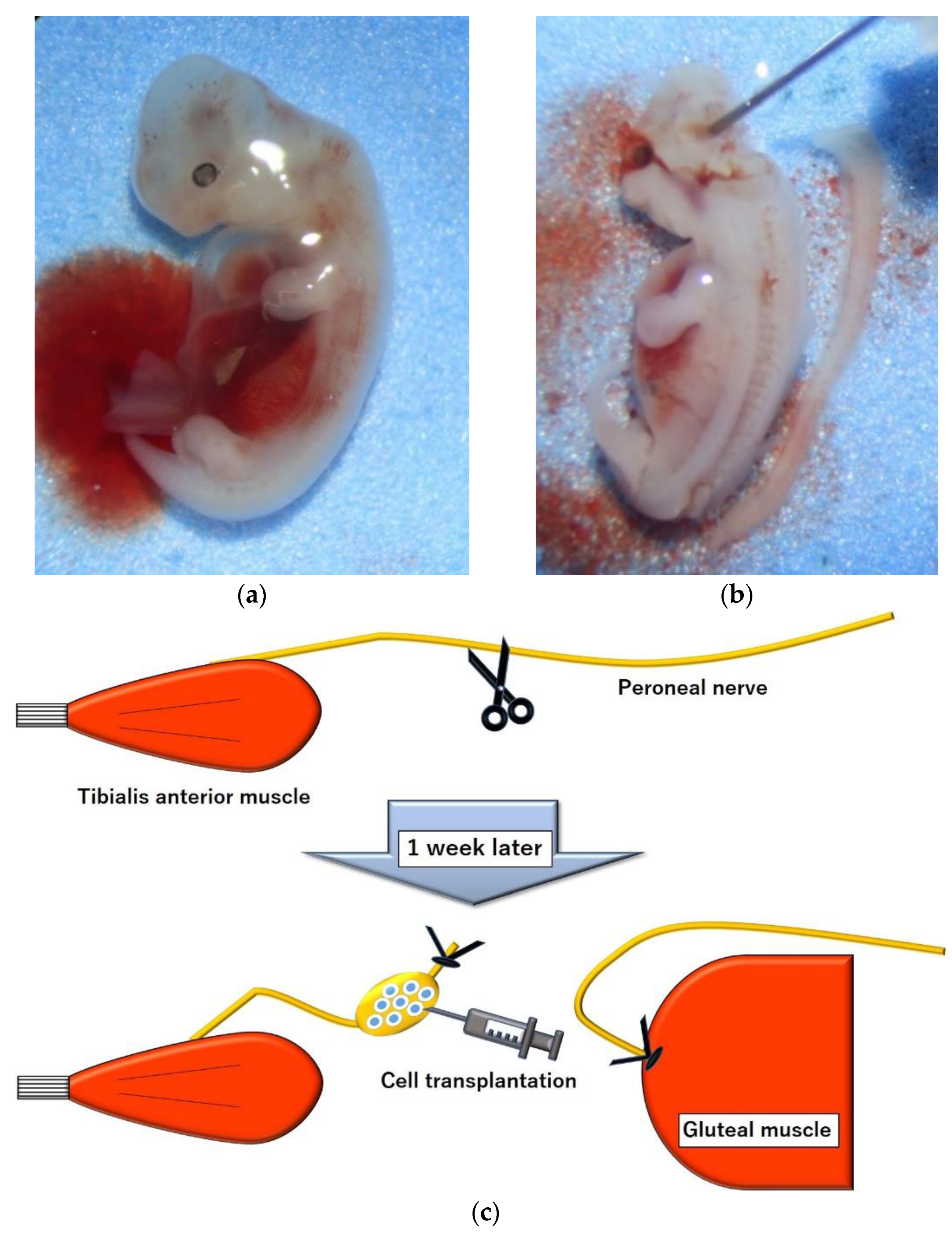
4. Materials and Methods
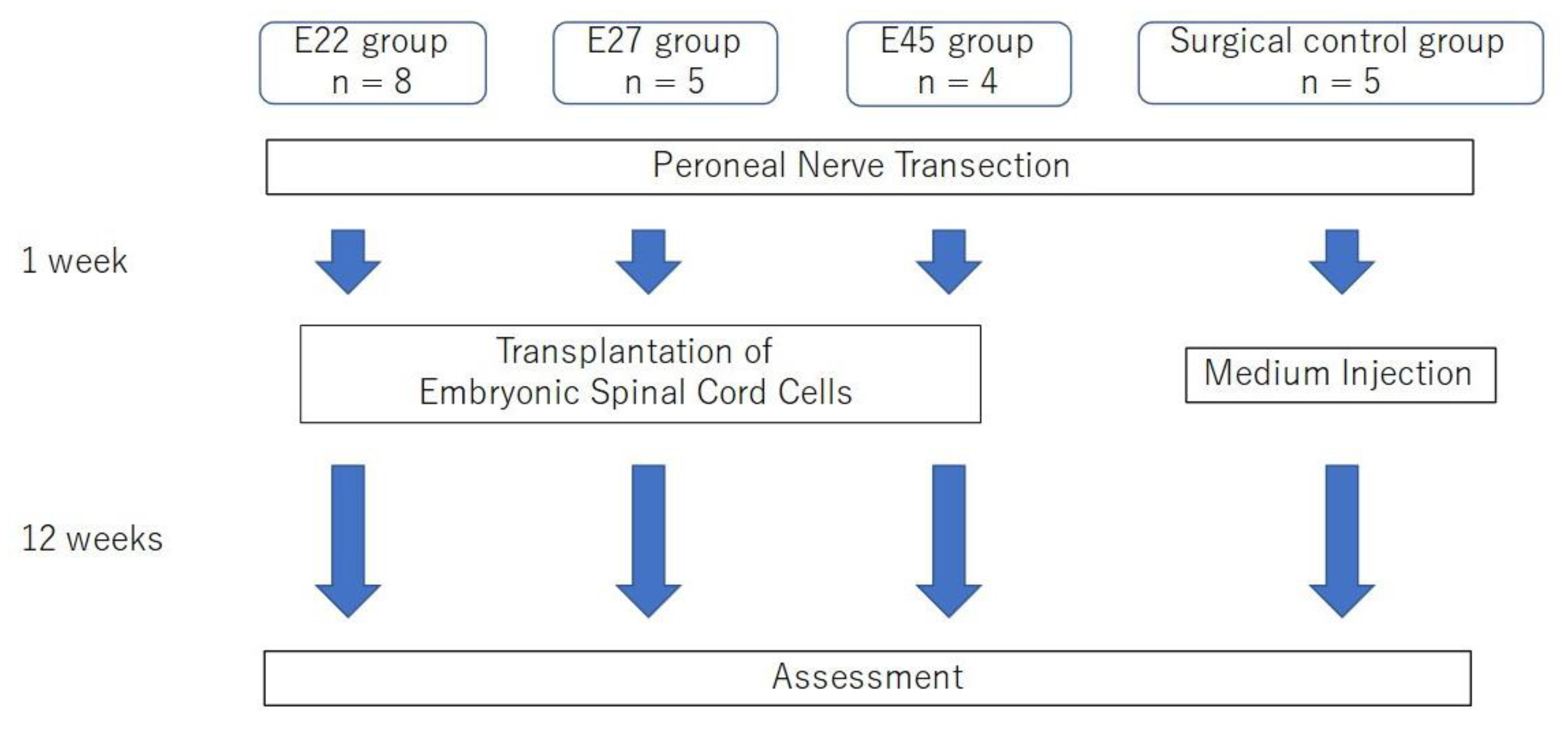
4.1. Animals
4.2. Cell Harvest, Preparation and Transplantation
4.3. Cell Culture
4.4. Electrophysiological Evaluation
4.5. Animal Sacrifice
4.6. Tissue Analysis
4.6.1. Evaluation of the Nodule Structure in the Distal Nerve Stump
4.6.2. Evaluation of the Neuromuscular Junction
4.6.3. Evaluation of Retrograde Tracing
4.6.4. Evaluation of Regenerated Myelinated Axons
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Assinck, P.; Duncan, G.J.; Hilton, B.J.; Plemel, J.R.; Tetzlaff, W. Cell Transplantation Therapy for Spinal Cord Injury. Nat. Neurosci. 2017, 20, 637–647. [Google Scholar] [CrossRef]
- Xu, X.; Shen, D.; Gao, Y.; Zhou, Q.; Ni, Y.; Meng, H.; Shi, H.; Le, W.; Chen, S.; Chen, S. A Perspective on Therapies for Amyotrophic Lateral Sclerosis: Can Disease Progression Be Curbed? Transl. Neurodegener. 2021, 10, 29. [Google Scholar] [CrossRef]
- Goutman, S.A.; Savelieff, M.G.; Sakowski, S.A.; Feldman, E.L. Stem Cell Treatments for Amyotrophic Lateral Sclerosis: A Critical Overview of Early Phase Trials. Expert Opin. Investig. Drugs 2019, 28, 525–543. [Google Scholar] [CrossRef]
- Gordon, T.; Tyreman, N.; Raji, M.A. The Basis for Diminished Functional Recovery after Delayed Peripheral Nerve Repair. J. Neurosci. 2011, 31, 5325–5334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarhane, K.A.; Slavin, B.R.; Hricz, N.; Malapati, H.; Guo, Y.N.; Grzelak, M.; Chang, I.A.; Shappell, H.; von Guionneau, N.; Wong, A.L.; et al. Defining the Relative Impact of Muscle versus Schwann Cell Denervation on Functional Recovery after Delayed Nerve Repair. Exp. Neurol. 2021, 339, 113650. [Google Scholar] [CrossRef] [PubMed]
- Vansteensel, M.J.; Pels, E.G.M.; Bleichner, M.G.; Branco, M.P.; Denison, T.; Freudenburg, Z.V.; Gosselaar, P.; Leinders, S.; Ottens, T.H.; Van Den Boom, M.A.; et al. Fully Implanted Brain-Computer Interface in a Locked-In Patient with ALS. N. Engl. J. Med. 2016, 375, 2060–2066. [Google Scholar] [CrossRef]
- Slutzky, M.W. Brain-Machine Interfaces: Powerful Tools for Clinical Treatment and Neuroscientific Investigations. Neuroscientist 2019, 25, 139–154. [Google Scholar] [CrossRef] [PubMed]
- Willett, F.R.; Avansino, D.T.; Hochberg, L.R.; Henderson, J.M.; Shenoy, K.V. High-Performance Brain-to-Text Communication via Handwriting. Nature 2021, 593, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Lewis, M.; Rushanan, S. The Role of Physical Therapy and Occupational Therapy in the Treatment of Amyotrophic Lateral Sclerosis. NeuroRehabilitation 2007, 22, 451–461. [Google Scholar] [CrossRef]
- Gonzalez-Bermejo, J.; Morélot-Panzini, C.; Tanguy, M.L.; Meininger, V.; Pradat, P.F.; Lenglet, T.; Bruneteau, G.; Forestier, N.L.; Couratier, P.; Guy, N.; et al. Early Diaphragm Pacing in Patients with Amyotrophic Lateral Sclerosis (RespiStimALS): A Randomised Controlled Triple-Blind Trial. Lancet Neurol. 2016, 15, 1217–1227. [Google Scholar] [CrossRef]
- Erb, D.E.; Mora, R.J.; Bunge, R.P. Reinnervation of Adult Rat Gastrocnemius Muscle by Embryonic Motoneurons Transplanted into the Axotomized Tibial Nerve. Exp. Neurol. 1993, 124, 372–376. [Google Scholar] [CrossRef] [PubMed]
- Grumbles, R.M.; Wood, P.; Rudinsky, M.; Gomez, A.M.; Thomas, C.K. Muscle Reinnervation with Delayed or Immediate Transplant of Embryonic Ventral Spinal Cord Cells into Adult Rat Peripheral Nerve. Cell Transplant. 2002, 11, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Bryson, J.B.; Machado, C.B.; Crossley, M.; Stevenson, D.; Bros-Facer, V.; Burrone, J.; Greensmith, L.; Lieberam, I. Optical Control of Muscle Function by Transplantation of Stem Cell-Derived Motor Neurons in Mice. Science 2014, 344, 94–97. [Google Scholar] [CrossRef] [Green Version]
- Kurimoto, S.; Kato, S.; Nakano, T.; Yamamoto, M.; Takanobu, N.; Hirata, H. Transplantation of Embryonic Motor Neurons into Peripheral Nerve Combined with Functional Electrical Stimulation Restores Functional Muscle Activity in the Rat Sciatic Nerve Transection Model. J. Tissue Eng. Regen. Med. 2016, 10, E477–E484. [Google Scholar] [CrossRef] [PubMed]
- Asano, K.; Nakano, T.; Tokutake, K.; Ishii, H.; Nishizuka, T.; Iwatsuki, K.; Onishi, T.; Kurimoto, S.; Yamamoto, M.; Tatebe, M.; et al. Muscle Spindle Reinnervation Using Transplanted Embryonic Dorsal Root Ganglion Cells after Peripheral Nerve Transection in Rats. Cell Prolif. 2019, 52, e12660. [Google Scholar] [CrossRef] [PubMed]
- Asano, K.; Nakano, T.; Tokutake, K.; Ishii, H.; Nishizuka, T.; Iwatsuki, K.; Onishi, T.; Kurimoto, S.; Yamamoto, M.; Tatebe, M.; et al. Innervation of Meissner’s Corpuscles And Merkel—Cells By Transplantation of Embryonic Dorsal Root Ganglion Cells after Peripheral Nerve Section in Rats. J. Tissue Eng. Regen. Med. 2021, 15, 586–595. [Google Scholar] [CrossRef]
- Yohn, D.C.; Miles, G.B.; Rafuse, V.F.; Brownstone, R.M. Transplanted mouse embryonic stem-cell-derived motoneurons form functional motor units and reduce muscle atrophy. J. Neurosci. 2008, 28, 12409–12418. [Google Scholar] [CrossRef] [Green Version]
- Toma, J.S.; Shettar, B.C.; Chipman, P.H.; Pinto, D.M.; Borowska, J.P.; Ichida, J.K.; Fawcett, J.P.; Zhang, Y.; Eggan, K.; Rafuse, V.F. Motoneurons derived from induced pluripotent stem cells develop mature phenotypes typical of endogenous spinal motoneurons. J. Neurosci. 2015, 35, 1291–1306. [Google Scholar] [CrossRef] [Green Version]
- Pepper, J.P.; Wang, T.V.; Hennes, V.; Sun, S.Y.; Ichida, J.K. Human Induced Pluripotent Stem Cell-Derived Motor Neuron Transplant for Neuromuscular Atrophy in a Mouse Model of Sciatic Nerve Injury. JAMA Facial Plast. Surg. 2017, 19, 197–205. [Google Scholar] [CrossRef]
- Charitos, I.A.; Ballini, A.; Cantore, S.; Boccellino, M.; Di Domenico, M.; Borsani, E.; Nocini, R.; Di Cosola, M.; Santacroce, L.; Bottalico, L. Stem Cells: A Historical Review about Biological, Religious, and Ethical Issues. Stem Cells Int. 2021, 2021, 9978837. [Google Scholar] [CrossRef]
- Guimaraes, M.Z.P.; De Vecchi, R.; Vitoria, G.; Sochacki, J.K.; Paulsen, B.S.; Lima, I.; Rodrigues da Silva, F.; da Costa, R.F.M.; Castro, N.G.; Breton, L.; et al. Generation of iPSC-Derived Human Peripheral Sensory Neurons Releasing Substance P Elicited by TRPV1 Agonists. Front. Mol. Neurosci. 2018, 11, 277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Güell, M.; Niu, D.; George, H.; Lesha, E.; Grishin, D.; Aach, J.; Shrock, E.; Xu, W.; Poci, J.; et al. Genome-Wide Inactivation of Porcine Endogenous Retroviruses (PERVs). Science 2015, 350, 1101–1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duggan, E.M.; Griesemer, A. Progress towards Xenogenic Tolerance. Curr. Opin. Organ Transplant. 2020, 25, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Sauvageot, C.M.; Stiles, C.D. Molecular Mechanisms Controlling Cortical Gliogenesis. Curr. Opin. Neurobiol. 2002, 12, 244–249. [Google Scholar] [CrossRef]
- Qian, X.; Shen, Q.; Goderie, S.K.; He, W.; Capela, A.; Davis, A.A.; Temple, S. Timing of CNS Cell Generation: A Programmed Sequence of Neuron and Glial Cell Production from Isolated Murine Cortical Stem Cells. Neuron 2000, 28, 69–80. [Google Scholar] [CrossRef] [Green Version]
- Cooper, D.K.; Ezzelarab, M.B.; Hara, H.; Iwase, H.; Lee, W.; Wijkstrom, M.; Bottino, R. The Pathobiology of Pig-to-Primate Xenotransplantation: A Historical Review. Xenotransplantation 2016, 23, 83–105. [Google Scholar] [CrossRef]
- Patience, C.; Takeuchi, Y.; Weiss, R.A. Infection of Human Cells by an Endogenous Retrovirus of Pigs. Nat. Med. 1997, 3, 282–286. [Google Scholar] [CrossRef]
- Matsumoto, S.; Abalovich, A.; Wechsler, C.; Wynyard, S.; Elliott, R.B. Clinical Benefit of Islet Xenotransplantation for the Treatment of Type 1 Diabetes. EBiomedicine 2016, 12, 255–262. [Google Scholar] [CrossRef] [Green Version]
- Deacon, T.; Schumacher, J.; Dinsmore, J.; Thomas, C.; Palmer, P.; Kott, S.; Edge, A.; Penney, D.; Kassissieh, S.; Dempsey, P.; et al. Histological Evidence of Fetal Pig Neural Cell Survival after Transplantation into a Patient with Parkinson’s Disease. Nat. Med. 1997, 3, 350–353. [Google Scholar] [CrossRef]
- Aron Badin, R.; Vadori, M.; Vanhove, B.; Nerriere-Daguin, V.; Naveilhan, P.; Neveu, I.; Jan, C.; Lévèque, X.; Venturi, E.; Mermillod, P.; et al. Cell Therapy for Parkinson’s Disease: A Translational Approach to Assess the Role of Local and Systemic Immunosuppression. Am. J. Transplant. 2016, 16, 2016–2029. [Google Scholar] [CrossRef] [Green Version]
- Björklund, A.; Stenevi, U.; Schmidt, R.H.; Dunnett, S.B.; Gage, F.H. Intracerebral Grafting of Neuronal Cell Suspensions. I. Introduction and General Methods of Preparation. Acta Physiol. Scand. Suppl. 1983, 522, 1–7. [Google Scholar]
- Barker, R.A.; Fricker, R.A.; Abrous, D.N.; Fawcett, J.; Dunnett, S.B. A Comparative Study of Preparation Techniques for Improving the Viability of Nigral Grafts Using Vital Stains, In Vitro Cultures, and In Vivo Grafts. Cell Transplant. 1995, 4, 173–200. [Google Scholar] [CrossRef]
- Barker, R.A.; Ratcliffe, E.; Richards, A.; Dunnett, S.B. Fetal Porcine Dopaminergic Cell Survival In Vitro and Its Relationship to Embryonic Age. Cell Transplant. 1999, 8, 593–599. [Google Scholar] [CrossRef]
- Grumbles, R.M.; Almeida, V.W.; Casella, G.T.; Wood, P.M.; Hemstapat, K.; Thomas, C.K. Motoneuron Replacement for Reinnervation of Skeletal Muscle in Adult Rats. J. Neuropathol. Exp. Neurol. 2012, 71, 921–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, S.; Kurimoto, S.; Nakano, T.; Yoneda, H.; Ishii, H.; Mita-Sugiura, S.; Hirata, H. Successful Transplantation of Motoneurons into the Peripheral Nerve Depends on the Number of Transplanted Cells. Nagoya J. Med. Sci. 2015, 77, 253–263. [Google Scholar]
- Sawada, H.; Kurimoto, S.; Tokutake, K.; Saeki, S.; Hirata, H. Optimal conditions for graft survival and reinnervation of denervated muscles after embryonic motoneuron transplantation into peripheral nerves undergoing Wallerian degeneration. J. Tissue Eng. Regen. Med. 2021, 15, 763–775. [Google Scholar] [CrossRef]
- Deshpande, D.M.; Kim, Y.S.; Martinez, T.; Carmen, J.; Dike, S.; Shats, I.; Rubin, L.L.; Drummond, J.; Krishnan, C.; Hoke, A.; et al. Recovery from Paralysis in Adult Rats Using Embryonic Stem Cells. Ann. Neurol. 2006, 60, 32–44. [Google Scholar] [CrossRef] [PubMed]
- Grumbles, R.M.; Casella, G.T.; Rudinsky, M.J.; Godfrey, S.; Wood, P.M.; Thomas, C.K. The Immunophilin Ligand FK506, but Not the P38 Kinase Inhibitor SB203580, Improves Function of Adult Rat Muscle Reinnervated from Transplants of Embryonic Neurons. Neuroscience 2005, 130, 619–630. [Google Scholar] [CrossRef]
- Grumbles, R.M.; Sesodia, S.; Wood, P.M.; Thomas, C.K. Neurotrophic Factors Improve Motoneuron Survival and Function of Muscle Reinnervated by Embryonic Neurons. J. Neuropathol. Exp. Neurol. 2009, 68, 736–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, P.; Matloub, H.S.; Sanger, J.R.; Narini, P. Gait Analysis in Rats with Peripheral Nerve Injury. Muscle Nerve 2001, 24, 231–239. [Google Scholar] [CrossRef]
- Rueden, C.T.; Schindelin, J.; Hiner, M.C.; DeZonia, B.E.; Walter, A.E.; Arena, E.T.; Eliceiri, K.W. ImageJ2: ImageJ for the next generation of scientific image data. BMC Bioinform. 2017, 18, 529. [Google Scholar] [CrossRef] [PubMed]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saeki, S.; Tokutake, K.; Takasu, M.; Kurimoto, S.; Asami, Y.; Onaka, K.; Saeki, M.; Hirata, H. Functional Reconstruction of Denervated Muscle by Xenotransplantation of Neural Cells from Porcine to Rat. Int. J. Mol. Sci. 2022, 23, 8773. https://doi.org/10.3390/ijms23158773
Saeki S, Tokutake K, Takasu M, Kurimoto S, Asami Y, Onaka K, Saeki M, Hirata H. Functional Reconstruction of Denervated Muscle by Xenotransplantation of Neural Cells from Porcine to Rat. International Journal of Molecular Sciences. 2022; 23(15):8773. https://doi.org/10.3390/ijms23158773
Chicago/Turabian StyleSaeki, Sota, Katsuhiro Tokutake, Masaki Takasu, Shigeru Kurimoto, Yuta Asami, Keiko Onaka, Masaomi Saeki, and Hitoshi Hirata. 2022. "Functional Reconstruction of Denervated Muscle by Xenotransplantation of Neural Cells from Porcine to Rat" International Journal of Molecular Sciences 23, no. 15: 8773. https://doi.org/10.3390/ijms23158773
APA StyleSaeki, S., Tokutake, K., Takasu, M., Kurimoto, S., Asami, Y., Onaka, K., Saeki, M., & Hirata, H. (2022). Functional Reconstruction of Denervated Muscle by Xenotransplantation of Neural Cells from Porcine to Rat. International Journal of Molecular Sciences, 23(15), 8773. https://doi.org/10.3390/ijms23158773