
Attainment of Sexual Maturity and Gonadotropin Priming in Gilts Determine Follicular Development, Endocrine Milieu and Response to Ovulatory Triggers

 and
and 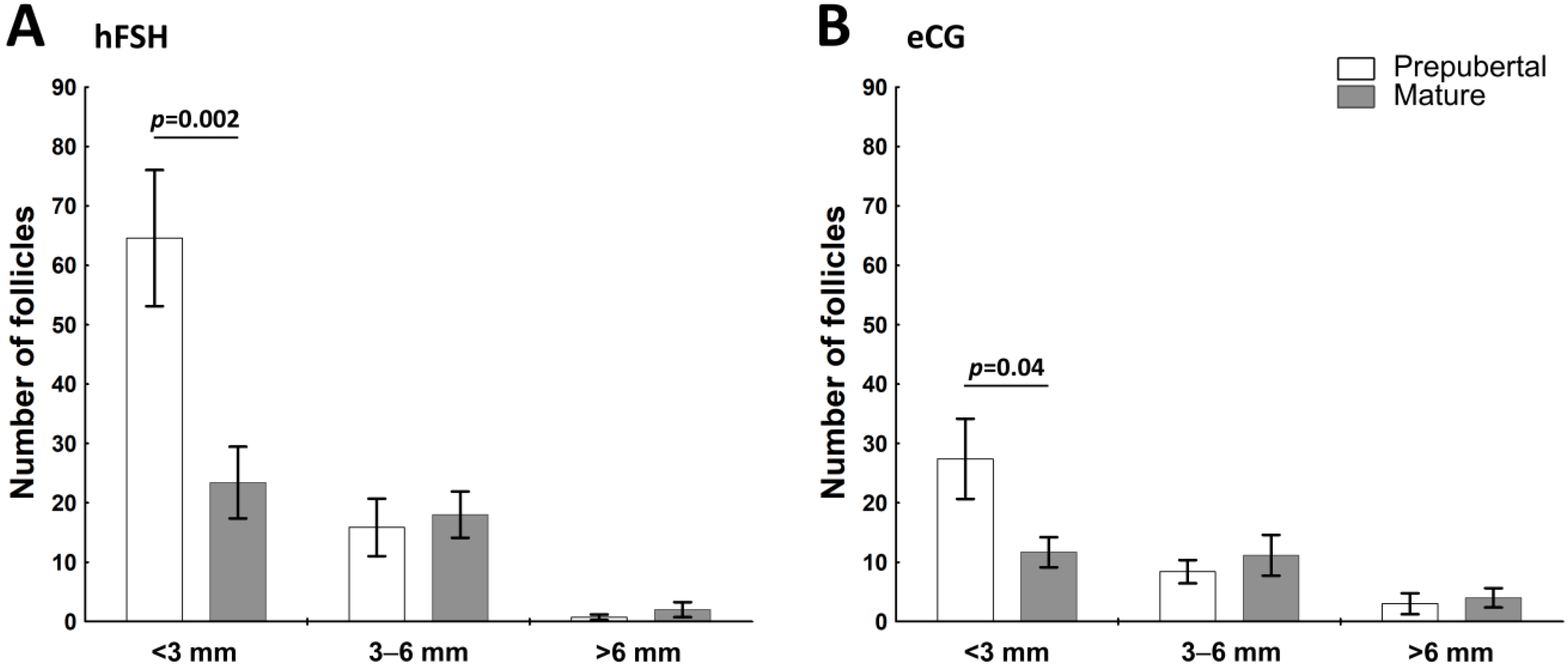{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. The Sexual Maturity Status Affects the Number of Small Visible Ovarian Follicles in hFSH- and eCG-Primed Gilts
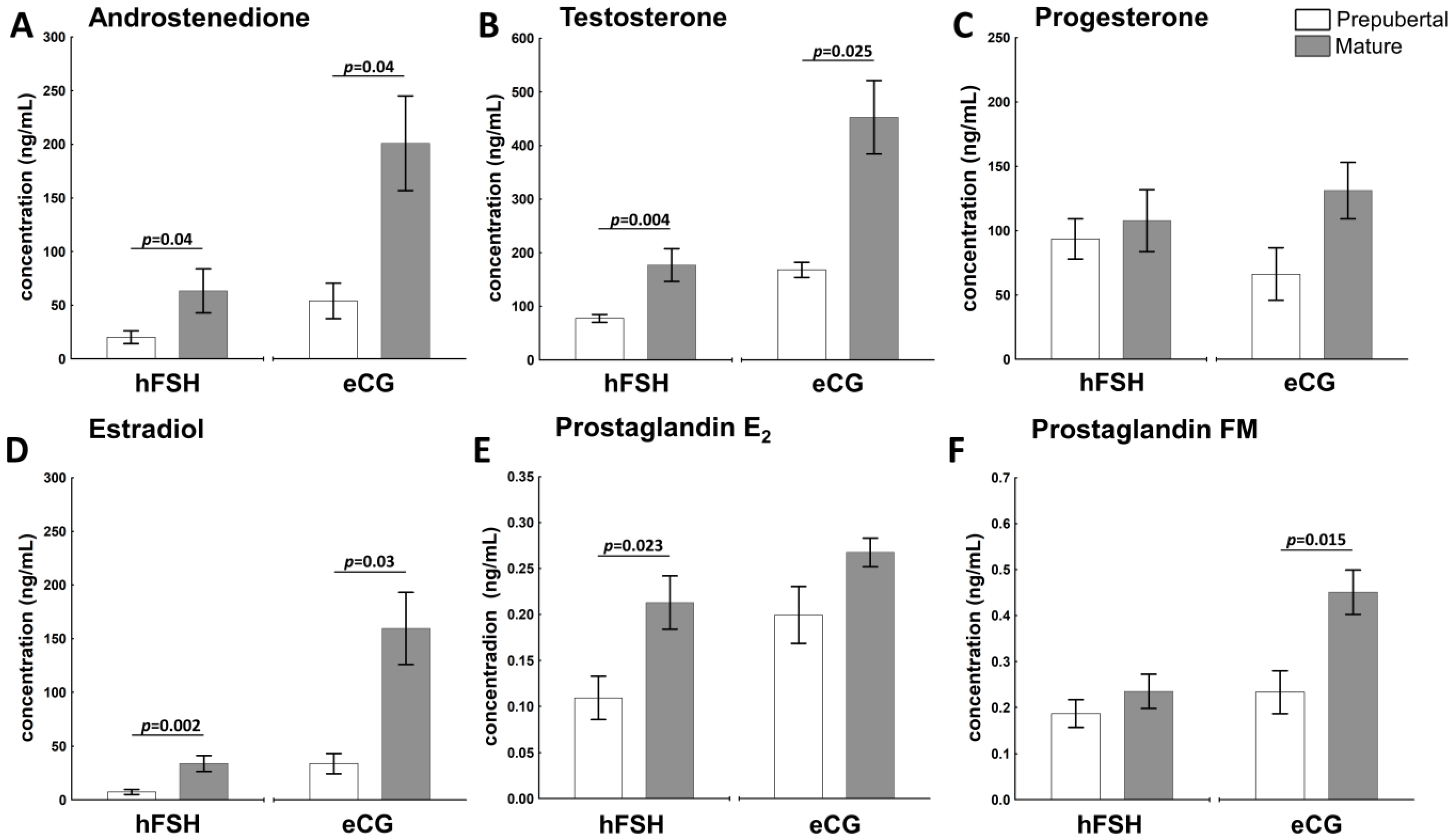
2.2. The Sexual Maturity Status Affects the Hormonal Milieu in Ovarian Follicles of hFSH- and eCG-Primed Gilts
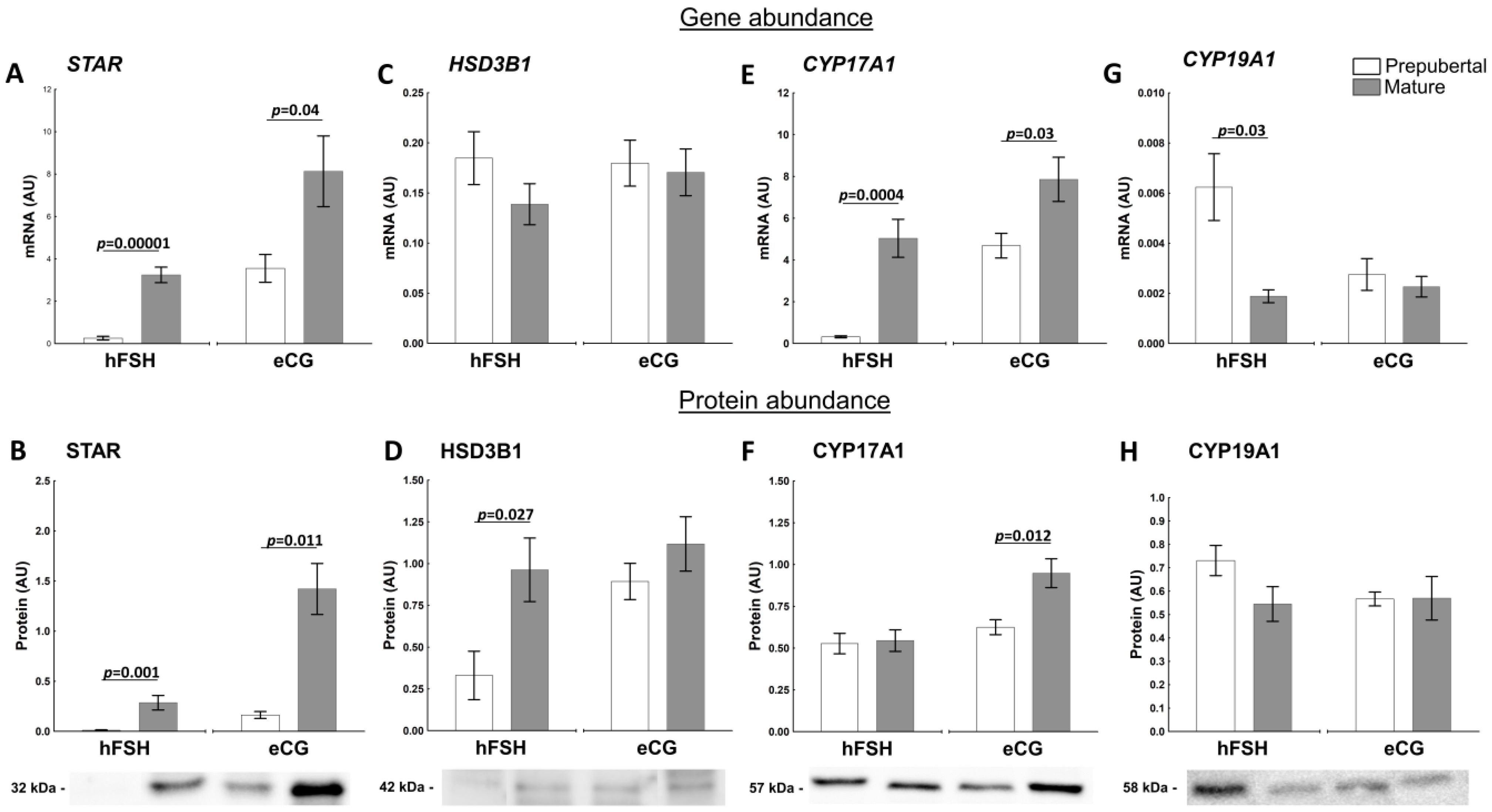
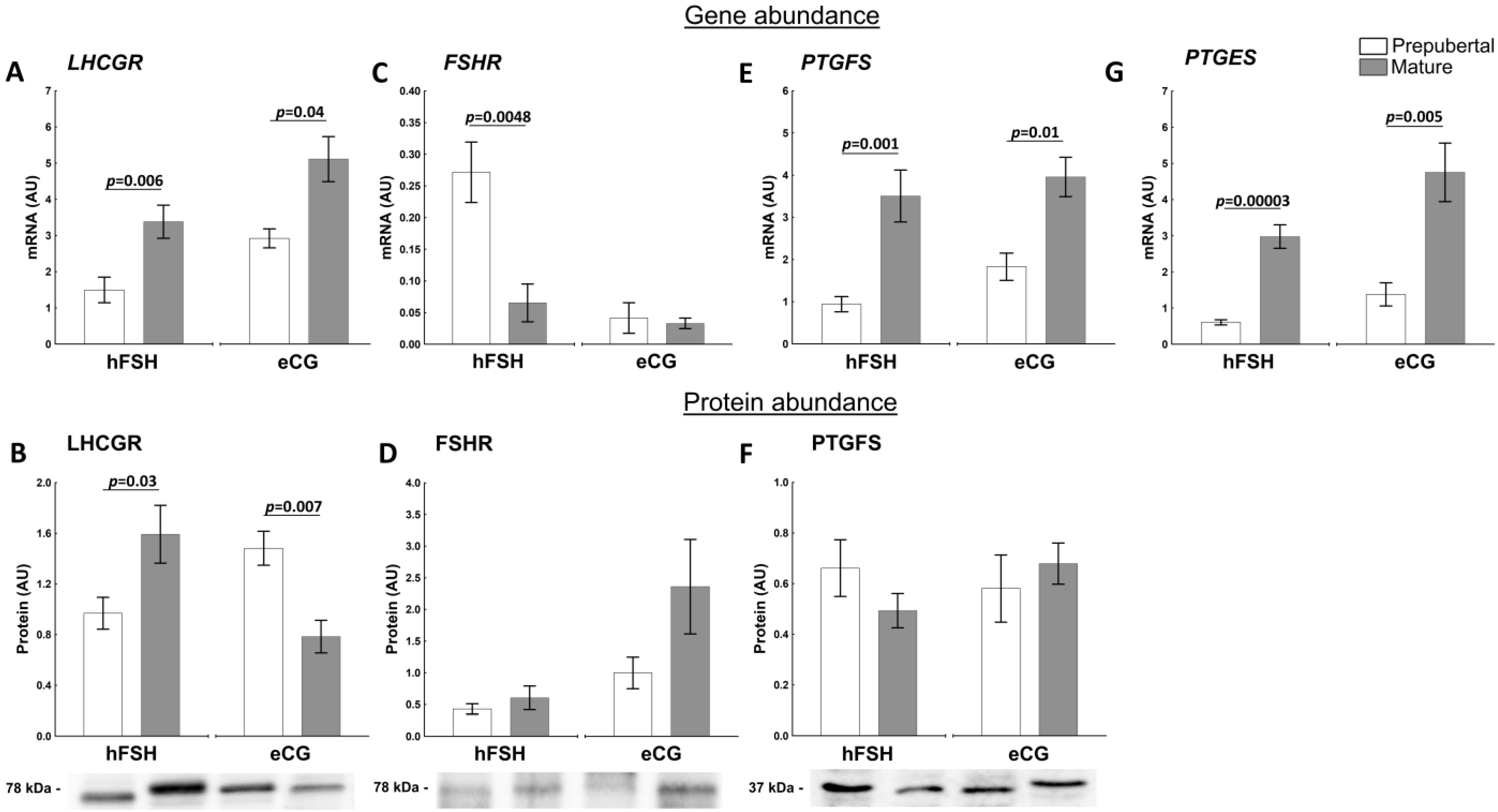
2.3. The Sexual Maturity Lead to Molecular Changes in Follicular Walls of Ovarian Follicles of hFSH- and eCG-Primed Gilts
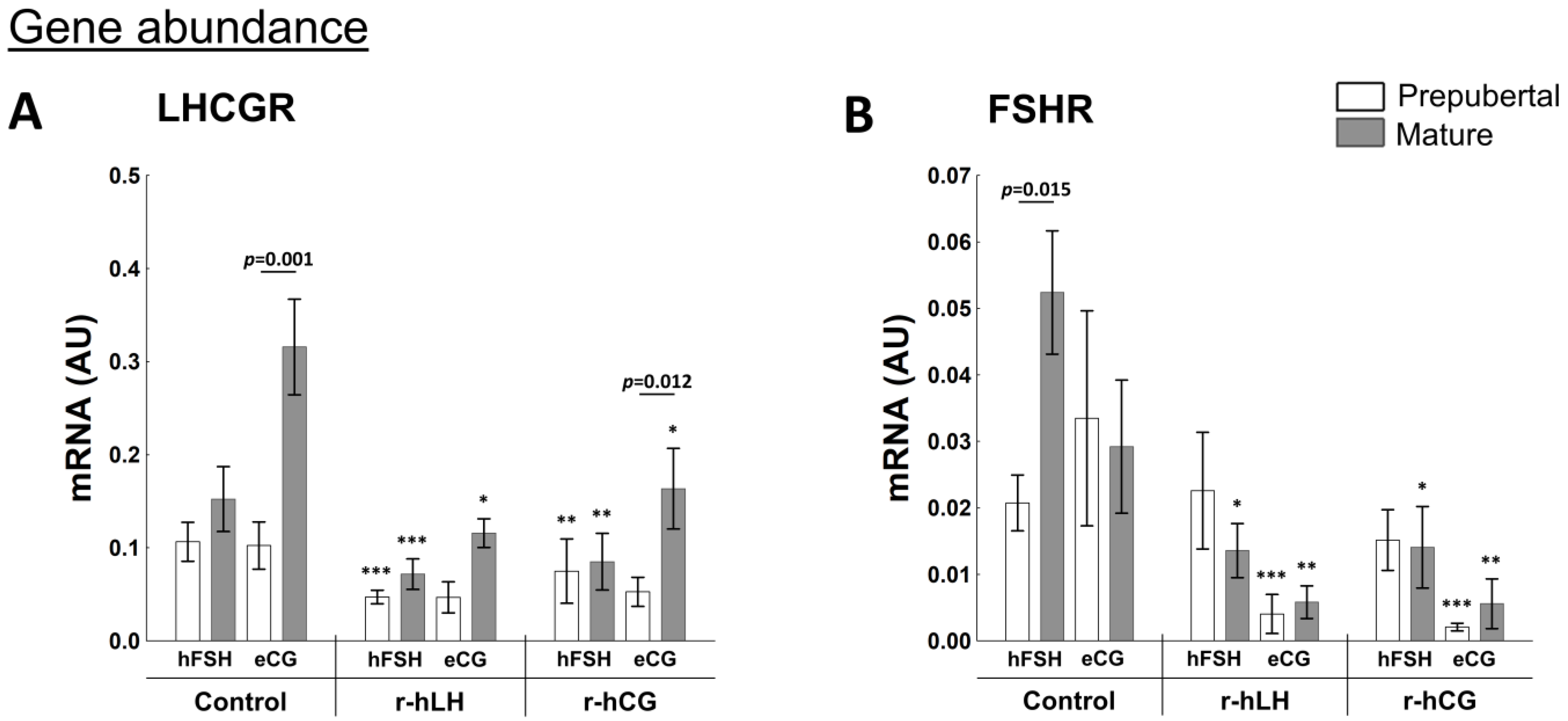
2.3.1. Factors Related to Progesterone and Estrogen Synthesis
2.3.2. Factors Related to the Production and Action of PGs
2.4. Effect of r-hLH and r-hCG In Vitro Stimulation on Molecular Changes in Follicular Walls of Prepubertal and Mature hFSH- and eCG-Primed Gilts
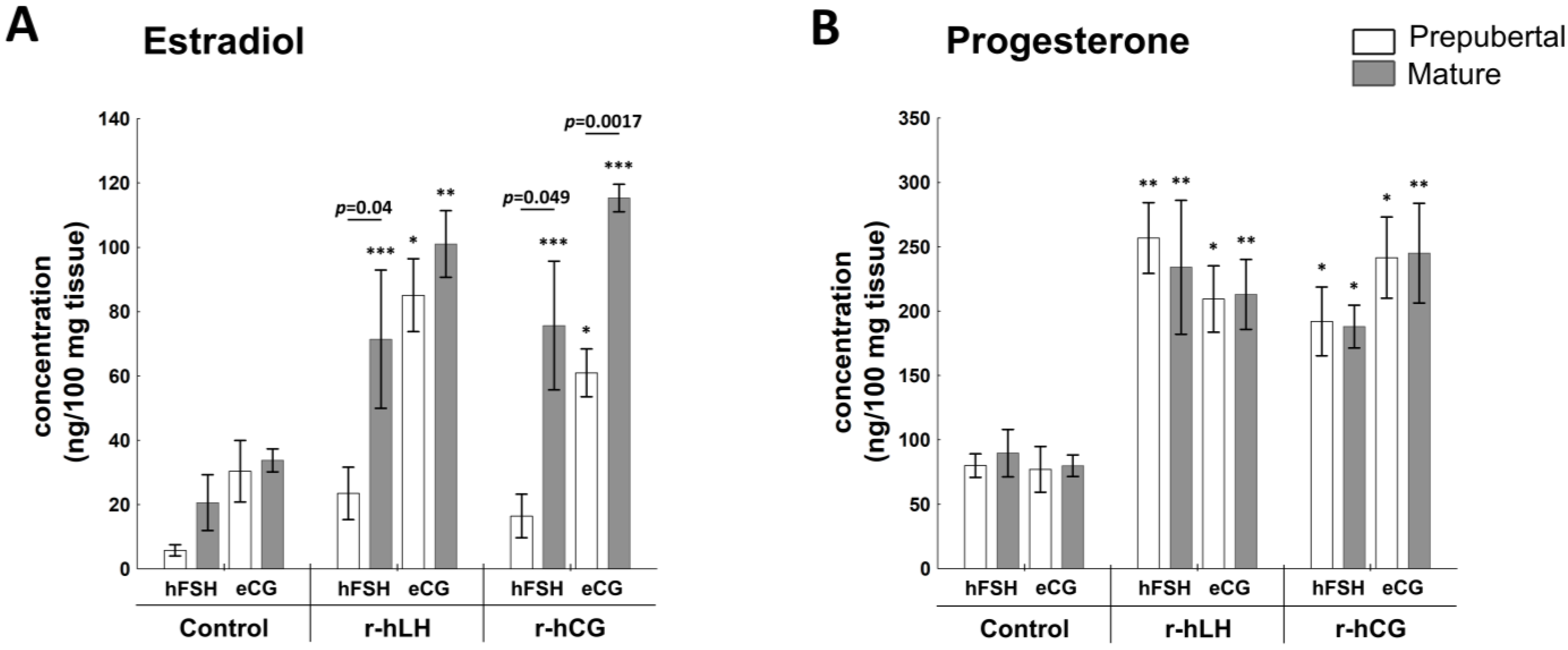
2.4.1. E2 and P4 Production
2.4.2. Receptors Related to Progesterone and Estrogen Synthesis
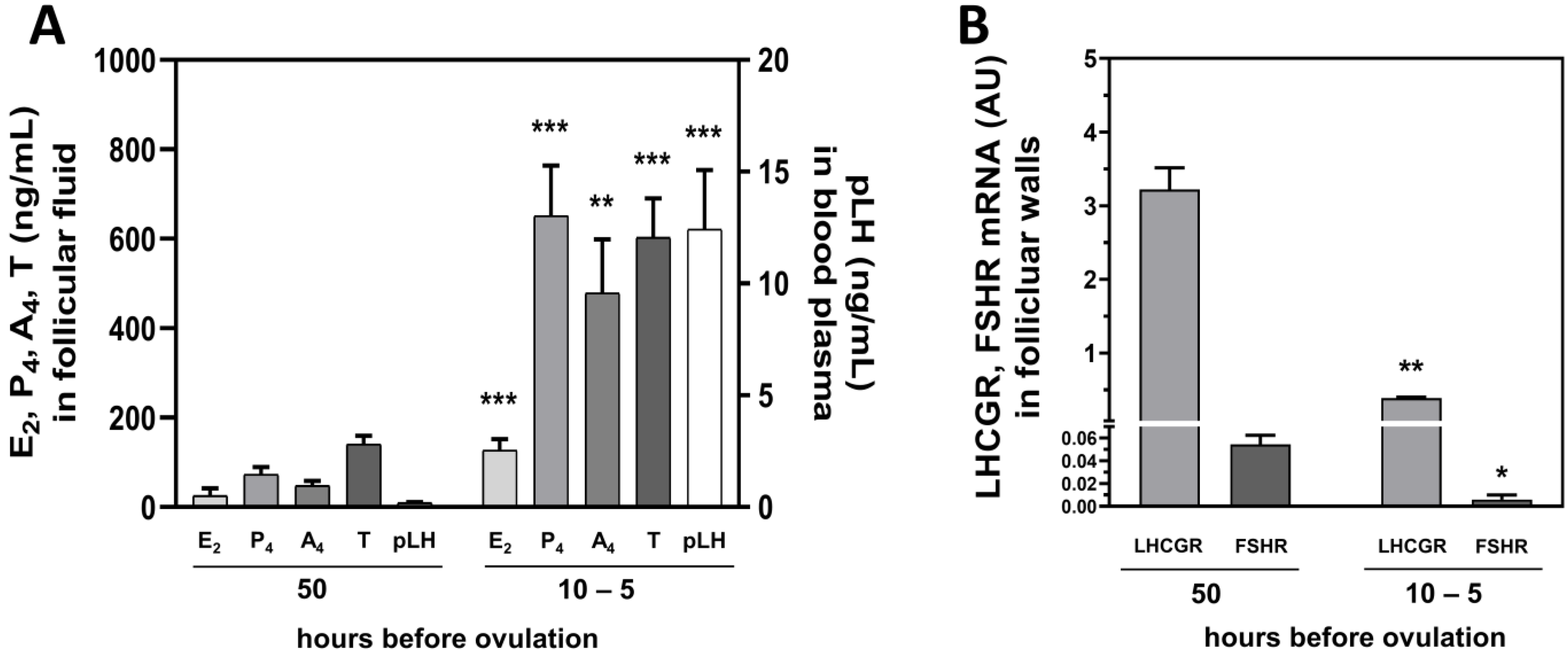
2.5. Hormonal Milieu and LHCGR and FSHR Levels in Follicular Walls Change before Ovulation
3. Discussion
4. Materials and Methods
4.1. Selection of Animals and Experimental Groups Recruitment
4.1.1. Experiment 1 [In Vivo]
4.1.2. Experiment 2 [In Vitro]
4.1.3. Experiment 3
4.2. Sample Collection
4.3. Steroid Hormones, Prostaglandins and pLH Assays
4.4. Protein Extraction
4.5. Western Blot
4.6. Total RNA Isolation and Real-Time PCR
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- LaVoie, H.A. Transcriptional control of genes mediating ovarian follicular growth, differentiation, and steroidogenesis in pigs. Mol. Reprod. Dev. 2017, 84, 788–801. [Google Scholar] [CrossRef] [PubMed]
- Tilton, J.E.; Foxcroft, G.R.; Ziecik, A.J.; Coombs, S.L.; Williams, G.L. Time of the preovulatory LH surge in the gilt and sow relative to the onset of behavioral estrus. Theriogenology 1982, 18, 227–236. [Google Scholar] [CrossRef]
- Soede, N.M.; Langendijk, P.; Kemp, B. Reproductive cycles in pigs. Anim. Reprod. Sci. 2011, 124, 251–258. [Google Scholar] [CrossRef] [PubMed]
- De Rensis, F.; Kirkwood, R.N. Control of estrus and ovulation: Fertility to timed insemination of gilts and sows. Theriogenology 2016, 86, 1460–1466. [Google Scholar] [CrossRef]
- Kraeling, R.P.; Dziuk, P.J.; Pursel, V.G.; Rampacek, G.B.; Webel, S.K. Synchronization of estrus in swine with allyl trenbolone (RU-2267). J. Anim. Sci. 1981, 52, 831–835. [Google Scholar] [CrossRef]
- Dura, M.; Tilton, J.E.; Ziecik, A.J.; Weigl, R.M.; Williams, G.L. Use of altrenogest alone or in combination with PMSG to control the preovulatory LH surge in gilts. Anim. Reprod. Sci. 1987, 14, 147–155. [Google Scholar] [CrossRef]
- Guthrie, H.D.; Bolt, D.J.; Cooper, B.S. Effects of gonadotropin treatment on ovarian follicle growth and granulosal cell aromatase activity in prepuberal gilts. J. Anim Sci. 1990, 68, 3719–3726. [Google Scholar] [CrossRef] [PubMed]
- Bolamba, D.; Dubuc, A.; Dufour, J.J.; Sirard, M.A. Effects of gonadotropin treatment on ovarian follicle growth, oocyte quality and in vitro fertilization of oocytes in prepubertal gilts. Theriogenology 1996, 46, 717–726. [Google Scholar] [CrossRef]
- Driancourt, M.A. Fixed time artificial insemination in gilts and sows. Tools, schedules and efficiency. In Control of Pig Reproduction IX. Context Products; Rodriguez-Martinez, H., Soede, N.M., Flowers, W.L., Eds.; Context Products Ltd.: Leicestershire, UK, 2013; pp. 89–99. [Google Scholar]
- Britt, J.H.; Day, B.N.; Webel, S.K.; Brauer, M.A. Induction of fertile estrus in prepuberal gilts by treatment with a combination of pregnant mares serum gonadotropin and human chorionic-gonadotropin. J. Anim. Sci. 1989, 67, 1148–1153. [Google Scholar] [CrossRef]
- Tummaruk, P.; Tantasuparuk, W.; Techakumphu, M.; Kunavongkrit, A. Age, body weight and backfat thickness at first observed oestrus in crossbred Landrace x Yorkshire gilts, seasonal variations and their influence on subsequence reproductive performance. Anim. Reprod. Sci. 2007, 99, 167–181. [Google Scholar] [CrossRef]
- Ziecik, A.J.; Klos, J.; Przygrodzka, E.; Milewski, R.; Jana, B. Aberrant effects of altrenogest and exposure to exogenous gonadotropins on follicular cysts appearance in gilts. Theriogenology 2017, 89, 250–254. [Google Scholar] [CrossRef] [PubMed]
- Ziecik, A.J.; Drzewiecka, K.; Gromadzka-Hliwa, K.; Klos, J.; Witek, P.; Knapczyk-Stwora, K.; Gajewski, Z.; Kaczmarek, M.M. Altrenogest affects the development and endocrine milieu of ovarian follicles in prepubertal and mature gilts. Biol. Reprod. 2020, 103, 1069–1084. [Google Scholar] [CrossRef] [PubMed]
- Ziecik, A.J.; Klos, J.; Gromadzka-Hliwa, K.; Dietrich, M.A.; Slowinska, M.; Likszo, P.; Knapczyk-Stwora, K.; Gajewski, Z.; Kaczmarek, M.M. Endocrine and molecular milieus of ovarian follicles are diversely affected by human chorionic gonadotropin and gonadotropin-releasing hormone in prepubertal and mature gilts. Sci. Rep. 2021, 11, 13465. [Google Scholar] [CrossRef] [PubMed]
- Allen, W.R.; Moor, R.M. The origin of the equine endometrial cups. I. Production of PMSG by fetal trophoblast cells. J. Reprod. Fertil. 1972, 29, 313–316. [Google Scholar] [CrossRef] [PubMed]
- Moore, W.T., Jr.; Ward, D.N. Pregnant mare serum gonadotropin. Rapid chromatographic procedures for the purification of intact hormone and isolation of subunits. J. Biol. Chem. 1980, 255, 6923–6929. [Google Scholar] [CrossRef]
- Sugino, H.; Bousfield, G.R.; Moore, W.T., Jr.; Ward, D.N. Structural studies on equine glycoprotein hormones. Amino acid sequence of equine chorionic gonadotropin beta-subunit. J. Biol. Chem. 1987, 262, 8603–8609. [Google Scholar] [CrossRef]
- Martinuk, S.D.; Manning, A.W.; Black, W.D.; Murphy, B.D. Effects of carbohydrates on the pharmacokinetics and biological activity. Biol. Reprod. 1991, 45, 598–604. [Google Scholar] [CrossRef]
- Schams, D.; Mentzer, C.; Schallenberger, E.; Hahn, J.; Hahn, R. Superovulating cows with follicle stimulating hormone and pregnant mare’s serum gonadotrophin. Theriogenology 1978, 9, 17–26. [Google Scholar]
- Dybala, J.A. Effect of Repeated Use of Gonadotropins on Reproductive Performance of Sows in Different Animal Welfare Commitments. Ph.D. Thesis, Institute of Animal Reprooduction and Food Research, Polish Academy of Sciences, Olsztyn, Poland, 2000. [Google Scholar]
- Murphy, B.D. Equine chorionic gonadotropin: An enigmatic but essential tool. Anim. Reprod. 2012, 9, 223–230. [Google Scholar]
- Foxcroft, G.R.; Hunter, M.G. Basic physiology of follicular maturation in the pig. J. Reprod. Fertil. Suppl. 1985, 33, 1–19. [Google Scholar]
- Karalus, U.; Downey, B.R.; Ainsworth, L. Maintenance of ovulatory cycles and pregnancy in prepubertal gilts treated with PMSG and hCG. Anim. Reprod. Sci. 1990, 22, 235–241. [Google Scholar] [CrossRef]
- Knox, R.V.; Tudor, K.W. Influence of norgestomet in combination with gonadotropins on induction of estrus and ovulation in prepubertal gilts. J. Anim. Sci. 1999, 77, 1348–1352. [Google Scholar] [CrossRef] [PubMed]
- Tilton, S.L.; Bates, R.O.; Prather, R.S. Evaluation of response to hormonal therapy in prepubertal gilts of different genetic lines. J. Anim. Sci. 1995, 73, 3062–3068. [Google Scholar] [CrossRef] [PubMed]
- do Lago, V.; Vianna, W.; Gama, R.; Campos Rosseto, A.; Pinese, M.; Sant’anna Moretti, A. Second oestrus synchronization and precocious embryo viability after puberty induction in gilts by the use of gonadotrophin treatment. Reprod. Domest. Anim. 2005, 40, 141–144. [Google Scholar] [CrossRef] [PubMed]
- Añazco, J.; Vinueza, N. Desarrollo de las estructuras ováricas en respuesta a la aplicación de dos gonadotropinas supeovulatorias en bovinos (PMSG y FSH-P). Carrera Ing. Agropecu. 2017. Available online: https://repositorio.espe.edu.ec/handle/21000/13423 (accessed on 17 January 2022).
- Flowers, B.; Martin, M.J.; Cantley, T.C.; Day, B.N. The effect of pregnant mare serum gonadotropin on follicle stimulating hormone and estradiol secretion in the prepuberal gilt. Anim. Reprod. Sci. 1989, 21, 93–100. [Google Scholar] [CrossRef]
- Hu, J.; Zhang, Z.; Shen, W.; Azhar, S. Cellular cholesterol delivery, intracellular processing and utilization for biosynthesis of steroid hormones. Nutr. Metab. 2010, 7, 47–72. [Google Scholar] [CrossRef]
- Conley, A.J.; Kaminski, M.A.; Dubowsky, S.A.; Jablonka-Shariff, A.; Redmer, D.A.; Reynolds, L.P. Immunohistochemical localization of 3 beta-hydroxysteroid dehydrogenase and P450 17 alpha-hydroxylase during follicular and luteal development in pigs, sheep, and cows. Biol. Reprod. 1995, 52, 1081–1094. [Google Scholar] [CrossRef]
- Ainsworth, L.; Tsang, B.K.; Downey, B.R.; Marcus, G.J. The synthesis and actions of steroids and prostaglandins during follicular maturation in the pig. J. Reprod. Fertil. Suppl. 1950, 40, 137–150. [Google Scholar] [CrossRef]
- Evans, G.; Dobias, M.; King, G.J.; Armstrong, D.T. Production of prostaglandins by porcine preovulatory follicular tissues and their roles in intrafollicular function. Biol. Reprod. 1983, 28, 322–328. [Google Scholar] [CrossRef]
- Lucy, M.C.; Liu, J.; Boyd, C.K.; Bracken, C. Ovarian folliCular growth in sows. Reprod. Suppl. 2001, 58, 31–45. [Google Scholar] [CrossRef] [PubMed]
- Knox, R.V. Recruitment and selection of ovarian follicles for determination of ovulation rate in the pig. Domest. Anim. Endocrinol. 2005, 29, 385–397. [Google Scholar] [CrossRef] [PubMed]
- Layman, L.C.; McDonough, P.G. Mutations of follicle stimulating hormone-beta and its receptor in human and mouse: Genotype/phenotype. Mol. Cell Endocrinol. 2000, 161, 9–17. [Google Scholar] [CrossRef]
- Zhang, C.; Xia, G.; Tsang, B.K. Interactions of thyroid hormone and FSH in the regulation of rat granulosa cell apoptosis. Front. Biosci. 2011, 1, 1401–1413. [Google Scholar] [CrossRef]
- Asahara, S.; Sato, A.; Aljonaid, A.A.; Maruo, T. Thyroid hormone synergizes with follicle stimulating hormone to inhibit apoptosis in porcine granulosa cells selectively from small follicles. Kobe J. Med. Sci. 2003, 49, 107–116. [Google Scholar] [PubMed]
- Guthrie, H.D.; Garrett, W.M. Apoptosis during folliculogenesis in pigs. Reprod. Suppl. 2001, 58, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.; Rui, R. Effects of follicular size and FSH on granulosa cell apoptosis and atresia in porcine antral follicles. Mol. Reprod. Dev. 2010, 77, 670–678. [Google Scholar] [CrossRef]
- Garrett, W.M.; Guthrie, H.D. Steroidogenic enzyme expression during preovulatory follicle maturation in pigs. Biol. Reprod. 1997, 56, 1424–1431. [Google Scholar] [CrossRef]
- Hafez, E.S.E. Reproduction in Farm Animals; Lea & Febiger: Philadelphia, PA, USA, 1974; pp. 212–221. [Google Scholar]
- Martinat-Botte, F.; Venturi, E.; Guillouet, P.; Driancourt, M.A.; Terqui, M. Induction and synchronization of ovulations of nulliparous and multiparous sows with an injection of gonadotropin-releasing hormone agonist (Receptal). Theriogenology 2010, 73, 332–342. [Google Scholar] [CrossRef]
- Blitek, A.; Waclawik, A.; Kaczmarek, M.M.; Kiewisz, J.; Ziecik, A.J. Effect of estrus induction on prostaglandin content and prostaglandin synthesis enzyme expression in the uterus of early pregnant pigs. Theriogenology 2010, 73, 1244–1256. [Google Scholar] [CrossRef]
- Ziecik, A.; Tilton, J.E.; Espana, F.; Weigl, R. Effect of human chorionic gonadotropin on preovulatory luteinizing hormone surge and ovarian hormone secretion in gilts. J. Anim. Sci. 1987, 64, 1134–1143. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Bonomi, M.; Busnelli, M.; Persani, L.; Vassart, G.; Costagliola, S. Structural differences in the hinge region of the glycoprotein hormone receptors: Evidence from the sulfated tyrosine residues. Mol. Endocrinol. 2006, 20, 3351–3363. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Fernald, R.D. Comprehensive algorithm for quantitative real-time polymerase chain reaction. J. Comput. Biol. 2005, 12, 1047–1064. [Google Scholar] [CrossRef]
- Andersen, C.; Jensen, J.L.; Ørntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Likszo, P.; Gromadzka-Hliwa, K.; Klos, J.; Kaczmarek, M.M.; Ziecik, A.J. Attainment of Sexual Maturity and Gonadotropin Priming in Gilts Determine Follicular Development, Endocrine Milieu and Response to Ovulatory Triggers. Int. J. Mol. Sci. 2022, 23, 9190. https://doi.org/10.3390/ijms23169190
Likszo P, Gromadzka-Hliwa K, Klos J, Kaczmarek MM, Ziecik AJ. Attainment of Sexual Maturity and Gonadotropin Priming in Gilts Determine Follicular Development, Endocrine Milieu and Response to Ovulatory Triggers. International Journal of Molecular Sciences. 2022; 23(16):9190. https://doi.org/10.3390/ijms23169190
Chicago/Turabian StyleLikszo, Pawel, Katarzyna Gromadzka-Hliwa, Jan Klos, Monika M. Kaczmarek, and Adam J. Ziecik. 2022. "Attainment of Sexual Maturity and Gonadotropin Priming in Gilts Determine Follicular Development, Endocrine Milieu and Response to Ovulatory Triggers" International Journal of Molecular Sciences 23, no. 16: 9190. https://doi.org/10.3390/ijms23169190
APA StyleLikszo, P., Gromadzka-Hliwa, K., Klos, J., Kaczmarek, M. M., & Ziecik, A. J. (2022). Attainment of Sexual Maturity and Gonadotropin Priming in Gilts Determine Follicular Development, Endocrine Milieu and Response to Ovulatory Triggers. International Journal of Molecular Sciences, 23(16), 9190. https://doi.org/10.3390/ijms23169190