
Functionalized Nanogels with Endothelin-1 and Bradykinin Receptor Antagonist Peptides Decrease Inflammatory and Cartilage Degradation Markers of Osteoarthritis in a Horse Organoid Model of Cartilage

 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Results
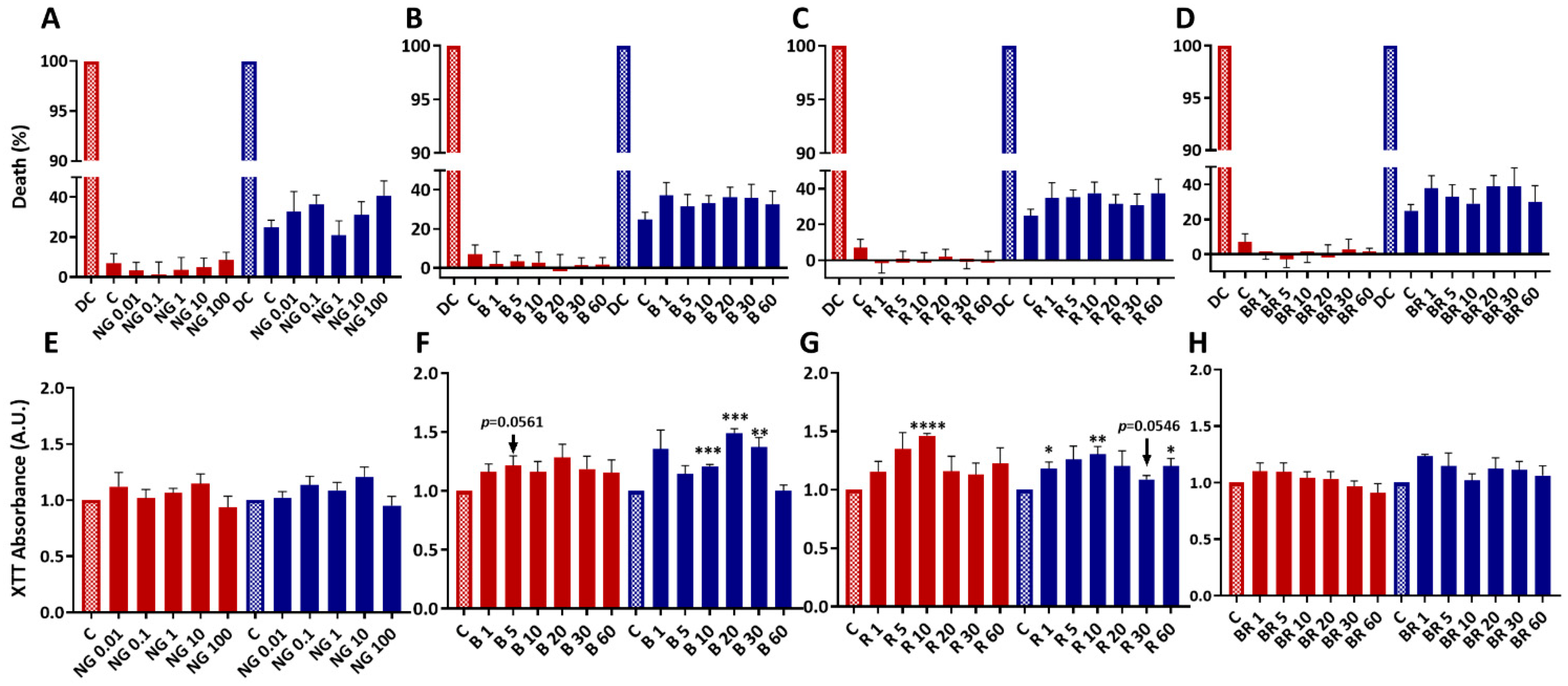
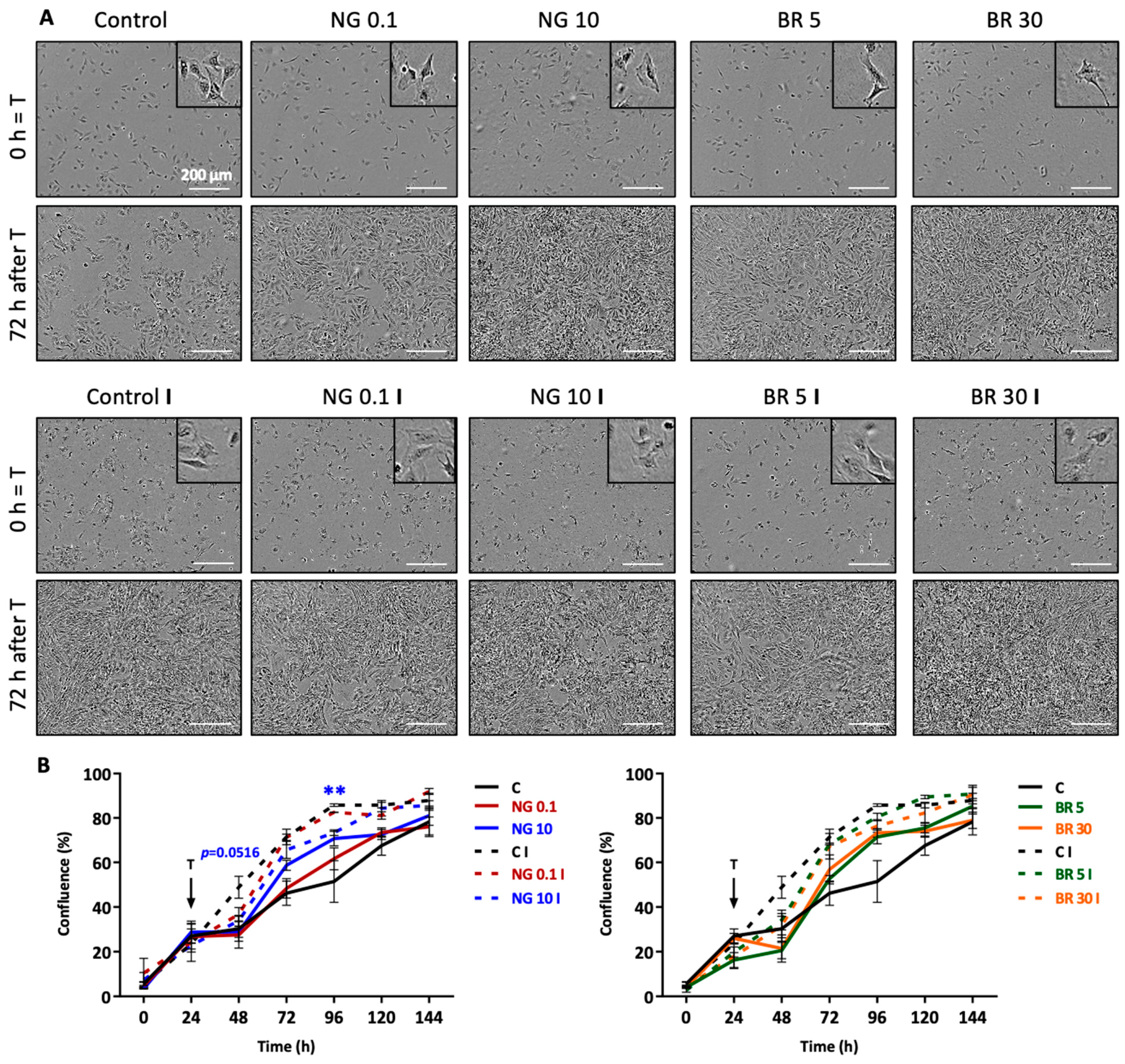
2.1. Nanogels Have No Cytotoxic Effects, Do Not Alter Viability, and Sustain Metabolic Activity and Proliferation of Equine Articular Chondrocytes
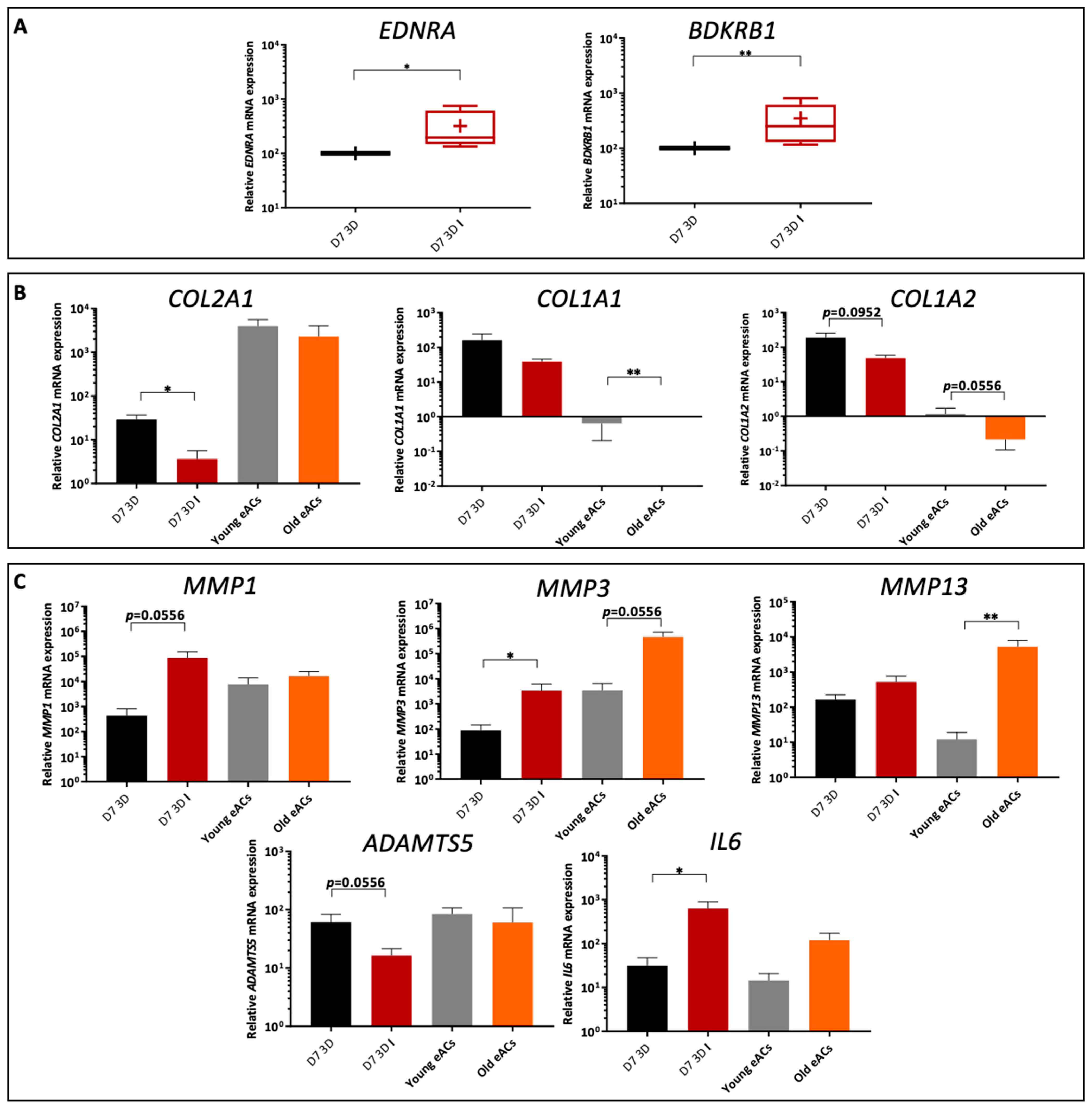
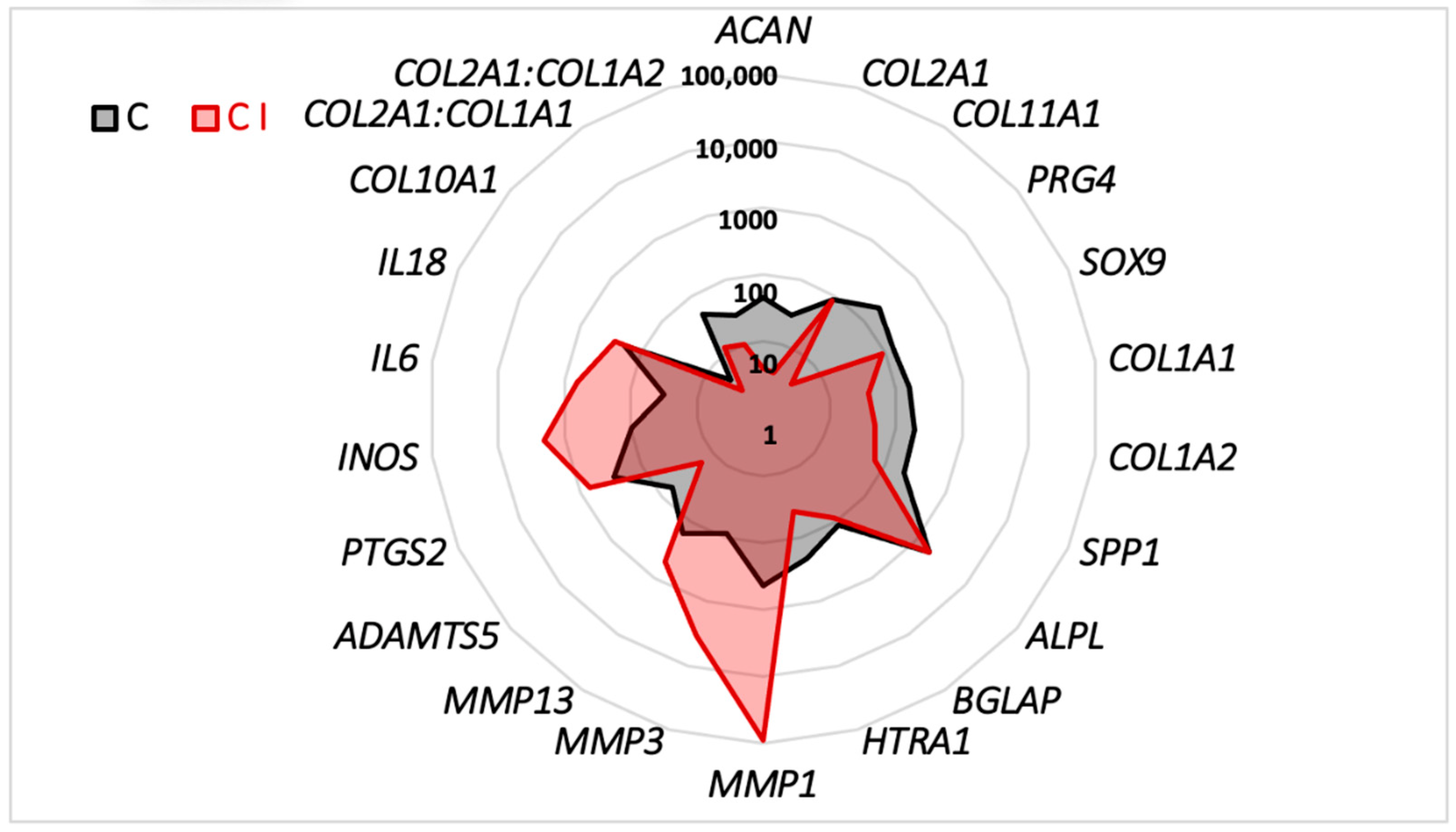
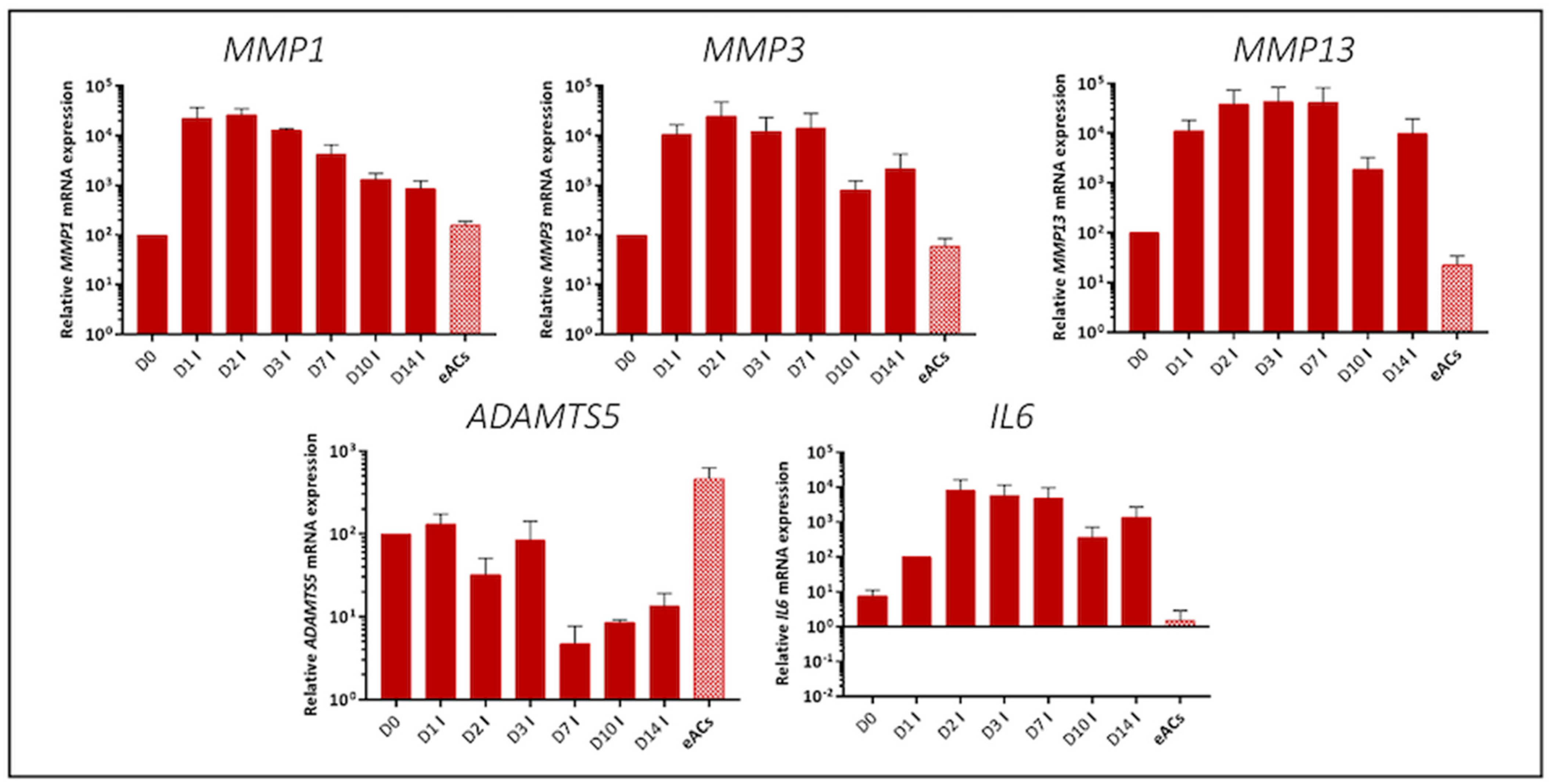
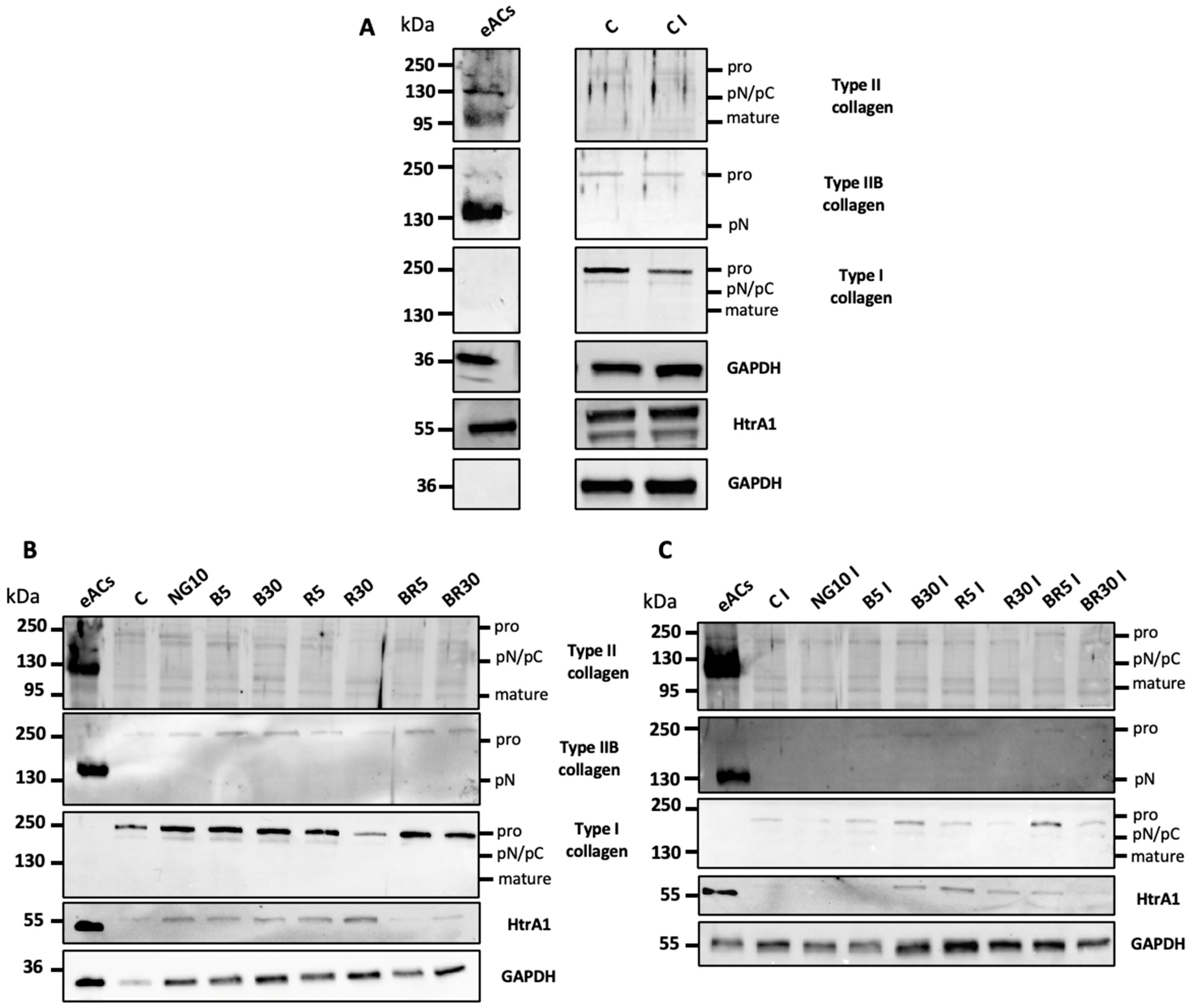
2.2. Characterization of the Organoid Model of Osteoarthritic Cartilage Tissue
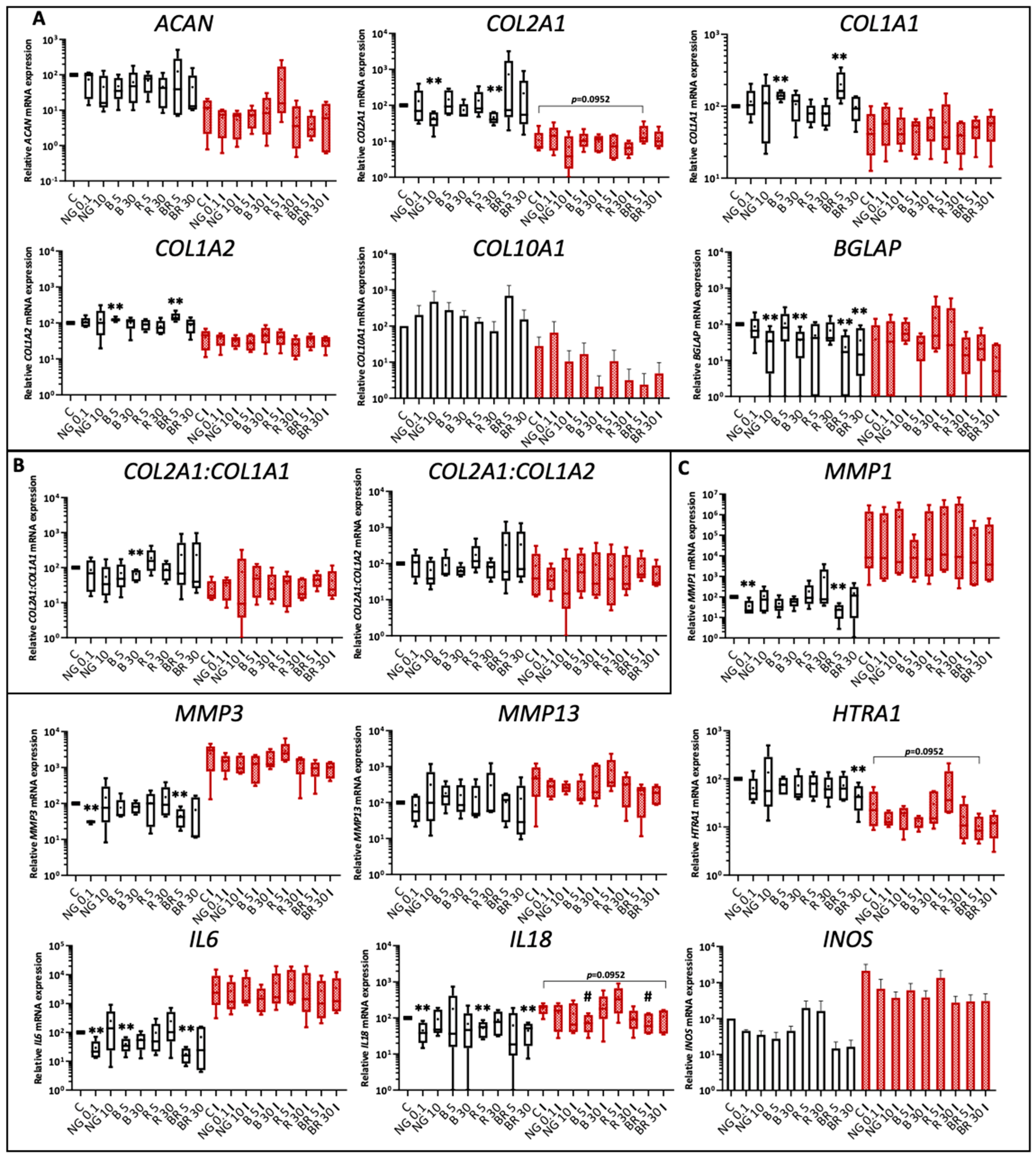
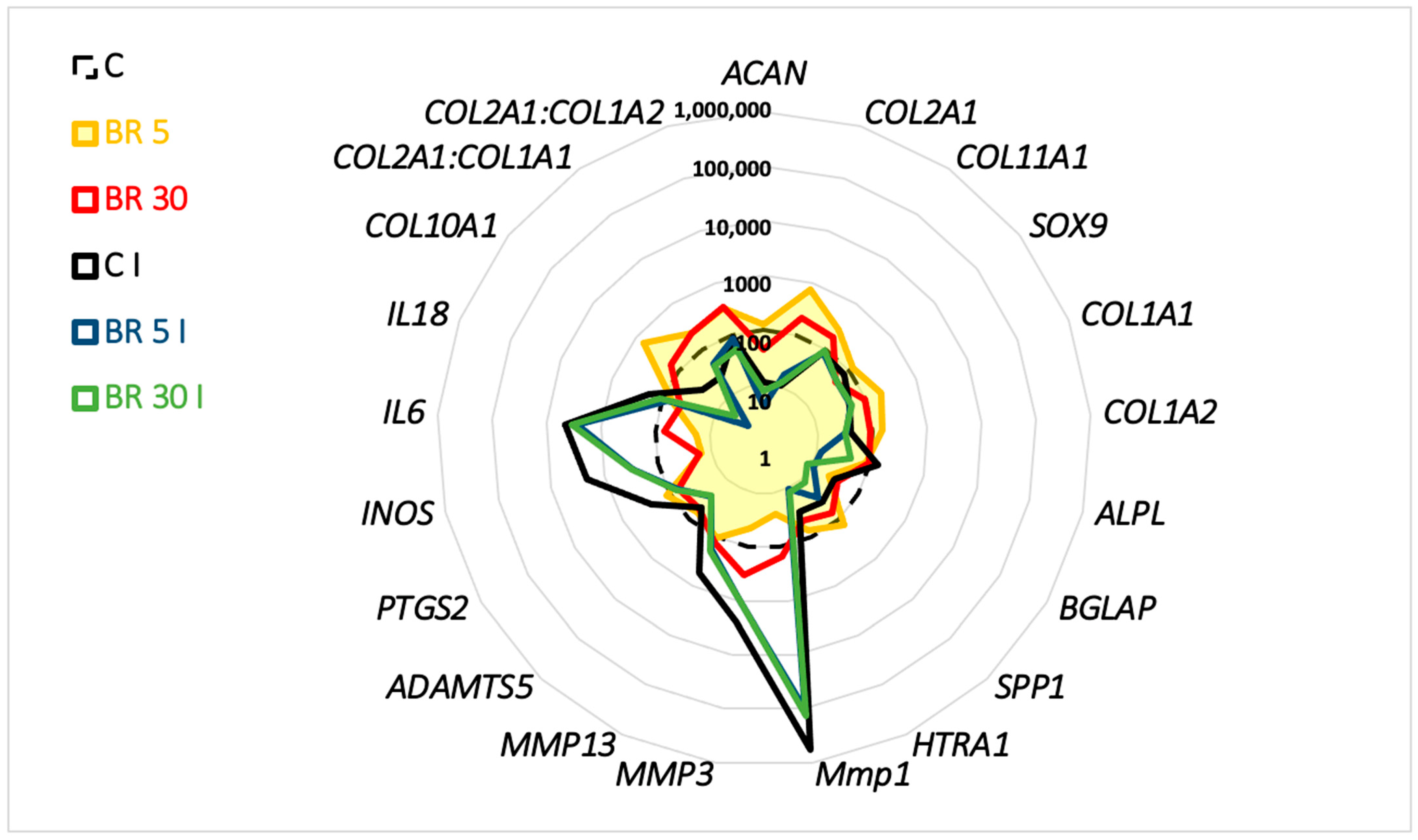
2.3. Functionalized Nanogels Decrease the Inflammatory and Catabolic Cartilage Markers
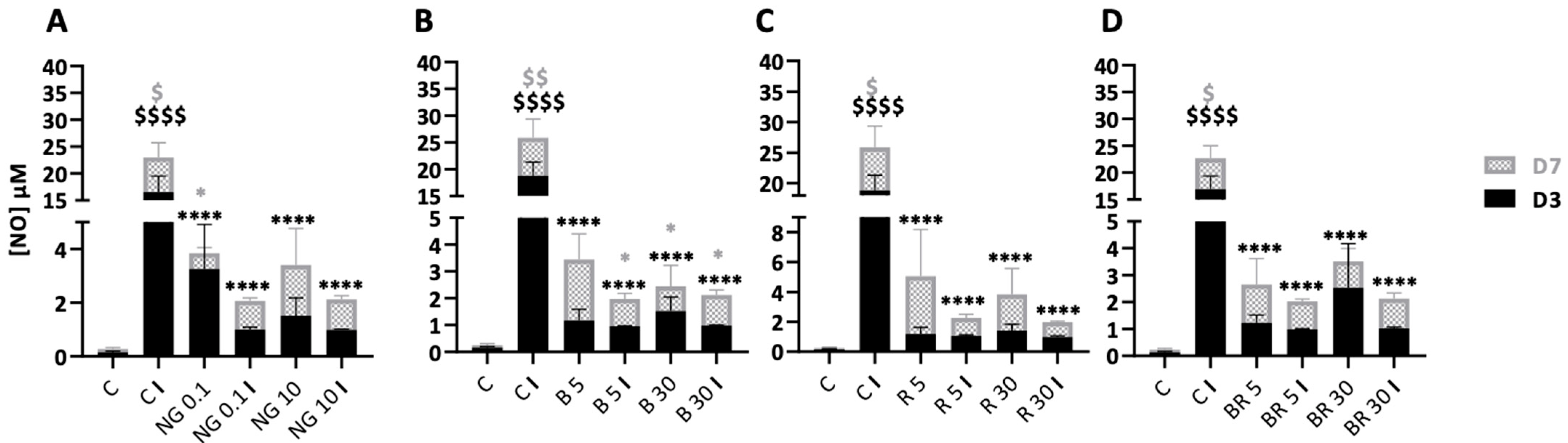
2.4. Non-Functionalized and Functionalized Nanogels Triggered a Decrease of Nitric Oxide Synthesis
3. Discussion
4. Materials and Methods
4.1. Nanogel Formulations
4.1.1. Peptides Grafting
4.1.2. Nanogel Synthesis, Purification, and Lyophilization
4.1.3. Determination of Drug Loading (DL%)
4.2. Isolation and Cell Culture
4.3. Culture of Equine Chondrocytes
4.4. Three-Dimensional Culture
4.5. Evaluation of Cytotoxicity
4.6. Determination of Metabolic Activity by the XTT Test
4.7. Evaluation of Proliferation
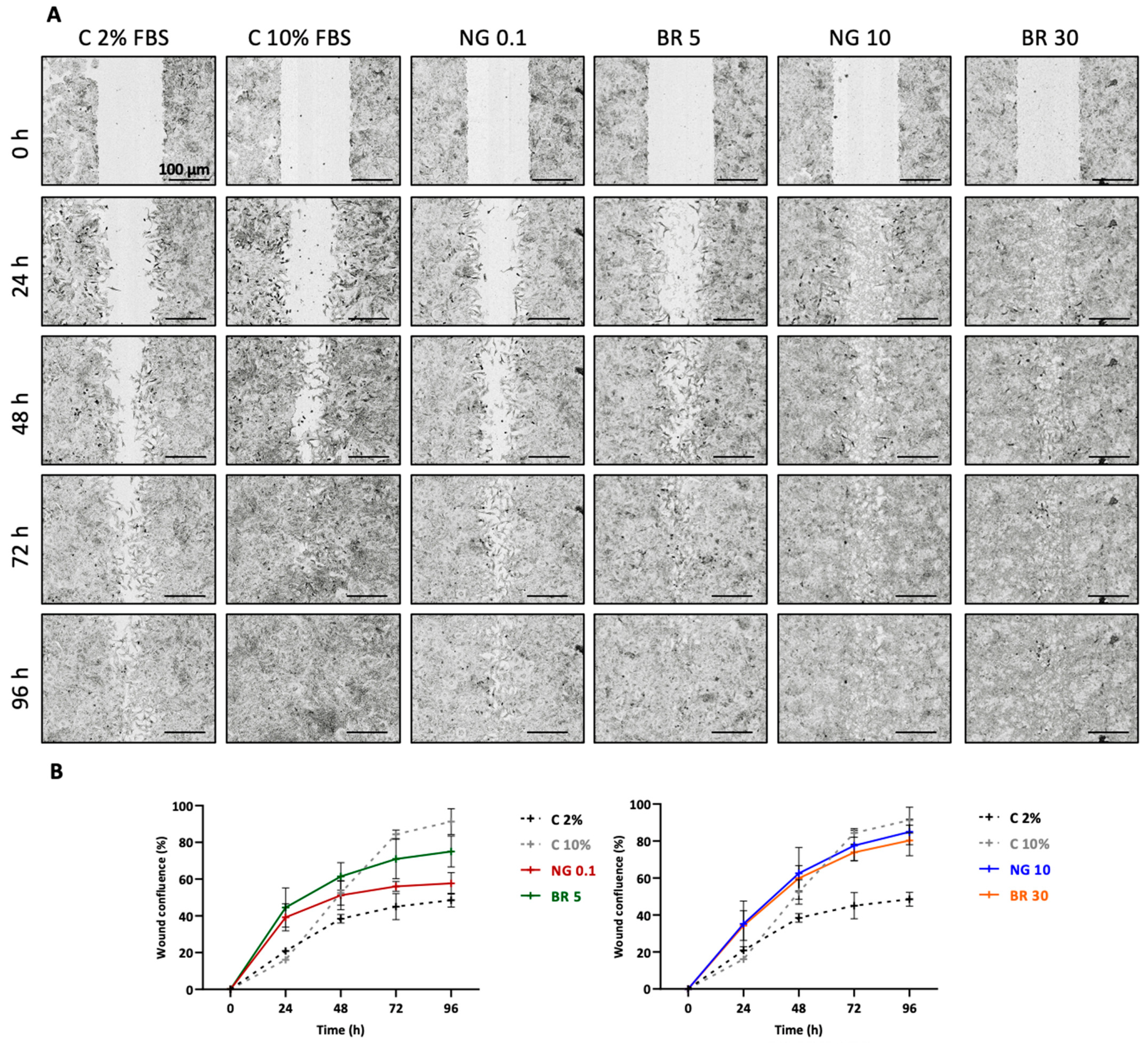
4.8. Scratch Wound Assay
4.9. RNA Isolation and RT-PCR
4.10. Western Blots
4.11. Nitrite Determination
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hunter, D.J.; March, L.; Chew, M. Osteoarthritis in 2020 and beyond: A Lancet Commission. Lancet 2020, 396, 1711–1712. [Google Scholar] [CrossRef]
- Sophia Fox, A.J.; Bedi, A.; Rodeo, S.A. The Basic Science of Articular Cartilage: Structure, Composition, and Function. Sports Health 2009, 1, 461–468. [Google Scholar] [CrossRef]
- Demoor, M.; Ollitrault, D.; Gomez-Leduc, T.; Bouyoucef, M.; Hervieu, M.; Fabre, H.; Lafont, J.; Denoix, J.-M.; Audigié, F.; Mallein-Gerin, F.; et al. Cartilage Tissue Engineering: Molecular Control of Chondrocyte Differentiation for Proper Cartilage Matrix Reconstruction. Biochim. Biophys. Acta 2014, 1840, 2414–2440. [Google Scholar] [CrossRef]
- Berenbaum, F. Osteoarthritis as an inflammatory disease (osteoarthritis is not osteoarthrosis!). Osteoarthr. Cartil. 2013, 21, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Goldring, M.B.; Goldring, S.R. Articular cartilage and subchondral bone in the pathogenesis of osteoarthritis. Ann. N. Y. Acad. Sci. 2010, 1192, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Xia, B.; Chen, D.; Zhang, J.; Hu, S.; Jin, H.; Tong, P. Osteoarthritis Pathogenesis: A Review of Molecular Mechanisms. Calcif. Tissue Res. 2014, 95, 495–505. [Google Scholar] [CrossRef] [PubMed]
- Goldring, M.B.; Otero, M. Inflammation in osteoarthritis. Curr. Opin. Rheumatol. 2011, 23, 471–478. [Google Scholar] [CrossRef] [PubMed]
- McIlwraith, C.W.; Frisbie, D.D.; Kawcak, C.E. The horse as a model of naturally occurring osteoarthritis. Bone Jt. Res. 2012, 1, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Perkins, N.R.; Reid, S.W.J.; Morris, R.S. Effect of training location and time period on racehorse performance in New Zealand. 2. Multivariable analysis. N. Z. Vet. J. 2004, 52, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Goldring, M.B.; Goldring, S.R. Osteoarthritis. J. Cell. Physiol. 2007, 213, 626–634. [Google Scholar] [CrossRef] [PubMed]
- Malda, J.; Benders, K.E.; Klein, T.J.; de Grauw, J.C.; Kik, M.J.; Hutmacher, D.W.; Saris, D.B.; van Weeren, P.R.; Dhert, W.J. Comparative study of depth-dependent characteristics of equine and human osteochondral tissue from the medial and lateral femoral condyles. Osteoarthr. Cartil. 2012, 20, 1147–1151. [Google Scholar] [CrossRef] [PubMed]
- Latourte, A.; Kloppenburg, M.; Richette, P. Emerging pharmaceutical therapies for osteoarthritis. Nat. Rev. Rheumatol. 2020, 16, 673–688. [Google Scholar] [CrossRef] [PubMed]
- Kolasinski, S.L.; Neogi, T.; Hochberg, M.C.; Oatis, C.; Guyatt, G.; Block, J.; Callahan, L.; Copenhaver, C.; Dodge, C.; Felson, D.; et al. 2019 American College of Rheumatology/Arthritis Foundation Guideline for the Management of Osteoarthritis of the Hand, Hip, and Knee. Arthritis Care Res. 2020, 72, 149–162. [Google Scholar] [CrossRef] [PubMed]
- Alexander, L.A.M.; Denisov, L.N.; Zotkin, E.G.; Dydykina, I.S.; Kochish, A.Y.; Rodionova, S.S.; Trofimov, E.A.; Yakupova, S.P.; Yakupov, E.Z.; Gallelli, L. Pharmacological Management of Osteoarthritis with a Focus on Symptomatic Slow-Acting Drugs. J. Clin. Rheumatol. 2021, 27, e533–e539. [Google Scholar] [CrossRef]
- Pavone, V.; Vescio, A.; Turchetta, M.; Giardina, S.M.C.; Culmone, A.; Testa, G. Injection-Based Management of Osteoarthritis of the Knee: A Systematic Review of Guidelines. Front. Pharmacol. 2021, 12, 661805. [Google Scholar] [CrossRef]
- Kloppenburg, M.; Berenbaum, F. Osteoarthritis year in review 2019: Epidemiology and therapy. Osteoarthr. Cartil. 2020, 28, 242–248. [Google Scholar] [CrossRef]
- Santilli, V.; Paoloni, M.; Mangone, M.; Alviti, F.; Bernetti, A. Hyaluronic acid in the management of osteoarthritis: Injection therapies innovations. Clin. Cases Miner. Bone Metab. 2016, 13, 131–134. [Google Scholar] [CrossRef]
- Koh, R.H.; Jin, Y.; Kim, J.; Hwang, N.S. Inflammation-Modulating Hydrogels for Osteoarthritis Cartilage Tissue Engineering. Cells 2020, 9, 419. [Google Scholar] [CrossRef]
- Wang, A.; Gu, F.; Zhang, L.; Chan, J.; Radovic-Moreno, A.; Shaikh, M.R.; Farokhzad, O.C. Biofunctionalized targeted nanoparticles for therapeutic applications. Expert Opin. Biol. Ther. 2008, 8, 1063–1070. [Google Scholar] [CrossRef]
- Jang, S.; Lee, K.; Ju, J.H. Recent Updates of Diagnosis, Pathophysiology, and Treatment on Osteoarthritis of the Knee. Int. J. Mol. Sci. 2021, 22, 2619. [Google Scholar] [CrossRef]
- Rahimi, M.; Charmi, G.; Matyjaszewski, K.; Banquy, X.; Pietrasik, J. Recent developments in natural and synthetic polymeric drug delivery systems used for the treatment of osteoarthritis. Acta Biomater. 2021, 123, 31–50. [Google Scholar] [CrossRef] [PubMed]
- Kaderli, S.; Boulocher, C.; Pillet-Michelland, E.; Watrelot-Virieux, D.; Rougemont, A.-L.; Roger, T.; Viguier, E.; Gurny, R.; Scapozza, L.; Jordan, O. A novel biocompatible hyaluronic acid–chitosan hybrid hydrogel for osteoarthrosis therapy. Int. J. Pharm. 2015, 483, 158–168. [Google Scholar] [CrossRef] [PubMed]
- De Falco, L.; Fioravanti, A.; Galeazzi, M.; Tenti, S. Bradykinin and its role in osteoarthritis. Reumatismo 2013, 65, 97–104. [Google Scholar] [CrossRef]
- Kaufman, G.N.; Zaouter, C.; Valteau, B.; Sirois, P.; Moldovan, F. Nociceptive tolerance is improved by bradykinin receptor B1 antagonism and joint morphology is protected by both endothelin type A and bradykinin receptor B1 antagonism in a surgical model of osteoarthritis. Arthritis Res. Ther. 2011, 13, R76. [Google Scholar] [CrossRef] [PubMed]
- Yanagisawa, H.; Hammer, R.E.; Richardson, J.A.; Williams, S.C.; Clouthier, D.E.; Yanagisawa, M. Role of Endothelin-1/Endothelin-A receptor-mediated signaling pathway in the aortic arch patterning in mice. J. Clin. Investig. 1998, 102, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Yanagisawa, M.; Kurihara, H.; Kimura, S.; Tomobe, Y.; Kobayashi, M.; Mitsui, Y.; Yazaki, Y.; Goto, K.; Masaki, T. A novel potent vasoconstrictor peptide produced by vascular endothelial cells. Nature 1988, 332, 411–415. [Google Scholar] [CrossRef] [PubMed]
- Manacu, C.A.; Martel-Pelletier, J.; Roy-Beaudry, M.; Pelletier, J.P.; Fernandes, J.C.; Shipkolye, F.; Moldovan, F. Endothelin-1 in osteoarthritic chondrocytes triggers nitric oxide production and upregulates collagenase production. Arthritis Res. Ther. 2005, 7, R324–R332. [Google Scholar] [CrossRef]
- Khatib, A.M.; Lomri, A.; Moldovan, F.; Soliman, H.; Fiet, J.; Mitrovic, D.R. Endothelin 1 Receptors, Signal Transduction and Effects on DNA and Proteoglycans Synthesis in Rat Articular Chondrocytes. Cytokine 1998, 10, 669–679. [Google Scholar] [CrossRef]
- Roy-Beaudry, M.; Martel-Pelletier, J.; Pelletier, J.-P.; M’Barek, K.N.; Christgau, S.; Shipkolye, F.; Moldovan, F. Endothelin 1 promotes osteoarthritic cartilage degradation via matrix metalloprotease 1 and matrix metalloprotease 13 induction. Arthritis Care Res. 2003, 48, 2855–2864. [Google Scholar] [CrossRef]
- Cruwys, S.; Garrett, N.; Perkins, M.; Blake, D.; Kidd, B. The role of bradykinin B1 receptors in the maintenance of intra-articular plasma extravasation in chronic antigen-induced arthritis. J. Cereb. Blood Flow Metab. 1994, 113, 940–944. [Google Scholar] [CrossRef]
- Meini, S.; Cucchi, P.; Catalani, C.; Bellucci, F.; Giuliani, S.; Maggi, C. Bradykinin and B2 receptor antagonism in rat and human articular chondrocytes. J. Cereb. Blood Flow Metab. 2011, 162, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Galéra, P.; Ollitrault, D.; Legendre, F.; Demoor, M.; Mallein-Gerin, F.; Boumediene, K.; Herbage, B.; Duterque-Coquillaud, M.; Damour, O. Brevet Français FR 2965278 du 23 Septembre 2010. Method for Obtaining Differentiated Articular Chondrocytes in vitro or ex vivo and Uses of Same. WO Patent 2012/038668, 29 March 2012. [Google Scholar]
- Ollitrault, D.; Legendre, F.; Drougard, C.; Briand, M.; Benateau, H.; Goux, D.; Chajra, H.; Poulain, L.; Hartmann, D.; Vivien, D.; et al. BMP-2, Hypoxia, and COL1A1/HtrA1 SiRNAs Favor Neo-Cartilage Hyaline Matrix Formation in Chondrocytes. Tissue Eng. Part C Methods 2015, 21, 133–147. [Google Scholar] [CrossRef] [PubMed]
- Bourdon, B.; Contentin, R.; Cassé, F.; Maspimby, C.; Oddoux, S.; Noël, A.; Legendre, F.; Gruchy, N.; Galéra, P. Marine Collagen Hydrolysates Downregulate the Synthesis of Pro-Catabolic and Pro-Inflammatory Markers of Osteoarthritis and Favor Collagen Production and Metabolic Activity in Equine Articular Chondrocyte Organoids. Int. J. Mol. Sci. 2021, 22, 580. [Google Scholar] [CrossRef]
- Rakic, R.; Bourdon, B.; Hervieu, M.; Branly, T.; Legendre, F.; Saulnier, N.; Audigié, F.; Maddens, S.; Demoor, M.; Galera, P. RNA Interference and BMP-2 Stimulation Allows Equine Chondrocytes Redifferentiation in 3D-Hypoxia Cell Culture Model: Application for Matrix-Induced Autologous Chondrocyte Implantation. Int. J. Mol. Sci. 2017, 18, 1842. [Google Scholar] [CrossRef]
- Van der Kraan, P.M.; Buma, P.; Van Kuppevelt, T.; Van den Berg, W.B. Interaction of chondrocytes, extracellular matrix and growth factors: Relevance for articular cartilage tissue engineering. Osteoarthr. Cartil. 2002, 10, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Darling, E.M.; Athanasiou, K.A. Rapid phenotypic changes in passaged articular chondrocyte subpopulations. J. Orthop. Res. 2005, 23, 425–432. [Google Scholar] [CrossRef]
- Schnabel, M.; Marlovits, S.; Eckhoff, G.; Fichtel, I.; Gotzen, L.; Vécsei, V.; Schlegel, J. Dedifferentiation-associated changes in morphology and gene expression in primary human articular chondrocytes in cell culture. Osteoarthr. Cartil. 2002, 10, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Charlier, E.; Deroyer, C.; Ciregia, F.; Malaise, O.; Neuville, S.; Plener, Z.; Malaise, M.; de Seny, D. Chondrocyte dedifferentiation and osteoarthritis (OA). Biochem. Pharmacol. 2019, 165, 49–65. [Google Scholar] [CrossRef]
- Garcia, A.; Moldovan, F.; Roullin, V.G.; Banquy, X.; Sauve, S. Stability and release of two peptides in a novel peptide-based drugs for local intra-articular purpose in osteoarthritis. Front. Bioeng. Biotechnol. 2016, 4. [Google Scholar] [CrossRef]
- Lange, C.; Madry, H.; Venkatesan, J.K.; Schmitt, G.; Speicher-Mentges, S.; Zurakowski, D.; Menger, M.D.; Laschke, M.W.; Cucchiarini, M. RAAV-Mediated Sox9 Overexpression Improves the Repair of Osteochondral Defects in a Clinically Relevant Large Animal Model Over Time In Vivo and Reduces Perifocal Osteoarthritic Changes. Am. J. Sports Med. 2021, 49, 3696–3707. [Google Scholar] [CrossRef]
- Seol, D.; Choe, H.H.; Zheng, H.; Brouillette, M.J.; Fredericks, D.C.; Petersen, M.E.B.; Song, I.; Chakka, L.R.; Salem, A.K.; Martin, J.A. Intra-Articular Adeno-Associated Virus-Mediated Proteoglycan 4 Gene Therapy for Preventing Posttraumatic Osteoarthritis. Hum. Gene Ther. 2022, 33, 529–540. [Google Scholar] [CrossRef] [PubMed]
- Carmona, J.U.; Ríos, D.L.; López, C.; Álvarez, M.E.; Pérez, J.E.; Bohórquez, M.E. In vitro effects of platelet-rich gel supernatants on histology and chondrocyte apoptosis scores, hyaluronan release and gene expression of equine cartilage explants challenged with lipopolysaccharide. BMC Vet. Res. 2016, 12, 135. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Ma, Y.; Yao, X.; Zhou, W.; Wang, X.; Li, C.; Lin, J.; He, Q.; Leptihn, S.; Ouyang, H. Advanced hydrogels for the repair of cartilage defects and regeneration. Bioact. Mater. 2021, 6, 998–1011. [Google Scholar] [CrossRef] [PubMed]
- Faivre, J.; Shrestha, B.R.; Burdynska, J.; Xie, G.; Moldovan, F.; Delair, T.; Benayoun, S.; David, L.; Matyjaszewski, K.; Banquy, X. Wear Protection without Surface Modification Using a Synergistic Mixture of Molecular Brushes and Linear Polymers. ACS Nano 2017, 11, 1762–1769. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.-Y.; Lin, Y.-C.; Lu, H.-T.; Ho, Y.-C.; Weng, S.-C.; Tsai, M.-L.; Mi, F.-L. A novel injectable in situ forming gel based on carboxymethyl hexanoyl chitosan/hyaluronic acid polymer blending for sustained release of berberine. Carbohydr. Polym. 2019, 206, 664–673. [Google Scholar] [CrossRef]
- Manivong, S.; Garcia Ac, A.; Patten, S.A.; Fernandes, J.C.; Benderdour, M.; Banquy, X.; Moldovan, F.; Roullin, V.G. Chitosan-Based Nanogels: Synthesis and Toxicity Profile for Drug Delivery to Articular Joints. Nanomaterials 2022, 12, 1337. [Google Scholar] [CrossRef]
- Bolduc, J.A.; Collins, J.A.; Loeser, R.F. Reactive oxygen species, aging and articular cartilage homeostasis. Free Radic. Biol. Med. 2019, 132, 73–82. [Google Scholar] [CrossRef]
- McCoy, A.M. Animal Models of Osteoarthritis. Vet. Pathol. 2015, 52, 803–818. [Google Scholar] [CrossRef]
- Towle, C.A.; Hung, H.H.; Bonassar, L.J.; Treadwell, B.V.; Mangham, D.C. Detection of interleukin-1 in the cartilage of patients with osteoarthritis: A possible autocrine/paracrine role in pathogenesis. Osteoarthr. Cartil. 1997, 5, 293–300. [Google Scholar] [CrossRef]
- Bondeson, J.; Wainwright, S.; Hughes, C.; Caterson, B. The regulation of the ADAMTS4 and ADAMTS5 aggrecanases in osteoarthritis: A review. Clin. Exp. Rheumatol. 2008, 26, 139–145. [Google Scholar]
- Naito, S.; Shiomi, T.; Okada, A.; Kimura, T.; Chijiiwa, M.; Fujita, Y.; Yatabe, T.; Komiya, K.; Enomoto, H.; Fujikawa, K.; et al. Expression of ADAMTS4 (aggrecanase-1) in human osteoarthritic cartilage. Pathol. Int. 2007, 57, 703–711. [Google Scholar] [CrossRef] [PubMed]
- Grau, S.; Richards, P.J.; Kerr, B.; Hughes, C.; Caterson, B.; Williams, A.S.; Junker, U.; Jones, S.A.; Clausen, T.; Ehrmann, M. The Role of Human HtrA1 in Arthritic Disease. J. Biol. Chem. 2006, 281, 6124–6129. [Google Scholar] [CrossRef] [PubMed]
- Roy, H.S.; Singh, R.; Ghosh, D. Recent advances in nanotherapeutic strategies that target nitric oxide pathway for preventing cartilage degeneration. Nitric Oxide 2021, 109–110, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Khatib, A.-M.; Siegfried, G.; Messai, H.; Moldovan, F.; Mitrovic, D.R. Mechanism of Inhibition of Endothelin-1-Stimulated Proteoglycan and Collagen Synthesis in Rat Articular Chondrocytes. Cytokine 2002, 17, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Rim, Y.A.; Nam, Y.; Ju, J.H. The Role of Chondrocyte Hypertrophy and Senescence in Osteoarthritis Initiation and Progression. Int. J. Mol. Sci. 2020, 21, 2358. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Size (DH, nm) | PdI | ZP (mV) | DL (%) | |
|---|---|---|---|---|
| BQ-123-CHI | 733 ± 169 | 0.60 ± 0.3 | 47 ± 5 | 1.35–2.60 |
| R-954-HA | 429 ± 52 | 0.29 ± 0.0 | 51 ± 8 | 0.30–0.63 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cullier, A.; Cassé, F.; Manivong, S.; Contentin, R.; Legendre, F.; Garcia Ac, A.; Sirois, P.; Roullin, G.; Banquy, X.; Moldovan, F.; et al. Functionalized Nanogels with Endothelin-1 and Bradykinin Receptor Antagonist Peptides Decrease Inflammatory and Cartilage Degradation Markers of Osteoarthritis in a Horse Organoid Model of Cartilage. Int. J. Mol. Sci. 2022, 23, 8949. https://doi.org/10.3390/ijms23168949
Cullier A, Cassé F, Manivong S, Contentin R, Legendre F, Garcia Ac A, Sirois P, Roullin G, Banquy X, Moldovan F, et al. Functionalized Nanogels with Endothelin-1 and Bradykinin Receptor Antagonist Peptides Decrease Inflammatory and Cartilage Degradation Markers of Osteoarthritis in a Horse Organoid Model of Cartilage. International Journal of Molecular Sciences. 2022; 23(16):8949. https://doi.org/10.3390/ijms23168949
Chicago/Turabian StyleCullier, Aurélie, Frédéric Cassé, Seng Manivong, Romain Contentin, Florence Legendre, Aracéli Garcia Ac, Pierre Sirois, Gaëlle Roullin, Xavier Banquy, Florina Moldovan, and et al. 2022. "Functionalized Nanogels with Endothelin-1 and Bradykinin Receptor Antagonist Peptides Decrease Inflammatory and Cartilage Degradation Markers of Osteoarthritis in a Horse Organoid Model of Cartilage" International Journal of Molecular Sciences 23, no. 16: 8949. https://doi.org/10.3390/ijms23168949
APA StyleCullier, A., Cassé, F., Manivong, S., Contentin, R., Legendre, F., Garcia Ac, A., Sirois, P., Roullin, G., Banquy, X., Moldovan, F., Bertoni, L., Audigié, F., Galéra, P., & Demoor, M. (2022). Functionalized Nanogels with Endothelin-1 and Bradykinin Receptor Antagonist Peptides Decrease Inflammatory and Cartilage Degradation Markers of Osteoarthritis in a Horse Organoid Model of Cartilage. International Journal of Molecular Sciences, 23(16), 8949. https://doi.org/10.3390/ijms23168949