
NOS1AP Interacts with α-Synuclein and Aggregates in Yeast and Mammalian Cells

 , , and
, , and 
Abstract
:1. Introduction
2. Results
2.1. NOS1AP Is a Potential Aggregation-Prone Protein Interacting with α-Synuclein
2.2. NOS1AP Does Not Demonstrate Amyloid Properties in C-DAG
2.3. NOS1AP Is Able to Aggregate When Overproduced in Eukaryotic Cells
2.4. NOS1AP Forms Non-Amyloid Aggregates In Vitro
2.5. NOS1AP Interacts with α-Synuclein in Mammalian and Yeast Cells
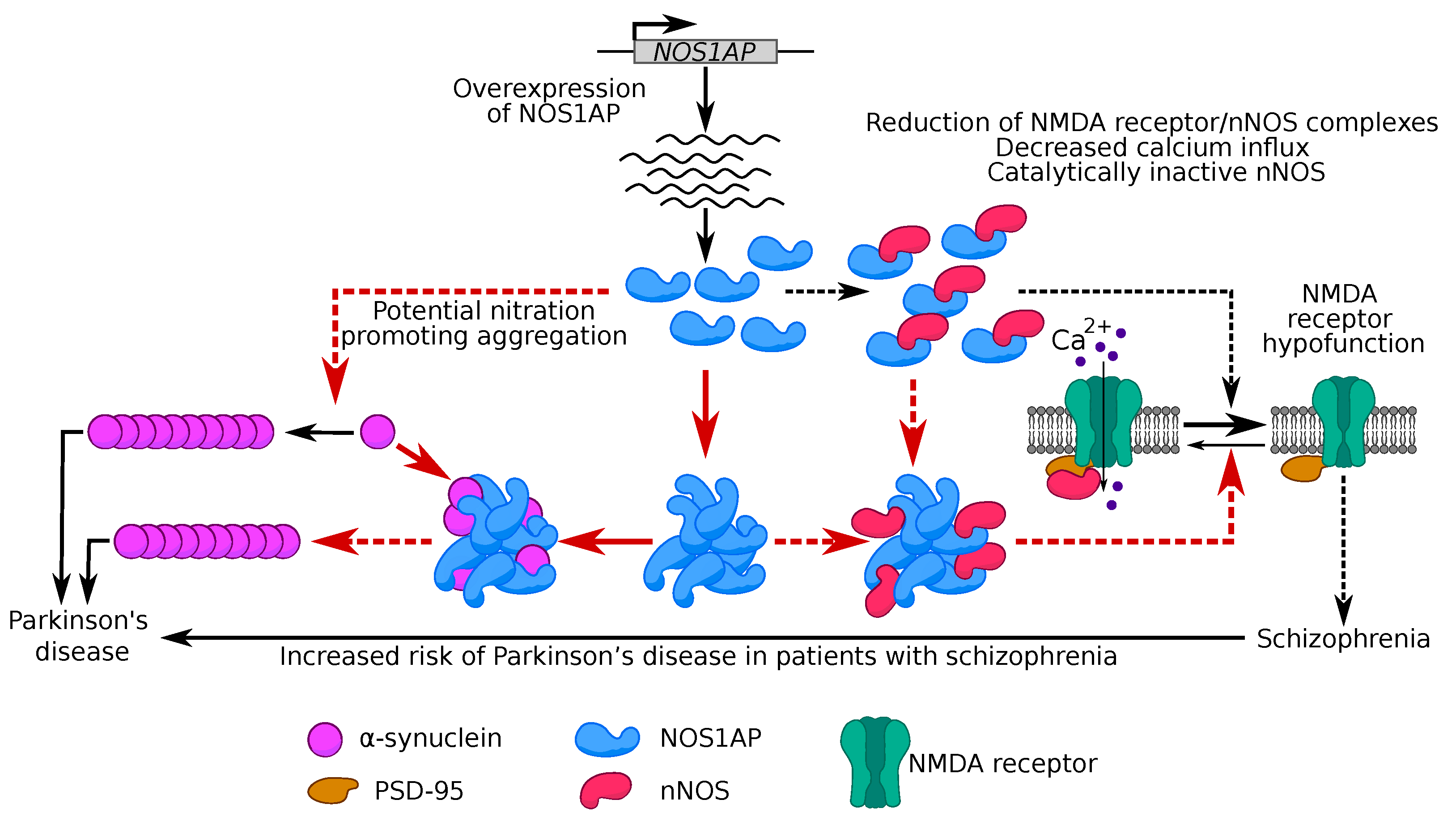
3. Discussion
3.1. NOS1AP Can Form Non-Amyloid Aggregates When Overproduced
3.2. Potential Role of NOS1AP Aggregation in Schizophrenia Development
3.3. Potential Role of NOS1AP and α-Synuclein Co-Aggregation in Development of Parkinson’s Disease
4. Methods and Materials
4.1. Bioinformatic Analysis
4.2. Plasmid Construction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vector | Signal Peptide, Fluorescent Protein, Tag, and Promoter | Experiment | Reference |
|---|---|---|---|
| pVSGW-ccdB | CsgA signal peptide on the N-termini of a protein, BAD promoter | C-DAG system | [40] |
| pgLAP1 | EGFP on the N-termini of a protein, CMV promoter | Analysis of protein localization in mammalian cells | A gift from Peter Jackson (Addgene plasmid #19702 http://n2t.net/addgene:19702 accessed on 13 July 2022) |
| pDEST-V1-ORF | N-terminal fusion protein with Venus fluorescent protein fragment 1 (V1), CMV promoter | Analysis of protein-protein interaction in mammalian cells | A gift from Darren Saunders (Addgene plasmid #73635 http://n2t.net/addgene:73635 accessed on 13 July 2022) [52] |
| pDEST-V2-ORF | N-terminal fusion protein with Venus fluorescent protein fragment 2 (V2), CMV promoter | Analysis of protein-protein interaction in mammalian cells | A gift from Darren Saunders (Addgene plasmid #73636 http://n2t.net/addgene:73636 accessed on 13 July 2022) [52] |
| pAG416GPD-EGFP-ccdB | EGFP on the N-termini of a protein, GAP promoter | Analysis of protein localization in yeast cells | A gift from Susan Lindquist (Addgene plasmid #14316 http://n2t.net/addgene:14316 accessed on 13 July 2022) |
| pAG416GPD-EYFP-ccdB | EYFP on the N-termini of a protein, GAP promoter | Analysis of protein localization in yeast cells | A gift from Susan Lindquist (Addgene plasmid #14340 http://n2t.net/addgene:14340 accessed on 13 July 2022) |
| pAG415GPD-Cerulean-ccdB | Cerulean on the N-termini of a protein, GAP promoter | Analysis of protein localization in yeast cells | A gift from Susan Lindquist (Addgene plasmid #14410 http://n2t.net/addgene:14410 accessed on 13 July 2022) |
| pDest-527 | His6 tag on the N-termini of a protein, T7 promoter | Protein purification | A gift from Dominic Esposito (Addgene plasmid #11518 http://n2t.net/addgene:11518 accessed on 13 July 2022) |
4.3. Yeast Strains, Transformation, and Microscopy
4.4. Curli-Dependent Amyloid Generator (C-DAG)
4.5. Cell Transfection and Microscopy
4.6. NOS1AP Antibodies
4.7. NOS1AP Aggregation, Electron Microscopy, and Congo Red Staining
4.8. Protein Electrophoresis and Hybridization
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Aβ | amyloid-β |
| AD | Alzheimer’s disease |
| BME | β-mercaptoethanol |
| C-DAG | curli-dependent amyloid generator |
| NMDA | N-methyl-D-aspartate |
| NO | nitric oxide |
| PD | Parkinson’s disease |
| SCZ | Schizophrenia |
| SDD-AGE | semi-denaturing detergent-agarose gel electrophoresis |
| αSyn | α-synuclein |
References
- Matiiv, A.B.; Trubitsina, N.P.; Matveenko, A.G.; Barbitoff, Y.A.; Zhouravleva, G.A.; Bondarev, S.A. Amyloid and amyloid-like aggregates: Diversity and the term crisis. Biochemistry 2020, 85, 1011–1034. [Google Scholar] [CrossRef]
- Sergeeva, A.V.; Galkin, A.P. Functional amyloids of eukaryotes: Criteria, classification, and biological significance. Curr. Genet. 2020, 66, 849–866. [Google Scholar] [CrossRef]
- Nizhnikov, A.A.; Antonets, K.S.; Inge-Vechtomov, S.G. Amyloids: From pathogenesis to function. Biochemistry 2015, 80, 1127–1144. [Google Scholar] [CrossRef] [PubMed]
- Benson, M.D.; Buxbaum, J.N.; Eisenberg, D.S.; Merlini, G.; Saraiva, M.J.; Sekijima, Y.; Sipe, J.D.; Westermark, P. Amyloid nomenclature 2020: Update and recommendations by the International Society of Amyloidosis (ISA) nomenclature committee. Amyloid 2020, 27, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Bondarev, S.; Antonets, K.; Kajava, A.; Nizhnikov, A.; Zhouravleva, G. Protein co-aggregation related to amyloids: Methods of investigation, diversity, and classification. Int. J. Mol. Sci. 2018, 19, 2292. [Google Scholar] [CrossRef]
- Gómez-Benito, M.; Granado, N.; García-Sanz, P.; Michel, A.; Dumoulin, M.; Moratalla, R. Modeling Parkinson’s Disease With the Alpha-Synuclein Protein. Front. Pharmacol. 2020, 11, 356. [Google Scholar] [CrossRef] [PubMed]
- Mucibabic, M.; Steneberg, P.; Lidh, E.; Straseviciene, J.; Ziolkowska, A.; Dahl, U.; Lindahl, E.; Edlund, H. α-Synuclein promotes IAPP fibril formation in vitro and β-cell amyloid formation in vivo in mice. Sci. Rep. 2020, 10, 20438. [Google Scholar] [CrossRef] [PubMed]
- Chai, Y.J.; Sierecki, E.; Tomatis, V.M.; Gormal, R.S.; Giles, N.; Morrow, I.C.; Xia, D.; Götz, J.; Parton, R.G.; Collins, B.M.; et al. Munc18-1 is a molecular chaperone for α-synuclein, controlling its self-replicating aggregation. J. Cell Biol. 2016, 214, 705–718. [Google Scholar] [CrossRef] [PubMed]
- Jaffrey, S.R.; Snowman, A.M.; Eliasson, M.J.; Cohen, N.A.; Snyder, S.H. CAPON: A protein associated with neuronal nitric oxide synthase that regulates its interactions with PSD95. Neuron 1998, 20, 115–124. [Google Scholar] [CrossRef]
- Freudenberg, F.; Alttoa, A.; Reif, A. Neuronal nitric oxide synthase (NOS1) and its adaptor, NOS1AP, as a genetic risk factors for psychiatric disorders. Genes, Brain Behav. 2015, 14, 46–63. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Jin, L.; Zhu, Y.; Zhou, X.; Yu, R.; Gao, S. Research progress in NOS1AP in neurological and psychiatric diseases. Brain Res. Bull. 2016, 125, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Li, X.; Gao, S.; Niu, S.; Chen, M.; Qin, J.; Guo, Z.; Zhao, J.; Shen, A. Expression of CAPON after spinal cord injury in rats. J. Mol. Neurosci. 2008, 34, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.; Lv, Q.; Yan, M.; Cheng, C.; Guo, Z.; Yang, J.; Chen, M.; Xia, Y.; Zhang, L.; Shen, A. Elevated expression of CAPON and neuronal nitric oxide synthase in the sciatic nerve of rats following constriction injury. Vet. J. 2011, 187, 374–380. [Google Scholar] [CrossRef] [PubMed]
- Brzustowicz, L.M.; Simone, J.; Mohseni, P.; Hayter, J.E.; Hodgkinson, K.A.; Chow, E.W.; Bassett, A.S. Linkage disequilibrium mapping of schizophrenia susceptibility to the CAPON region of chromosome 1q22. Am. J. Hum. Genet. 2004, 74, 1057–1063. [Google Scholar] [CrossRef]
- Zheng, Y.; Li, H.; Qin, W.; Chen, W.; Duan, Y.; Xiao, Y.; Li, C.; Zhang, J.; Li, X.; Feng, G.; et al. Association of the carboxyl-terminal PDZ ligand of neuronal nitric oxide synthase gene with schizophrenia in the Chinese Han population. Biochem. Biophys. Res. Commun. 2005, 328, 809–815. [Google Scholar] [CrossRef]
- Miranda, A.; García, J.; López, C.; Gordon, D.; Palacio, C.; Restrepo, G.; Ortiz, J.; Montoya, G.; Cardeño, C.; Calle, J.; et al. Putative association of the carboxy-terminal PDZ ligand of neuronal nitric oxide synthase gene (CAPON) with schizophrenia in a Colombian population. Schizophr. Res. 2006, 82, 283–285. [Google Scholar] [CrossRef]
- Xu, B.; Wratten, N.; Charych, E.I.; Buyske, S.; Firestein, B.L.; Brzustowicz, L.M. Increased expression in dorsolateral prefrontal cortex of CAPON in schizophrenia and bipolar disorder. PLoS Med. 2005, 2, 999–1007. [Google Scholar] [CrossRef]
- Hadzimichalis, N.M.; Previtera, M.L.; Moreau, M.P.; Li, B.; Lee, G.H.; Dulencin, A.M.; Matteson, P.G.; Buyske, S.; Millonig, J.H.; Brzustowicz, L.M.; et al. NOS1AP protein levels are altered in BA46 and cerebellum of patients with schizophrenia. Schizophr. Res. 2010, 124, 248–250. [Google Scholar] [CrossRef]
- Oughtred, R.; Rust, J.; Chang, C.; Breitkreutz, B.J.; Stark, C.; Willems, A.; Boucher, L.; Leung, G.; Kolas, N.; Zhang, F.; et al. The BioGRID database: A comprehensive biomedical resource of curated protein, genetic, and chemical interactions. Protein Sci. Publ. Protein Soc. 2021, 30, 187–200. [Google Scholar] [CrossRef]
- Ahmed, A.B.; Znassi, N.; Château, M.T.; Kajava, A.V. A structure-based approach to predict predisposition to amyloidosis. Alzheimer’S Dement. 2015, 11, 681–690. [Google Scholar] [CrossRef]
- Kajava, A.V.; Baxa, U.; Steven, A.C. β arcades: Recurring motifs in naturally occurring and disease-related amyloid fibrils. Faseb J. 2010, 24, 1311–1319. [Google Scholar] [CrossRef]
- Bondarev, S.A.; Zhouravleva, G.A.; Belousov, M.V.; Kajava, A.V. Structure-based view on [PSI+] prion properties. Prion 2015, 9, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Roche, D.B.; Villain, E.; Kajava, A.V. Usage of a dataset of NMR resolved protein structures to test aggregation versus solubility prediction algorithms. Protein Sci. Publ. Protein Soc. 2017, 26, 1864–1869. [Google Scholar] [CrossRef]
- Dosztányi, Z.; Csizmók, V.; Tompa, P.; Simon, I. The pairwise energy content estimated from amino acid composition discriminates between folded and intrinsically unstructured proteins. J. Mol. Biol. 2005, 347, 827–839. [Google Scholar] [CrossRef]
- Dosztanyi, Z.; Csizmok, V.; Tompa, P.; Simon, I. IUPred: Web server for the prediction of intrinsically unstructured regions of proteins based on estimated energy content. Bioinformatics 2005, 21, 3433–3434. [Google Scholar] [CrossRef] [PubMed]
- Maurer-Stroh, S.; Debulpaep, M.; Kuemmerer, N.; De La Paz, M.L.; Martins, I.C.; Reumers, J.; Morris, K.L.; Copland, A.; Serpell, L.; Serrano, L.; et al. Exploring the sequence determinants of amyloid structure using position-specific scoring matrices. Nat. Methods 2010, 7, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Conchillo-Solé, O.; de Groot, N.S.; Avilés, F.X.; Vendrell, J.; Daura, X.; Ventura, S. AGGRESCAN: A server for the prediction and evaluation of “hot spots” of aggregation in polypeptides. BMC Bioinform. 2007, 8, 65. [Google Scholar] [CrossRef] [PubMed]
- Garbuzynskiy, S.O.; Lobanov, M.Y.; Galzitskaya, O.V. FoldAmyloid: A method of prediction of amyloidogenic regions from protein sequence. Bioinformatics 2009, 26, 326–332. [Google Scholar] [CrossRef]
- Rubel, M.S.; Fedotov, S.A.; Grizel, A.V.; Sopova, J.V.; Malikova, O.A.; Chernoff, Y.O.; Rubel, A.A. Functional mammalian amyloids and amyloid-like proteins. Life 2020, 10, 156. [Google Scholar] [CrossRef]
- Chandramowlishwaran, P.; Sun, M.; Casey, K.L.; Romanyuk, A.V.; Grizel, A.V.; Sopova, J.V.; Rubel, A.A.; Nussbaum-Krammer, C.; Vorberg, I.M.; Chernoff, Y.O. Mammalian amyloidogenic proteins promote prion nucleation in yeast. J. Biol. Chem. 2018, 293, 3436–3450. [Google Scholar] [CrossRef] [PubMed]
- Sivanathan, V.; Hochschild, A. A bacterial export system for generating extracellular amyloid aggregates. Nat. Protoc. 2013, 8, 1381–1390. [Google Scholar] [CrossRef] [PubMed]
- Alberti, S.; Halfmann, R.; King, O.; Kapila, A.; Lindquist, S. A systematic survey identifies prions and illuminates sequence features of prionogenic proteins. Cell 2009, 137, 146–158. [Google Scholar] [CrossRef] [PubMed]
- Liebman, S.W.; Chernoff, Y.O. Prions in yeast. Genetics 2012, 191, 1041–1072. [Google Scholar] [CrossRef]
- Muley, V.Y.; Akhter, Y.; Galande, S.; Gojobori, T. PDZ domains across the microbial world: Molecular link to the proteases, stress response, and protein synthesis. Genome Biol. Evol. 2019, 11, 644–659. [Google Scholar] [CrossRef] [PubMed]
- Kryndushkin, D.S.; Alexandrov, I.M.; Ter-Avanesyan, M.D.; Kushnirov, V.V. Yeast [PSI+] prion aggregates are formed by small Sup35 polymers fragmented by Hsp104. J. Biol. Chem. 2003, 278, 49636–49643. [Google Scholar] [CrossRef]
- Hein, M.Y.; Hubner, N.C.; Poser, I.; Cox, J.; Nagaraj, N.; Toyoda, Y.; Gak, I.A.; Weisswange, I.; Mansfeld, J.; Buchholz, F.; et al. A human interactome in three quantitative dimensions organized by stoichiometries and abundances. Cell 2015, 163, 712–723. [Google Scholar] [CrossRef]
- Chiti, F.; Dobson, C.M. Protein misfolding, amyloid formation, and human disease: A summary of progress over the last decade. Annu. Rev. Biochem. 2017, 86, 27–68. [Google Scholar] [CrossRef]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Liu, C.; Sawaya, M.R.; Eisenberg, D. β2-microglobulin forms three-dimensional domain-swapped amyloid fibrils with disulfide linkages. Nat. Struct. Mol. Biol. 2011, 18, 49–55. [Google Scholar] [CrossRef]
- Danilov, L.G.; Moskalenko, S.E.; Matveenko, A.G.; Sukhanova, X.V.; Belousov, M.V.; Zhouravleva, G.A.; Bondarev, S.A. The human NUP58 nucleoporin can form amyloids in vitro and in vivo. Biomedicines 2021, 9, 1451. [Google Scholar] [CrossRef]
- Eastwood, S.L. Does the CAPON gene confer susceptibility to schizophrenia? PLoS Med. 2005, 2, 946–947. [Google Scholar] [CrossRef]
- Moghaddam, B.; Javitt, D. From revolution to evolution: The glutamate hypothesis of schizophrenia and its implication for treatment. Neuropsychopharmacology 2012, 37, 4–15. [Google Scholar] [CrossRef]
- Winter, C.; Juckel, G.; Plotkin, M.; Niehaus, L.; Kupsch, A. Paranoid schizophrenia and idiopathic Parkinson’s disease do coexist: A challenge for clinicians. Psychiatry Clin. Neurosci. 2006, 60, 639. [Google Scholar] [CrossRef] [PubMed]
- Kuusimäki, T.; Al-Abdulrasul, H.; Kurki, S.; Hietala, J.; Hartikainen, S.; Koponen, M.; Tolppanen, A.M.; Kaasinen, V. Increased risk of Parkinson’s disease in patients with schizophrenia spectrum disorders. Mov. Disord. 2021, 36, 1353–1361. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, S.; Matsuba, Y.; Kamano, N.; Mihira, N.; Sahara, N.; Takano, J.; Muramatsu, S.i.; Saido, T.C.; Saito, T. Tau binding protein CAPON induces tau aggregation and neurodegeneration. Nat. Commun. 2019, 10, 2394. [Google Scholar] [CrossRef] [PubMed]
- Souza, J.M.; Giasson, B.I.; Chen, Q.; Lee, V.M.; Ischiropoulos, H. Dityrosine cross-linking promotes formation of stable α-synuclein polymers: Implication of nitrative and oxidative stress in the pathogenesis of neurodegenerative synucleinopathies. J. Biol. Chem. 2000, 275, 18344–18349. [Google Scholar] [CrossRef]
- Takahashi, T.; Yamashita, H.; Nakamura, T.; Nagano, Y.; Nakamura, S. Tyrosine 125 of α-synuclein plays a critical role for dimerization following nitrative stress. Brain Res. 2002, 938, 73–80. [Google Scholar] [CrossRef]
- Olivares, D.; Huang, X.; Branden, L.; Greig, N.H.; Rogers, J.T. Physiological and pathological role of alpha-synuclein in parkinson’s disease through iron mediated oxidative stress; the role of a putative iron-responsive element. Int. J. Mol. Sci. 2009, 10, 1226–1260. [Google Scholar] [CrossRef]
- Giasson, B.I.; Duda, J.E.; Murray, I.V.J.; Chen, Q.; Souza, J.M.; Hurtig, H.I.; Ischiropoulos, H.; Trojanowski, J.Q.-Y.; Lee, V.M. Oxidative damage linked to neurodegeneration by selective α-synuclein nitration in synucleinopathy lesions. Science 2000, 290, 985–989. [Google Scholar] [CrossRef]
- Hodara, R.; Norris, E.H.; Giasson, B.I.; Mishizen-Eberz, A.J.; Lynch, D.R.; Lee, V.M.; Ischiropoulos, H. Functional consequences of α-synuclein tyrosine nitration: Diminished binding to lipid vesicles and increased fibril formation. J. Biol. Chem. 2004, 279, 47746–47753. [Google Scholar] [CrossRef]
- Hanahan, D. Studies on transformation of Escherichia coli with plasmids. J. Mol. Biol. 1983, 166, 557–580. [Google Scholar] [CrossRef]
- Croucher, D.R.; Iconomou, M.; Hastings, J.F.; Kennedy, S.P.; Han, J.Z.; Shearer, R.F.; McKenna, J.; Wan, A.; Lau, J.; Aparicio, S.; et al. Bimolecular complementation affinity purification (BiCAP) reveals dimer-specific protein interactions for ERBB2 dimers. Sci. Signal. 2016, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Derkatch, I.L.; Bradley, M.; Zhou, P.; Chernoff, Y.O.; Liebman, S.W. Genetic and environmental factors affecting the de novo appearance of the [PSI+] prion in Saccharomyces cerevisiae. Genetics 1997, 147, 507–519. [Google Scholar] [CrossRef] [PubMed]
- Gietz, R.D.; Woods, R.A. Transformation of yeast by lithium acetate/single-stranded carrier DNA/polyethylene glycol method. Methods Enzymol. 2002, 350, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, C.; Michaelis, S.; Mitchell, A. Methods in Yeast Genetics; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1994; p. 234. [Google Scholar]
- Studier, F.W.; Moffatt, B.A. Use of bacteriophage T7 RNA polymerase to direct selective high-level expression of cloned genes. J. Mol. Biol. 1986, 189, 113–130. [Google Scholar] [CrossRef]
- Kushnirov, V.V.; Alexandrov, I.M.; Mitkevich, O.V.; Shkundina, I.S.; Ter-Avanesyan, M.D. Purification and analysis of prion and amyloid aggregates. Methods 2006, 39, 50–55. [Google Scholar] [CrossRef]
- Drozdova, P.B.; Barbitoff, Y.A.; Belousov, M.V.; Skitchenko, R.K.; Rogoza, T.M.; Leclercq, J.Y.; Kajava, A.V.; Matveenko, A.G.; Zhouravleva, G.A.; Bondarev, S.A. Estimation of amyloid aggregate sizes with semi-denaturing detergent agarose gel electrophoresis and its limitations. Prion 2020, 14, 118–128. [Google Scholar] [CrossRef]






| Primer | Sequence |
|---|---|
| NOS1AP-F-292-390 | GGGGACAAGTTTGTACAAAAAAGCAGGCTACCAGATGCAGCTCC |
| R_NOS1AP(HindIII)_attB2 | GGGGACCACTTTGTACAAGAAAGCTGGGTAAGCTTCTAGTCACAGAGCACGGGCAG |
| NOS1AP_1_291_NAR_attB1 forward_primer | GGGGACAAGTTTGTACAAAAAAGCAGGCTTCATGCCTAGCAAAACCAAG |
| NOS1AP_1_291_NAR_attB2 reverse_primer | GGGGACCACTTTGTACAAGAAAGCTGGGTTCTAGTGGTGAGTGGACAG |
| NOS1AP_391_506_NAR_attB1 forward_primer | GGGGACAAGTTTGTACAAAAAAGCAGGCTTCCCCACGACCCCTAAGCC |
| NOS1AP_391_506_ NAR_attB2 reverse_primer | GGGGACCACTTTGTACAAGAAAGCTGGGTTCTACACGGCGATCTCATC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matiiv, A.B.; Moskalenko, S.E.; Sergeeva, O.S.; Zhouravleva, G.A.; Bondarev, S.A. NOS1AP Interacts with α-Synuclein and Aggregates in Yeast and Mammalian Cells. Int. J. Mol. Sci. 2022, 23, 9102. https://doi.org/10.3390/ijms23169102
Matiiv AB, Moskalenko SE, Sergeeva OS, Zhouravleva GA, Bondarev SA. NOS1AP Interacts with α-Synuclein and Aggregates in Yeast and Mammalian Cells. International Journal of Molecular Sciences. 2022; 23(16):9102. https://doi.org/10.3390/ijms23169102
Chicago/Turabian StyleMatiiv, Anton B., Svetlana E. Moskalenko, Olga S. Sergeeva, Galina A. Zhouravleva, and Stanislav A. Bondarev. 2022. "NOS1AP Interacts with α-Synuclein and Aggregates in Yeast and Mammalian Cells" International Journal of Molecular Sciences 23, no. 16: 9102. https://doi.org/10.3390/ijms23169102
APA StyleMatiiv, A. B., Moskalenko, S. E., Sergeeva, O. S., Zhouravleva, G. A., & Bondarev, S. A. (2022). NOS1AP Interacts with α-Synuclein and Aggregates in Yeast and Mammalian Cells. International Journal of Molecular Sciences, 23(16), 9102. https://doi.org/10.3390/ijms23169102