
Prostate Cancer Secretome and Membrane Proteome from Pten Conditional Knockout Mice Identify Potential Biomarkers for Disease Progression

 ,
, 
 ,
,  and
and
Abstract
:1. Introduction
2. Results
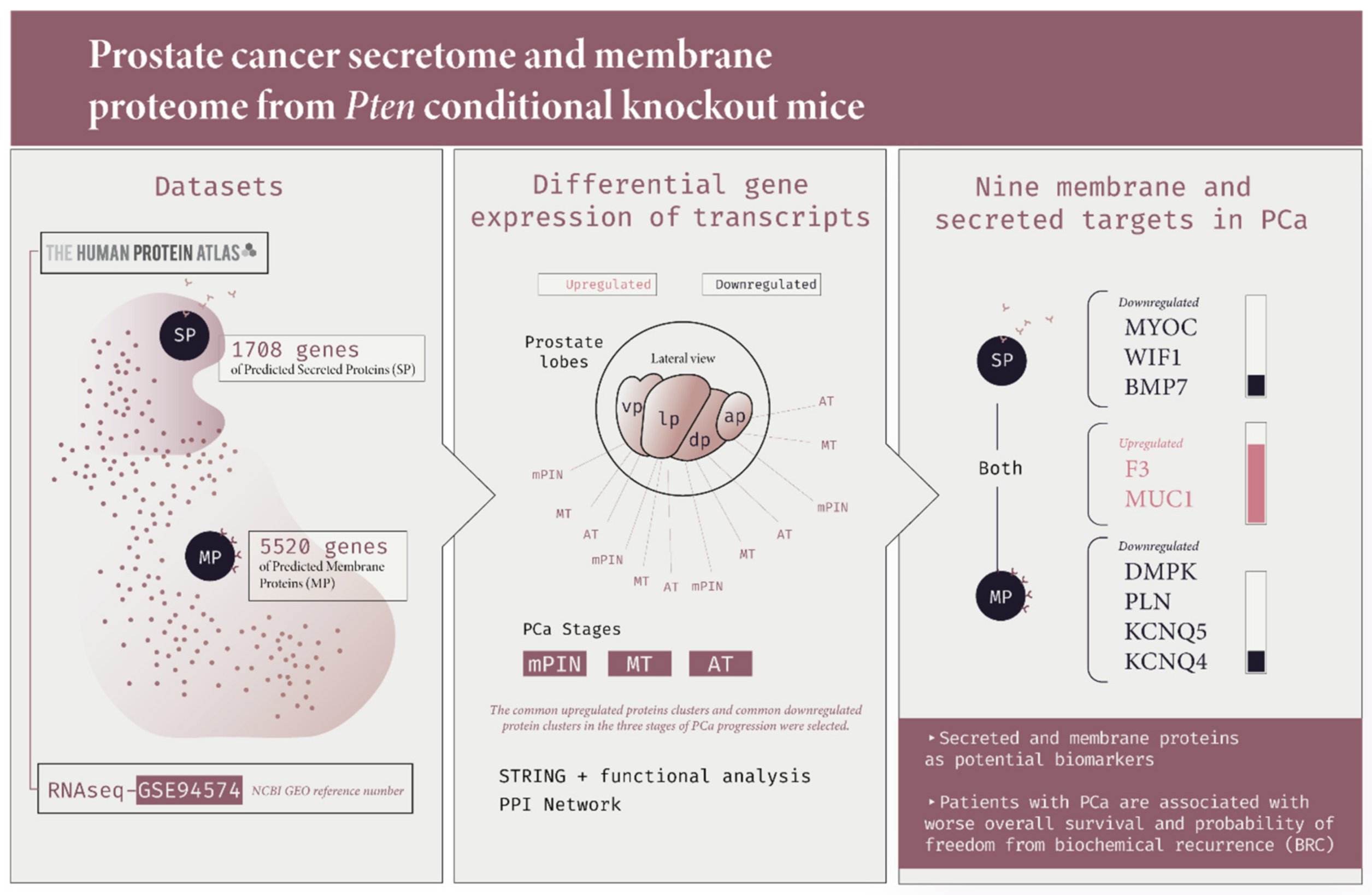
2.1. Identification of Gene Expression Profile in Prostate Cancer of Pb-Cre4/Ptenf/f Mice
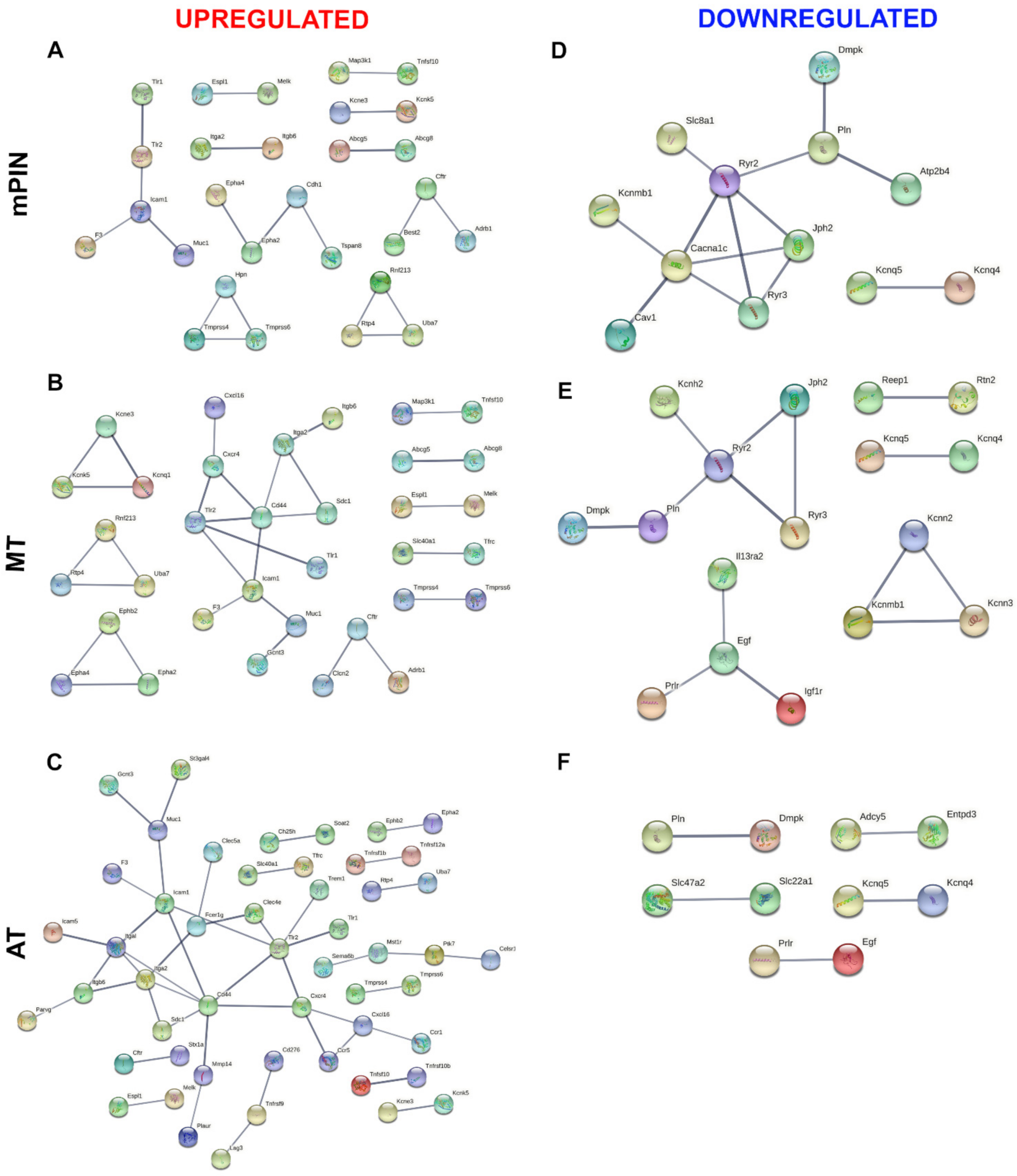
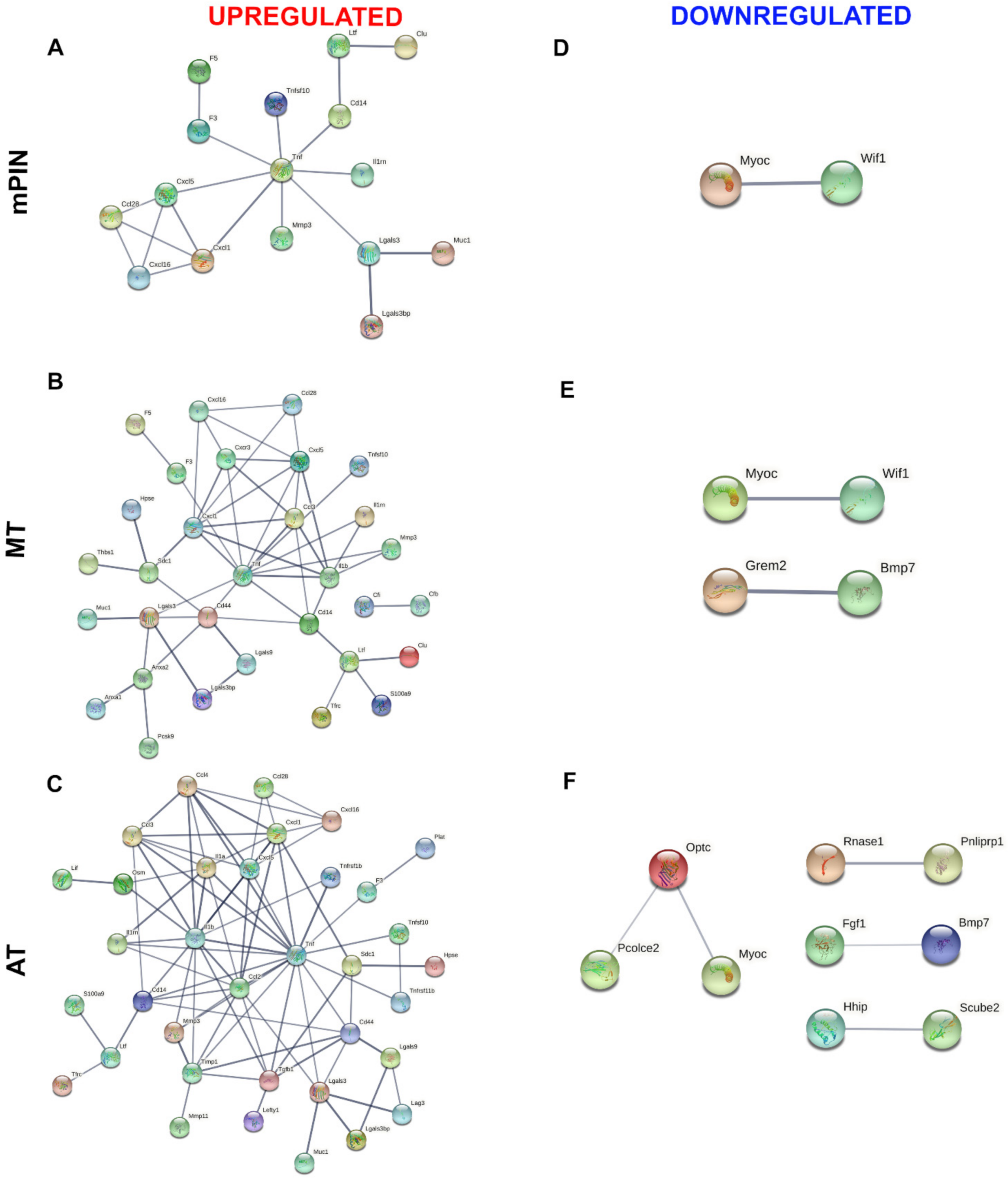
2.2. Protein–Protein Interaction (PPI) Network of Membrane and Secreted Proteins Enriched in Prostate Cancer
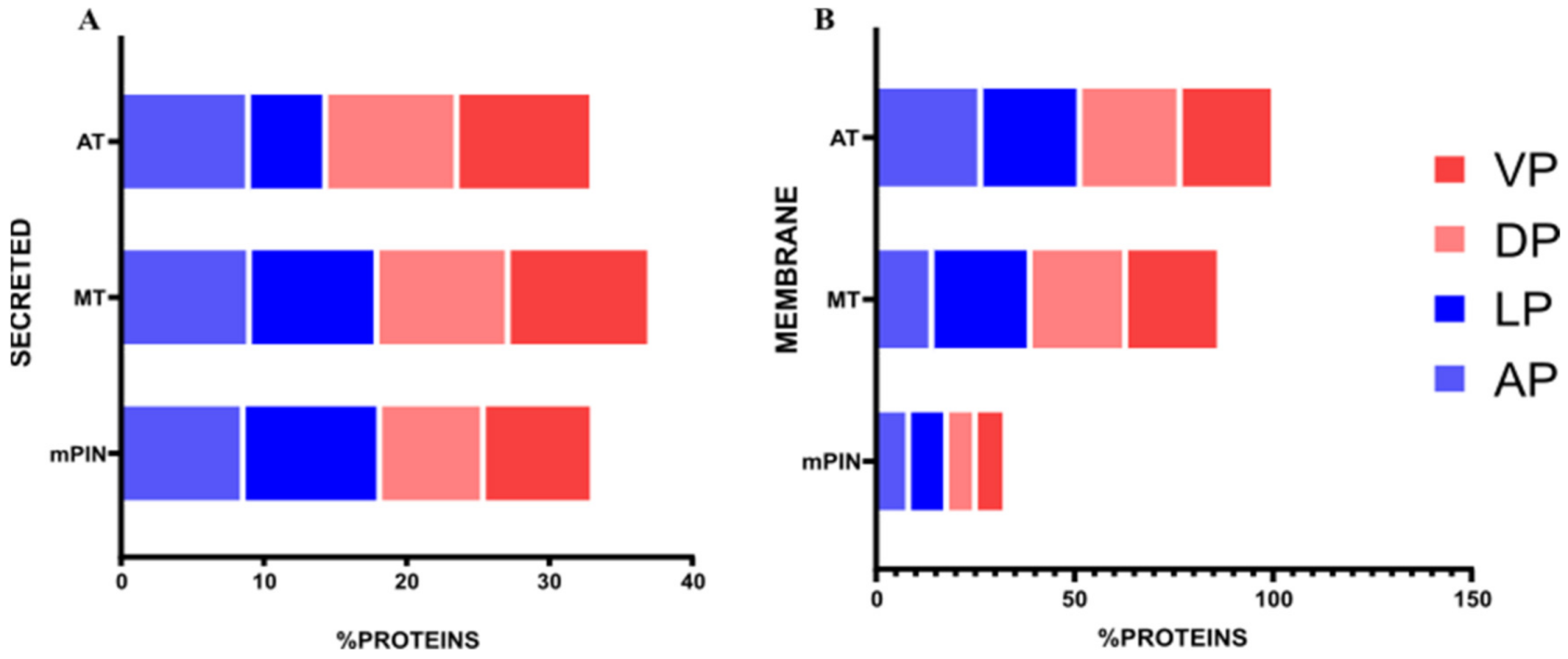
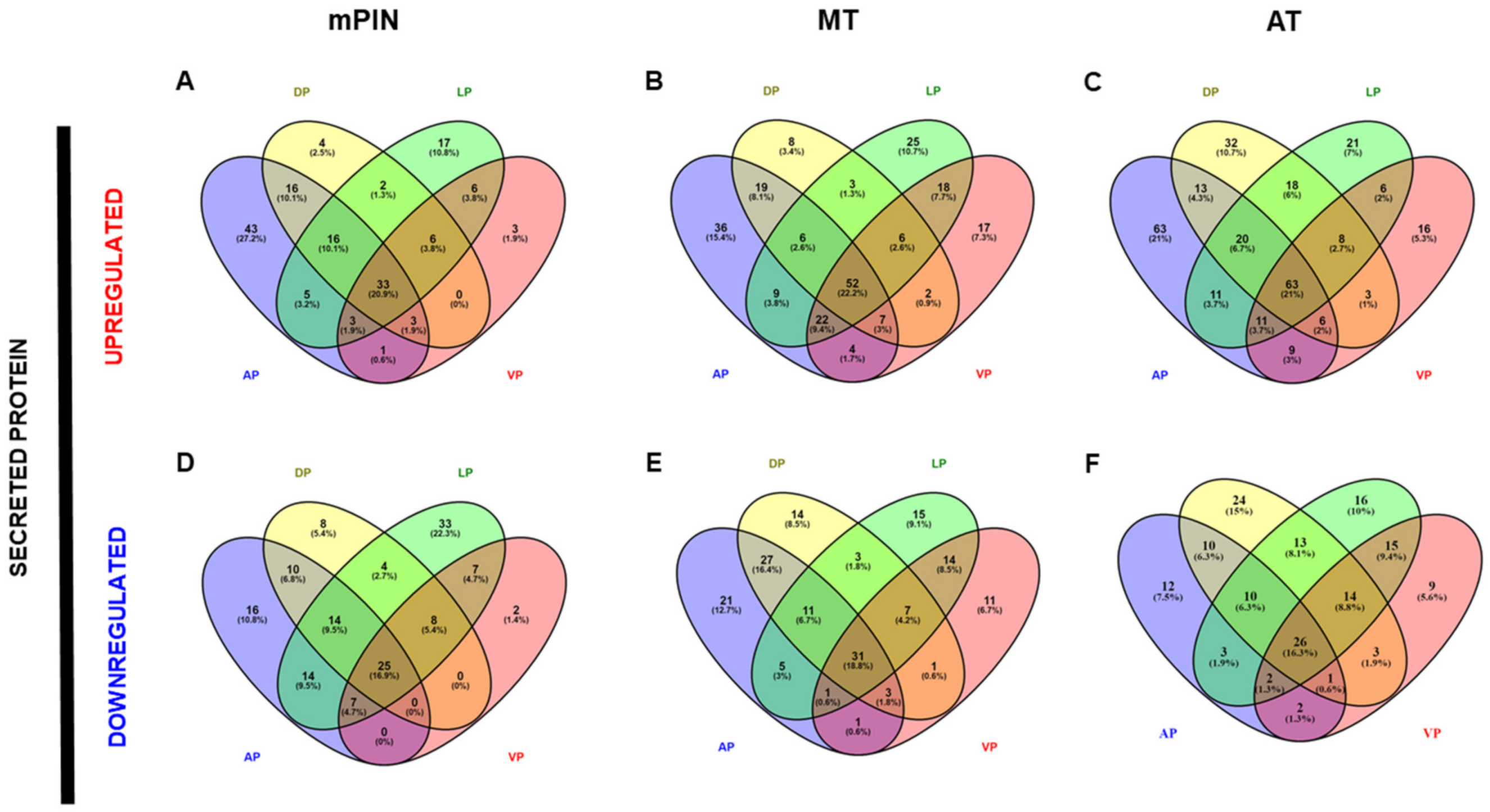
2.3. Differential Gene Expression of Transcripts Translated into The Membrane and Secreted Proteins in Prostate Cancer
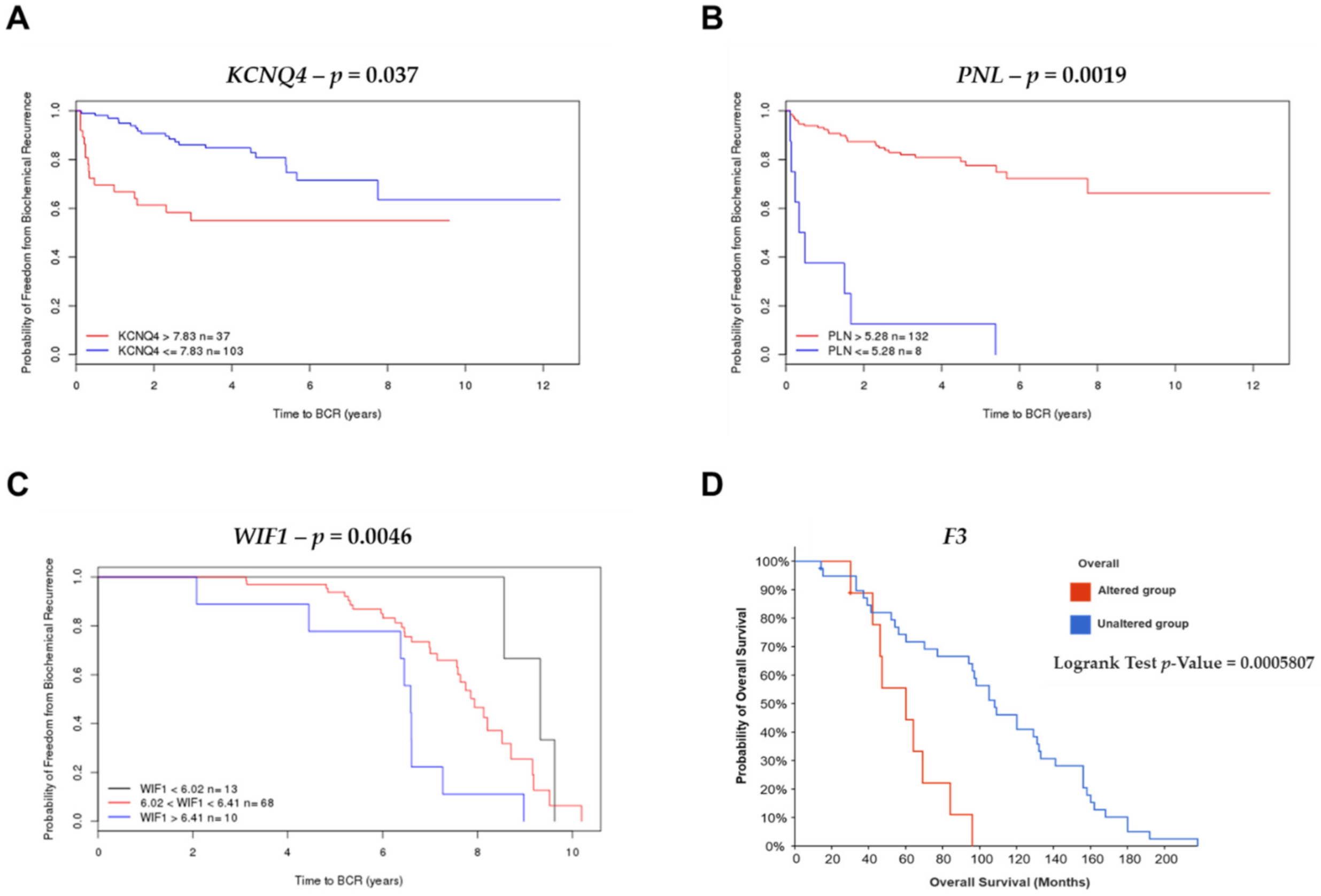
2.4. Survival Analysis and Risk Assessment
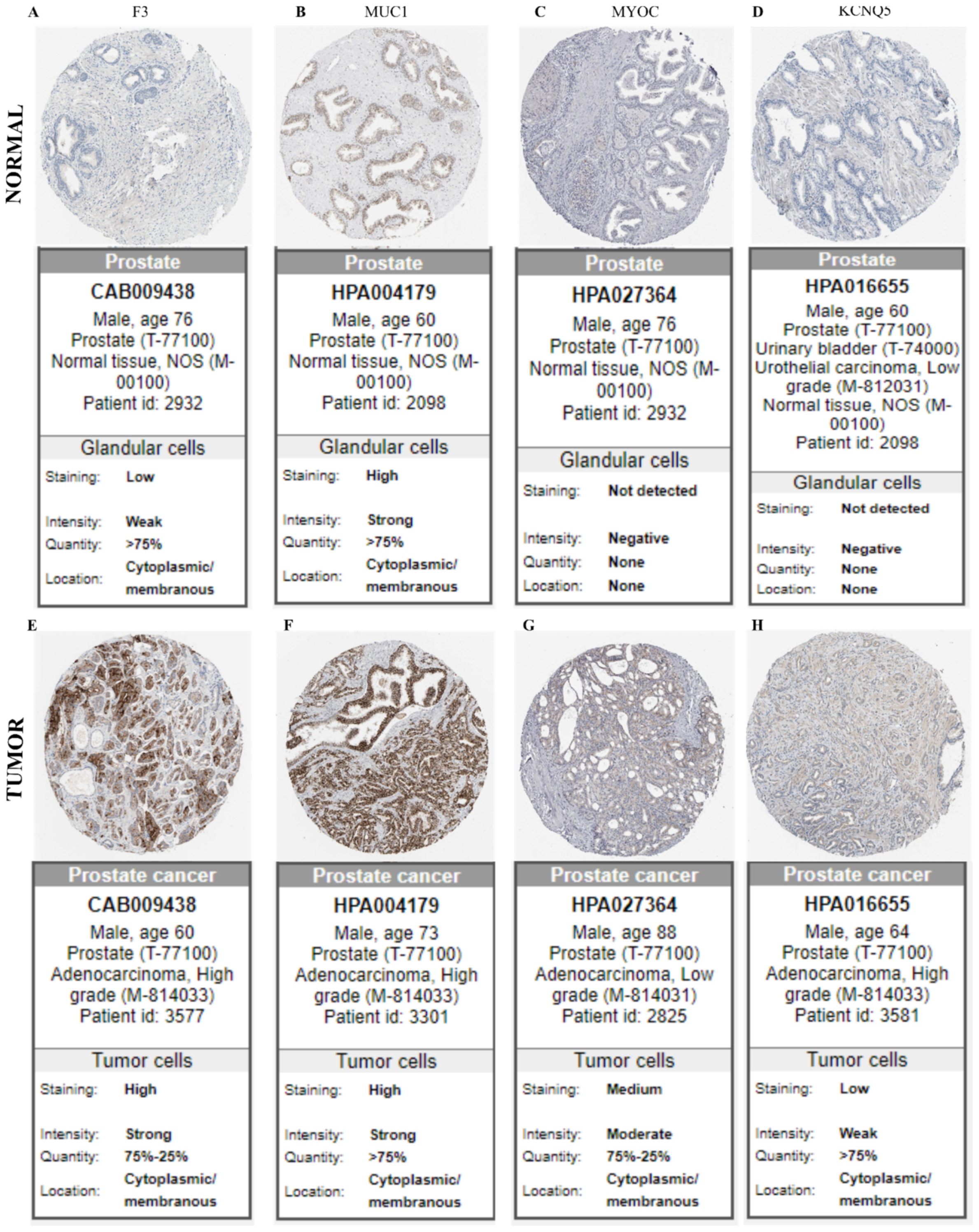
2.5. In Silico Validation of Protein Expression in Membrane and Secreted Proteins in Human Prostate Cancer
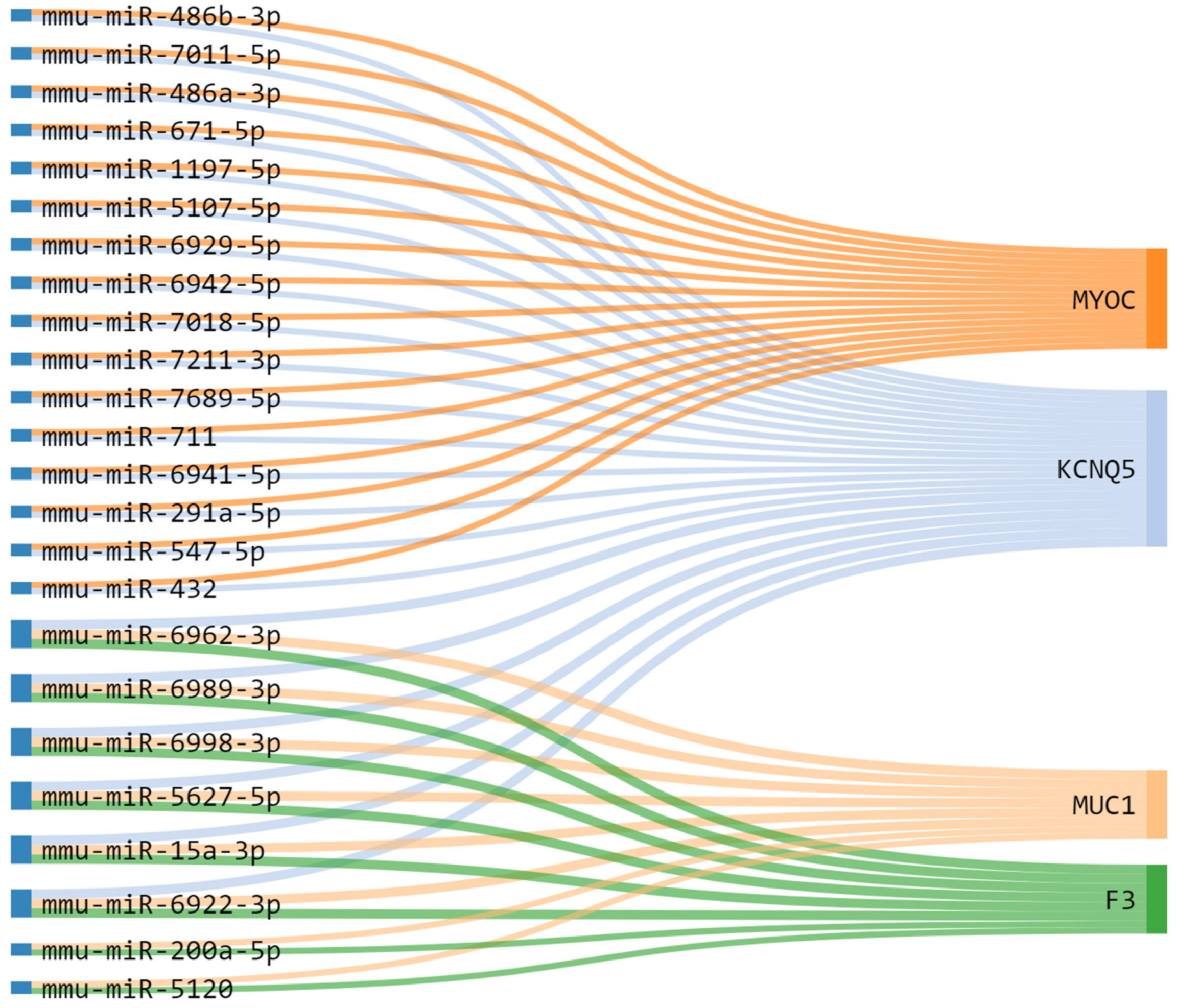
2.6. In Silico Prediction of miRNAs-mRNA Regulatory Modules in Prostate Cancer
3. Discussion
4. Materials and Methods
4.1. Analysis of RNA-Seq Data of the Genetically Engineered Mouse Model (GEMM) for PCa: The Pten Conditional Knockout
4.2. Integration of Secretome and Membrane Proteome Analyses to Identify Prostate Cancer Biomarkers
4.3. Protein–Protein Interaction Network and Functional Enrichment Analysis
4.4. Gene Expression Profile in Prostate Cancer
4.5. Survival Analysis and Risk Assessment
4.6. In Silico Validation of Differentially Expressed Genes (DEGs)
4.7. Prediction of Commonly Dysregulated miRNAs-mRNA Targets
4.8. Data Representation and Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef]
- Schatten, H. Brief Overview of Prostate Cancer Statistics, Grading, Diagnosis and Treatment Strategies. In Cell & Molecular Biology of Prostate Cancer; Springer: Cham, Switzerland, 2018; Volume 1095, pp. 1–14. [Google Scholar] [CrossRef]
- Litwin, M.S.; Tan, H.J. The Diagnosis and Treatment of Prostate Cancer: A Review. JAMA 2017, 317, 2532–2542. [Google Scholar] [CrossRef]
- Quigley, D.A.; Dang, H.X.; Zhao, S.G.; Lloyd, P.; Aggarwal, R.; Alumkal, J.J.; Foye, A.; Kothari, V.; Perry, M.D.; Bailey, A.M.; et al. Genomic Hallmarks and Structural Variation in Metastatic Prostate Cancer. Cell 2018, 174, 758–769.e9. [Google Scholar] [CrossRef]
- Hussain, M.; Mateo, J.; Fizazi, K.; Saad, F.; Shore, N.; Sandhu, S.; Chi, K.N.; Sartor, O.; Agarwal, N.; Olmos, D.; et al. Survival with Olaparib in Metastatic Castration-Resistant Prostate Cancer. N. Engl. J. Med. 2020, 383, 2345–2357. [Google Scholar] [CrossRef]
- Taylor, B.S.; Schultz, N.; Hieronymus, H.; Gopalan, A.; Xiao, Y.; Carver, B.S.; Arora, V.K.; Kaushik, P.; Cerami, E.; Reva, B.; et al. Integrative genomic profiling of human prostate cancer. Cancer Cell 2010, 18, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, A.D. PTEN-PI3K pathway alterations in advanced prostate cancer and clinical implications. Prostate 2022, 82 (Suppl. 1), S60–S72. [Google Scholar] [CrossRef]
- Pungsrinont, T.; Kallenbach, J.; Baniahmad, A. Role of PI3K-AKT-mTOR Pathway as a Pro-Survival Signaling and Resistance-Mediating Mechanism to Therapy of Prostate Cancer. Int. J. Mol. Sci. 2021, 22, 1088. [Google Scholar] [CrossRef]
- Qi, Z.; Xu, Z.; Zhang, L.; Zou, Y.; Li, J.; Yan, W.; Li, C.; Liu, N.; Wu, H. Overcoming resistance to immune checkpoint therapy in PTEN-null prostate cancer by intermittent anti-PI3Kα/β/δ treatment. Nat. Commun. 2022, 13, 182. [Google Scholar] [CrossRef]
- Jamaspishvili, T.; Berman, D.M.; Ross, A.E.; Scher, H.I.; De Marzo, A.M.; Squire, J.A.; Lotan, T.L. Clinical implications of PTEN loss in prostate cancer. Nat. Rev. Urol. 2018, 15, 222–234. [Google Scholar] [CrossRef] [PubMed]
- Turnham, D.J.; Bullock, N.; Dass, M.S.; Staffurth, J.N.; Pearson, H.B. The PTEN Conundrum: How to Target PTEN-Deficient Prostate Cancer. Cells 2020, 9, 2342. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Gao, J.; Lei, Q.; Rozengurt, N.; Pritchard, C.; Jiao, J.; Thomas, G.V.; Li, G.; Roy-Burman, P.; Nelson, P.S.; et al. Prostate-specific deletion of the murine Pten tumor suppressor gene leads to metastatic prostate cancer. Cancer Cell 2003, 4, 209–221. [Google Scholar] [CrossRef]
- Irshad, S.; Abate-Shen, C. Modeling prostate cancer in mice: Something old, something new, something premalignant, something metastatic. Cancer Metastasis Rev. 2013, 32, 109–122. [Google Scholar] [CrossRef]
- Chen, M.; Zhang, J.; Sampieri, K.; Clohessy, J.G.; Mendez, L.; Gonzalez-Billalabeitia, E.; Liu, X.S.; Lee, Y.R.; Fung, J.; Katon, J.M.; et al. An aberrant SREBP-dependent lipogenic program promotes metastatic prostate cancer. Nat. Genet. 2018, 50, 206–218. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, G.; Freire, P.P.; Cury, S.S.; de Moraes, D.; Oliveira, J.S.; Dal-Pai-Silva, M.; Reis, P.P.; Carvalho, R.F. An Integrated Meta-Analysis of Secretome and Proteome Identify Potential Biomarkers of Pancreatic Ductal Adenocarcinoma. Cancers 2020, 12, 716. [Google Scholar] [CrossRef]
- Latha, N.R.; Rajan, A.; Nadhan, R.; Achyutuni, S.; Sengodan, S.K.; Hemalatha, S.K.; Varghese, G.R.; Thankappan, R.; Krishnan, N.; Patra, D.; et al. Gene expression signatures: A tool for analysis of breast cancer prognosis and therapy. Crit. Rev. Oncol. Hematol. 2020, 151, 102964. [Google Scholar] [CrossRef]
- Liu, Y.; Wu, L.; Ao, H.; Zhao, M.; Leng, X.; Liu, M.; Ma, J.; Zhu, J. Prognostic implications of autophagy-associated gene signatures in non-small cell lung cancer. Aging 2019, 11, 11440–11462. [Google Scholar] [CrossRef]
- Tonella, L.; Giannoccaro, M.; Alfieri, S.; Canevari, S.; De Cecco, L. Gene Expression Signatures for Head and Neck Cancer Patient Stratification: Are Results Ready for Clinical Application? Curr. Treat. Options Oncol. 2017, 18, 32. [Google Scholar] [CrossRef]
- Venet, D.; Dumont, J.E.; Detours, V. Most random gene expression signatures are significantly associated with breast cancer outcome. PLoS Comput. Biol. 2011, 7, e1002240. [Google Scholar] [CrossRef]
- Hu, D.; Jiang, L.; Luo, S.; Zhao, X.; Hu, H.; Zhao, G.; Tang, W. Development of an autophagy-related gene expression signature for prognosis prediction in prostate cancer patients. J. Transl. Med. 2020, 18, 160. [Google Scholar] [CrossRef] [PubMed]
- Penney, K.L.; Sinnott, J.A.; Fall, K.; Pawitan, Y.; Hoshida, Y.; Kraft, P.; Stark, J.R.; Fiorentino, M.; Perner, S.; Finn, S.; et al. mRNA expression signature of Gleason grade predicts lethal prostate cancer. J. Clin. Oncol. 2011, 29, 2391–2396. [Google Scholar] [CrossRef] [PubMed]
- Shao, N.; Tang, H.; Mi, Y.; Zhu, Y.; Wan, F.; Ye, D. A novel gene signature to predict immune infiltration and outcome in patients with prostate cancer. Oncoimmunology 2020, 9, 1762473. [Google Scholar] [CrossRef] [PubMed]
- Ong, C.W.; Maxwell, P.; Alvi, M.A.; McQuaid, S.; Waugh, D.; Mills, I.; Salto-Tellez, M. A gene signature associated with PTEN activation defines good prognosis intermediate risk prostate cancer cases. J. Pathol. Clin. Res. 2018, 4, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Abida, W.; Cyrta, J.; Heller, G.; Prandi, D.; Armenia, J.; Coleman, I.; Cieslik, M.; Benelli, M.; Robinson, D.; Van Allen, E.M.; et al. Genomic correlates of clinical outcome in advanced prostate cancer. Proc. Natl. Acad. Sci. USA 2019, 116, 11428–11436. [Google Scholar] [CrossRef]
- Chibon, F. Cancer gene expression signatures—The rise and fall? Eur. J. Cancer 2013, 49, 2000–2009. [Google Scholar] [CrossRef]
- Cooper, C.S.; Campbell, C.; Jhavar, S. Mechanisms of Disease: Biomarkers and molecular targets from microarray gene expression studies in prostate cancer. Nat. Clin. Pract. Urol. 2007, 4, 677–687. [Google Scholar] [CrossRef]
- Michiels, S.; Koscielny, S.; Hill, C. Interpretation of microarray data in cancer. Br. J. Cancer 2007, 96, 1155–1158. [Google Scholar] [CrossRef]
- Cary, K.C.; Cooperberg, M.R. Biomarkers in prostate cancer surveillance and screening: Past, present, and future. Ther. Adv. Urol. 2013, 5, 318–329. [Google Scholar] [CrossRef]
- Saini, S. PSA and beyond: Alternative prostate cancer biomarkers. Cell Oncol. 2016, 39, 97–106. [Google Scholar] [CrossRef]
- Szeliski, K.; Adamowicz, J.; Gastecka, A.; Drewa, T.; Pokrywczyńska, M. Modern urology perspectives on prostate cancer biomarkers. Cent. Eur. J. Urol. 2018, 71, 420–426. [Google Scholar] [CrossRef]
- Denton, A.E.; Roberts, E.W.; Fearon, D.T. Stromal Cells in the Tumor Microenvironment. Adv. Exp. Med. Biol. 2018, 1060, 99–114. [Google Scholar] [CrossRef]
- Lalonde, E.; Ishkanian, A.S.; Sykes, J.; Fraser, M.; Ross-Adams, H.; Erho, N.; Dunning, M.J.; Halim, S.; Lamb, A.D.; Moon, N.C.; et al. Tumour genomic and microenvironmental heterogeneity for integrated prediction of 5-year biochemical recurrence of prostate cancer: A retrospective cohort study. Lancet Oncol. 2014, 15, 1521–1532. [Google Scholar] [CrossRef]
- Lunt, S.J.; Chaudary, N.; Hill, R.P. The tumor microenvironment and metastatic disease. Clin. Exp. Metastasis 2009, 26, 19–34. [Google Scholar] [CrossRef] [PubMed]
- Berglund, E.; Maaskola, J.; Schultz, N.; Friedrich, S.; Marklund, M.; Bergenstråhle, J.; Tarish, F.; Tanoglidi, A.; Vickovic, S.; Larsson, L.; et al. Spatial maps of prostate cancer transcriptomes reveal an unexplored landscape of heterogeneity. Nat. Commun. 2018, 9, 2419. [Google Scholar] [CrossRef]
- Bacac, M.; Provero, P.; Mayran, N.; Stehle, J.C.; Fusco, C.; Stamenkovic, I. A mouse stromal response to tumor invasion predicts prostate and breast cancer patient survival. PLoS ONE 2006, 1, e32. [Google Scholar] [CrossRef]
- Planche, A.; Bacac, M.; Provero, P.; Fusco, C.; Delorenzi, M.; Stehle, J.C.; Stamenkovic, I. Identification of prognostic molecular features in the reactive stroma of human breast and prostate cancer. PLoS ONE 2011, 6, e18640. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Morris, J.H.; Cook, H.; Kuhn, M.; Wyder, S.; Simonovic, M.; Santos, A.; Doncheva, N.T.; Roth, A.; Bork, P.; et al. The STRING database in 2017: Quality-controlled protein-protein association networks, made broadly accessible. Nucleic Acids Res. 2017, 45, D362–D368. [Google Scholar] [CrossRef]
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Zhang, Z. GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017, 45, W98–W102. [Google Scholar] [CrossRef]
- Dunning, M.J.; Vowler, S.L.; Lalonde, E.; Ross-Adams, H.; Boutros, P.; Mills, I.G.; Lynch, A.G.; Lamb, A.D. Mining Human Prostate Cancer Datasets: The “camcAPP” Shiny App. EBioMedicine 2017, 17, 5–6. [Google Scholar] [CrossRef]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci. Signal. 2013, 6, pl1. [Google Scholar] [CrossRef] [PubMed]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [PubMed]
- Ross-Adams, H.; Lamb, A.D.; Dunning, M.J.; Halim, S.; Lindberg, J.; Massie, C.M.; Egevad, L.A.; Russell, R.; Ramos-Montoya, A.; Vowler, S.L.; et al. Integration of copy number and transcriptomics provides risk stratification in prostate cancer: A discovery and validation cohort study. EBioMedicine 2015, 2, 1133–1144. [Google Scholar] [CrossRef]
- Grasso, C.S.; Wu, Y.M.; Robinson, D.R.; Cao, X.; Dhanasekaran, S.M.; Khan, A.P.; Quist, M.J.; Jing, X.; Lonigro, R.J.; Brenner, J.C.; et al. The mutational landscape of lethal castration-resistant prostate cancer. Nature 2012, 487, 239–243. [Google Scholar] [CrossRef]
- Haffner, M.C.; Zwart, W.; Roudier, M.P.; True, L.D.; Nelson, W.G.; Epstein, J.I.; De Marzo, A.M.; Nelson, P.S.; Yegnasubramanian, S. Genomic and phenotypic heterogeneity in prostate cancer. Nat. Rev. Urol. 2021, 18, 79–92. [Google Scholar] [CrossRef]
- Labrecque, M.P.; Coleman, I.M.; Brown, L.G.; True, L.D.; Kollath, L.; Lakely, B.; Nguyen, H.M.; Yang, Y.C.; da Costa, R.M.G.; Kaipainen, A.; et al. Molecular profiling stratifies diverse phenotypes of treatment-refractory metastatic castration-resistant prostate cancer. J. Clin. Investig. 2019, 129, 4492–4505. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Guo, J.; Liu, K.; Chen, L.; Liu, D.; Dong, S.; Xia, J.; Long, Q.; Yue, Y.; Zhao, P.; et al. Identification of a distinct luminal subgroup diagnosing and stratifying early stage prostate cancer by tissue-based single-cell RNA sequencing. Mol. Cancer 2020, 19, 147. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Dai, Y. Tumor microenvironment and therapeutic response. Cancer Lett. 2017, 387, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Ippolito, L.; Morandi, A.; Taddei, M.L.; Parri, M.; Comito, G.; Iscaro, A.; Raspollini, M.R.; Magherini, F.; Rapizzi, E.; Masquelier, J.; et al. Cancer-associated fibroblasts promote prostate cancer malignancy via metabolic rewiring and mitochondrial transfer. Oncogene 2019, 38, 5339–5355. [Google Scholar] [CrossRef]
- Sun, D.Y.; Wu, J.Q.; He, Z.H.; He, M.F.; Sun, H.B. Cancer-associated fibroblast regulate proliferation and migration of prostate cancer cells through TGF-β signaling pathway. Life Sci. 2019, 235, 116791. [Google Scholar] [CrossRef]
- Archer, M.; Dogra, N.; Kyprianou, N. Inflammation as a Driver of Prostate Cancer Metastasis and Therapeutic Resistance. Cancers 2020, 12, 2984. [Google Scholar] [CrossRef] [PubMed]
- Larionova, I.; Tuguzbaeva, G.; Ponomaryova, A.; Stakheyeva, M.; Cherdyntseva, N.; Pavlov, V.; Choinzonov, E.; Kzhyshkowska, J. Tumor-Associated Macrophages in Human Breast, Colorectal, Lung, Ovarian and Prostate Cancers. Front. Oncol. 2020, 10, 566511. [Google Scholar] [CrossRef]
- Andersen, M.K.; Rise, K.; Giskeødegård, G.F.; Richardsen, E.; Bertilsson, H.; Størkersen, Ø.; Bathen, T.F.; Rye, M.; Tessem, M.B. Integrative metabolic and transcriptomic profiling of prostate cancer tissue containing reactive stroma. Sci. Rep. 2018, 8, 14269. [Google Scholar] [CrossRef] [PubMed]
- Jia, Z.; Rahmatpanah, F.B.; Chen, X.; Lernhardt, W.; Wang, Y.; Xia, X.Q.; Sawyers, A.; Sutton, M.; McClelland, M.; Mercola, D. Expression changes in the stroma of prostate cancer predict subsequent relapse. PLoS ONE 2012, 7, e41371. [Google Scholar] [CrossRef]
- Karkampouna, S.; De Filippo, M.R.; Ng, C.K.Y.; Klima, I.; Zoni, E.; Spahn, M.; Stein, F.; Haberkant, P.; Thalmann, G.N.; Kruithof-de Julio, M. Stroma Transcriptomic and Proteomic Profile of Prostate Cancer Metastasis Xenograft Models Reveals Prognostic Value of Stroma Signatures. Cancers 2020, 12, 3786. [Google Scholar] [CrossRef]
- Mo, F.; Lin, D.; Takhar, M.; Ramnarine, V.R.; Dong, X.; Bell, R.H.; Volik, S.V.; Wang, K.; Xue, H.; Wang, Y.; et al. Stromal Gene Expression is Predictive for Metastatic Primary Prostate Cancer. Eur. Urol. 2018, 73, 524–532. [Google Scholar] [CrossRef]
- Santos, N.J.; Ximenes, P.P.; Constantino, F.B.; Carvalho, H.F.; Felisbino, S.L. Mucinous metaplasia in Pten conditional knockout mice and mucin family genes as prognostic markers for prostate cancer. Life Sci. 2022, 293, 120264. [Google Scholar] [CrossRef] [PubMed]
- Santos, N.J.; Barquilha, C.N.; Barbosa, I.C.; Macedo, R.T.; Lima, F.O.; Justulin, L.A.; Barbosa, G.O.; Carvalho, H.F.; Felisbino, S.L. Syndecan Family Gene and Protein Expression and Their Prognostic Values for Prostate Cancer. Int. J. Mol. Sci. 2021, 22, 8669. [Google Scholar] [CrossRef]
- Jurmeister, S.; Ramos-Montoya, A.; Sandi, C.; Pertega-Gomes, N.; Wadhwa, K.; Lamb, A.D.; Dunning, M.J.; Attig, J.; Carroll, J.S.; Fryer, L.G.; et al. Identification of potential therapeutic targets in prostate cancer through a cross-species approach. EMBO Mol. Med. 2018, 10, e8274. [Google Scholar] [CrossRef]
- Kaliman, P.; Llagostera, E. Myotonic dystrophy protein kinase (DMPK) and its role in the pathogenesis of myotonic dystrophy 1. Cell. Signal. 2008, 20, 1935–1941. [Google Scholar] [CrossRef]
- Itoh, K.; Ebata, T.; Hirata, H.; Torii, T.; Sugimoto, W.; Onodera, K.; Nakajima, W.; Uehara, I.; Okuzaki, D.; Yamauchi, S.; et al. DMPK is a New Candidate Mediator of Tumor Suppressor p53-Dependent Cell Death. Molecules 2019, 24, 3175. [Google Scholar] [CrossRef] [PubMed]
- MacLennan, D.H.; Kranias, E.G. Phospholamban: A crucial regulator of cardiac contractility. Nat. Rev. Mol. Cell Biol. 2003, 4, 566–577. [Google Scholar] [CrossRef] [PubMed]
- Gautier, M.; Trebak, M.; Fleig, A.; Vandier, C.; Ouadid-Ahidouch, H. Ca2+ homeostasis and cancer. Cell Calcium 2019, 84, 102084. [Google Scholar] [CrossRef] [PubMed]
- Kania, E.; Pająk, B.; Orzechowski, A. Calcium homeostasis and ER stress in control of autophagy in cancer cells. Biomed. Res. Int. 2015, 2015, 352794. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Zhao, G.; Yuan, M.; Liu, X.; Ma, Y.; Miao, B.; Zhao, S.; Li, D.; Xiong, S.; Zheng, M.; et al. KCNQ5 and C9orf50 Methylation in Stool DNA for Early Detection of Colorectal Cancer. Front. Oncol. 2020, 10, 621295. [Google Scholar] [CrossRef]
- Jensen, S.; Øgaard, N.; Ørntoft, M.W.; Rasmussen, M.H.; Bramsen, J.B.; Kristensen, H.; Mouritzen, P.; Madsen, M.R.; Madsen, A.H.; Sunesen, K.G.; et al. Novel DNA methylation biomarkers show high sensitivity and specificity for blood-based detection of colorectal cancer—A clinical biomarker discovery and validation study. Clin. Epigenetics 2019, 11, 158. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wu, Q.; Xu, L.; Wang, H.; Liu, X.; Li, S.; Hu, T.; Liu, Y.; Peng, Q.; Chen, Z.; et al. Sensitive detection of colorectal cancer in peripheral blood by a novel methylation assay. Clin. Epigenetics 2021, 13, 90. [Google Scholar] [CrossRef]
- Bouzid, A.; Smeti, I.; Dhouib, L.; Roche, M.; Achour, I.; Khalfallah, A.; Gibriel, A.A.; Charfeddine, I.; Ayadi, H.; Lachuer, J.; et al. Down-expression of P2RX2, KCNQ5, ERBB3 and SOCS3 through DNA hypermethylation in elderly women with presbycusis. Biomarkers 2018, 23, 347–356. [Google Scholar] [CrossRef]
- Zimmer, J.; Takahashi, T.; Hofmann, A.D.; Puri, P. Downregulation of KCNQ5 expression in the rat pulmonary vasculature of nitrofen-induced congenital diaphragmatic hernia. J. Pediatr. Surg. 2017, 52, 702–705. [Google Scholar] [CrossRef]
- Goldwich, A.; Scholz, M.; Tamm, E.R. Myocilin promotes substrate adhesion, spreading and formation of focal contacts in podocytes and mesangial cells. Histochem. Cell Biol. 2009, 131, 167–180. [Google Scholar] [CrossRef]
- Liuska, P.J.; Harju, M.; Kivelä, T.T.; Turunen, J.A. Prevalence of MYOC risk variants for glaucoma in different populations. Acta Ophthalmol. 2021, 99, e1090–e1097. [Google Scholar] [CrossRef]
- Wiggs, J.L.; Pasquale, L.R. Genetics of glaucoma. Hum. Mol. Genet. 2017, 26, R21–R27. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Ke, J.; Du, X.; Yu, Z.; Gao, D. Genetic characterization of thymoma. Sci. Rep. 2019, 9, 2369. [Google Scholar] [CrossRef] [PubMed]
- Gajos-Michniewicz, A.; Czyz, M. WNT Signaling in Melanoma. Int. J. Mol. Sci. 2020, 21, 4852. [Google Scholar] [CrossRef]
- Nusse, R.; Clevers, H. Wnt/β-Catenin Signaling, Disease, and Emerging Therapeutic Modalities. Cell 2017, 169, 985–999. [Google Scholar] [CrossRef]
- Galluzzi, L.; Spranger, S.; Fuchs, E.; López-Soto, A. WNT Signaling in Cancer Immunosurveillance. Trends Cell Biol. 2019, 29, 44–65. [Google Scholar] [CrossRef] [PubMed]
- Ring, A.; Kim, Y.M.; Kahn, M. Wnt/catenin signaling in adult stem cell physiology and disease. Stem. Cell Rev. Rep. 2014, 10, 512–525. [Google Scholar] [CrossRef]
- Kerekes, K.; Bányai, L.; Trexler, M.; Patthy, L. Structure, function and disease relevance of Wnt inhibitory factor 1, a secreted protein controlling the Wnt and hedgehog pathways. Growth Factors 2019, 37, 29–52. [Google Scholar] [CrossRef]
- Beggs, A.D.; Mehta, S.; Deeks, J.J.; James, J.D.; Caldwell, G.M.; Dilworth, M.P.; Stockton, J.D.; Blakeway, D.; Pestinger, V.; Vince, A.; et al. Validation of epigenetic markers to identify colitis associated cancer: Results of module 1 of the ENDCAP-C study. EBioMedicine 2019, 39, 265–271. [Google Scholar] [CrossRef]
- Hu, H.; Li, B.; Zhou, C.; Ying, X.; Chen, M.; Huang, T.; Chen, Y.; Ji, H.; Pan, R.; Wang, T.; et al. Diagnostic value of WIF1 methylation for colorectal cancer: A meta-analysis. Oncotarget 2018, 9, 5378–5386. [Google Scholar] [CrossRef]
- Liu, S.; Chen, X.; Chen, R.; Wang, J.; Zhu, G.; Jiang, J.; Wang, H.; Duan, S.; Huang, J. Diagnostic role of Wnt pathway gene promoter methylation in non small cell lung cancer. Oncotarget 2017, 8, 36354–36367. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Fu, J.; Bi, H.; Ge, A.; Xia, T.; Liu, Y.; Sun, H.; Li, D.; Zhao, Y. DNA methylation of SFRP1, SFRP2, and WIF1 and prognosis of postoperative colorectal cancer patients. BMC Cancer 2019, 19, 1212. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Zhong, X.; Gao, J.; Song, R.; Wu, H.; Zi, S.; Yang, S.; Du, P.; Cui, L.; Yang, C.; et al. Coexpression of SFRP1 and WIF1 as a prognostic predictor of favorable outcomes in patients with colorectal carcinoma. Biomed. Res. Int. 2014, 2014, 256723. [Google Scholar] [CrossRef] [PubMed]
- Cortez, M.A.; Masrorpour, F.; Ivan, C.; Zhang, J.; Younes, A.I.; Lu, Y.; Estecio, M.R.; Barsoumian, H.B.; Menon, H.; Caetano, M.D.S.; et al. Bone morphogenetic protein 7 promotes resistance to immunotherapy. Nat. Commun. 2020, 11, 4840. [Google Scholar] [CrossRef]
- Perron, J.C.; Rodrigues, A.A.; Surubholta, N.; Dodd, J. Chemotropic signaling by BMP7 requires selective interaction at a key residue in ActRIIA. Biol. Open 2019, 8, bio042283. [Google Scholar] [CrossRef]
- Aoki, M.; Ishigami, S.; Uenosono, Y.; Arigami, T.; Uchikado, Y.; Kita, Y.; Kurahara, H.; Matsumoto, M.; Ueno, S.; Natsugoe, S. Expression of BMP-7 in human gastric cancer and its clinical significance. Br. J. Cancer 2011, 104, 714–718. [Google Scholar] [CrossRef]
- Chen, J.; Ye, L.; Xie, F.; Yang, Y.; Zhang, L.; Jiang, W.G. Expression of bone morphogenetic protein 7 in lung cancer and its biological impact on lung cancer cells. Anticancer Res. 2010, 30, 1113–1120. [Google Scholar]
- Witkowski, M.; Landmesser, U.; Rauch, U. Tissue factor as a link between inflammation and coagulation. Trends Cardiovasc. Med. 2016, 26, 297–303. [Google Scholar] [CrossRef]
- Hisada, Y.; Mackman, N. Tissue Factor and Cancer: Regulation, Tumor Growth, and Metastasis. Semin. Thromb. Hemost. 2019, 45, 385–395. [Google Scholar] [CrossRef]
- Khorana, A.A.; Ahrendt, S.A.; Ryan, C.K.; Francis, C.W.; Hruban, R.H.; Hu, Y.C.; Hostetter, G.; Harvey, J.; Taubman, M.B. Tissue factor expression, angiogenesis, and thrombosis in pancreatic cancer. Clin. Cancer Res. 2007, 13, 2870–2875. [Google Scholar] [CrossRef]
- Kobayashi, S.; Koizume, S.; Takahashi, T.; Ueno, M.; Oishi, R.; Nagashima, S.; Sano, Y.; Fukushima, T.; Tezuka, S.; Morimoto, M.; et al. Tissue factor and its procoagulant activity on cancer-associated thromboembolism in pancreatic cancer. Cancer Sci. 2021, 112, 4679–4691. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Cheng, C.; Gou, J.; Yi, T.; Qian, Y.; Du, X. Expression of tissue factor in human cervical carcinoma tissue. Exp. Ther. Med. 2018, 16, 4075–4081. [Google Scholar] [CrossRef] [PubMed]
- Shaker, H.; Bundred, N.J.; Landberg, G.; Pritchard, S.A.; Albadry, H.; Nicholson, S.L.; Harries, L.J.; Heah, J.Y.E.; Castle, J.; Kirwan, C.C. Breast cancer stromal clotting activation (Tissue Factor and thrombin): A pre-invasive phenomena that is prognostic in invasion. Cancer Med. 2020, 9, 1768–1778. [Google Scholar] [CrossRef] [PubMed]
- Hell, L.; Däullary, T.; Burghart, V.; Mauracher, L.M.; Grilz, E.; Moser, B.; Kramer, G.; Schmid, J.A.; Ay, C.; Pabinger, I.; et al. Extracellular Vesicle-Associated Tissue Factor Activity in Prostate Cancer Patients with Disseminated Intravascular Coagulation. Cancers 2021, 13, 1487. [Google Scholar] [CrossRef]
- Chen, W.; Zhang, Z.; Zhang, S.; Zhu, P.; Ko, J.K.; Yung, K.K. MUC1: Structure, Function, and Clinic Application in Epithelial Cancers. Int. J. Mol. Sci. 2021, 22, 6567. [Google Scholar] [CrossRef] [PubMed]
- Nath, S.; Mukherjee, P. MUC1: A multifaceted oncoprotein with a key role in cancer progression. Trends Mol. Med. 2014, 20, 332–342. [Google Scholar] [CrossRef] [PubMed]
- Bouillez, A.; Rajabi, H.; Pitroda, S.; Jin, C.; Alam, M.; Kharbanda, A.; Tagde, A.; Wong, K.K.; Kufe, D. Inhibition of MUC1-C Suppresses MYC Expression and Attenuates Malignant Growth in KRAS Mutant Lung Adenocarcinomas. Cancer Res. 2016, 76, 1538–1548. [Google Scholar] [CrossRef]
- Hagiwara, M.; Yasumizu, Y.; Yamashita, N.; Rajabi, H.; Fushimi, A.; Long, M.D.; Li, W.; Bhattacharya, A.; Ahmad, R.; Oya, M.; et al. MUC1-C Activates the BAF (mSWI/SNF) Complex in Prostate Cancer Stem Cells. Cancer Res. 2021, 81, 1111–1122. [Google Scholar] [CrossRef]
- Yasumizu, Y.; Rajabi, H.; Jin, C.; Hata, T.; Pitroda, S.; Long, M.D.; Hagiwara, M.; Li, W.; Hu, Q.; Liu, S.; et al. MUC1-C regulates lineage plasticity driving progression to neuroendocrine prostate cancer. Nat. Commun. 2020, 11, 338. [Google Scholar] [CrossRef]
- Acunzo, M.; Romano, G.; Wernicke, D.; Croce, C.M. MicroRNA and cancer—A brief overview. Adv. Biol. Regul. 2015, 57, 1–9. [Google Scholar] [CrossRef]
- Rupaimoole, R.; Slack, F.J. MicroRNA therapeutics: Towards a new era for the management of cancer and other diseases. Nat. Rev. Drug Discov. 2017, 16, 203–222. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Jing, L.; Han, J.; Wang, G.; Liu, Y.; Zhang, X.; Wang, Y.; Wang, F.; Ma, H. MicroRNA 486-3p directly targets BIK and regulates apoptosis and invasion in colorectal cancer cells. Onco Targets Ther. 2018, 11, 8791–8801. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Ai, J.; Long, H.; Liu, W.; Wang, X.; Zuo, Y.; Li, Y.; Wu, Q.; Deng, Y. Integrative microRNA and gene profiling data analysis reveals novel biomarkers and mechanisms for lung cancer. Oncotarget 2016, 7, 8441–8454. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Ye, Z. Identification of Serum miR-337-3p, miR-484, miR-582, and miR-3677 as Promising Biomarkers for Osteosarcoma. Clin. Lab. 2021, 67. [Google Scholar] [CrossRef]
- Fan, B.; Chen, L.P.; Yuan, Y.H.; Xiao, H.N.; Lv, X.S.; Xia, Z.Y. MiR-15a-3p suppresses the growth and metastasis of ovarian cancer cell by targeting Twist1. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 1934–1946. [Google Scholar] [CrossRef]
- Furuya, T.K.; Murta, C.B.; Murillo Carrasco, A.G.; Uno, M.; Sichero, L.; Villa, L.L.; Cardilli, L.; Coelho, R.F.; Guglielmetti, G.B.; Cordeiro, M.D.; et al. Disruption of miRNA-mRNA Networks Defines Novel Molecular Signatures for Penile Carcinogenesis. Cancers 2021, 13, 4745. [Google Scholar] [CrossRef]
- Liu, S.J.; Wang, W.T.; Zhang, F.L.; Yu, Y.H.; Yu, H.J.; Liang, Y.; Li, N.; Li, Y.B. miR-15a-3p affects the proliferation, migration and apoptosis of lens epithelial cells. Mol. Med. Rep. 2019, 19, 1110–1116. [Google Scholar] [CrossRef]
- Cui, Y.; Yang, Y.; Ren, L.; Yang, J.; Wang, B.; Xing, T.; Chen, H.; Chen, M. miR-15a-3p Suppresses Prostate Cancer Cell Proliferation and Invasion by Targeting SLC39A7 via Downregulating Wnt/β-Catenin Signaling Pathway. Cancer Biother. Radiopharm. 2019, 34, 472–479. [Google Scholar] [CrossRef]
- Xin, C.; Lu, S.; Li, Y.; Zhang, Y.; Tian, J.; Zhang, S.; Yang, S.; Gao, T.; Xu, J. miR-671-5p Inhibits Tumor Proliferation by Blocking Cell Cycle in Osteosarcoma. DNA Cell Biol. 2019, 38, 996–1004. [Google Scholar] [CrossRef]
- Hu, Y.; Liang, D.; Chen, X.; Chen, L.; Bai, J.; Li, H.; Yin, C.; Zhong, W. MiR-671-5p negatively regulates SMAD3 to inhibit migration and invasion of osteosarcoma cells. Nan Fang Yi Ke Da Xue Xue Bao 2021, 41, 1562–1568. [Google Scholar] [CrossRef]
- Barquilha, C.N.; Santos, N.J.; Monção, C.C.D.; Barbosa, I.C.; Lima, F.O.; Justulin, L.A.; Pértega-Gomes, N.; Felisbino, S.L. Sulfiredoxin as a Potential Therapeutic Target for Advanced and Metastatic Prostate Cancer. Oxidative Med. Cell. Longev. 2020, 2020, 2148562. [Google Scholar] [CrossRef] [PubMed]
- Thul, P.J.; Åkesson, L.; Wiking, M.; Mahdessian, D.; Geladaki, A.; Ait Blal, H.; Alm, T.; Asplund, A.; Björk, L.; Breckels, L.M.; et al. A subcellular map of the human proteome. Science 2017, 356, eaal3321. [Google Scholar] [CrossRef] [PubMed]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Proteomics. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Fagerberg, L.; Jonasson, K.; von Heijne, G.; Uhlén, M.; Berglund, L. Prediction of the human membrane proteome. Proteomics 2010, 10, 1141–1149. [Google Scholar] [CrossRef] [PubMed]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef] [PubMed]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef]
- Ge, S.X.; Jung, D.; Yao, R. ShinyGO: A graphical gene-set enrichment tool for animals and plants. Bioinformatics 2020, 36, 2628–2629. [Google Scholar] [CrossRef]
- Sticht, C.; De La Torre, C.; Parveen, A.; Gretz, N. miRWalk: An online resource for prediction of microRNA binding sites. PLoS ONE 2018, 13, e0206239. [Google Scholar] [CrossRef]
- Oliveros, J.C. An Interactive Tool for Comparing Lists with Venn Diagrams. 2007. Available online: https://bioinfogp.cnb.csic.es/tools/venny/index.html (accessed on 21 December 2021).
- Hothorn, T.; Hornik, K.; Zeileis, A. Unbiased Recursive Partitioning: A Conditional Inference Framework. J. Comput. Graph. Stat. 2006, 15, 651–674. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| mPIN Upregulated Gene Ontology (GO) | ||||
| Pathway ID | Pathway Description | Gene Count | Adjusted p-Value | Log10 |
| GO:0005887 | Integral component of plasma membrane | 32 | 1.73 × 10−11 | 10.76299509 |
| GO:0004713 | Protein tyrosine kinase activity | 5 | 0.012138352 | 1.915840284 |
| GO:0016758 | Hexosyltransferase activity | 5 | 0.014297294 | 1.844746144 |
| GO:0043190 | ATP-binding cassette (ABC) transporter complex | 2 | 0.019180885 | 1.717131357 |
| GO:0008194 | UDP-glycosyltransferase activity | 4 | 0.024662147 | 1.607969117 |
| GO:0009312 | Oligosaccharide biosynthetic process | 3 | 0.034162979 | 1.466444268 |
| GO:0016051 | Carbohydrate biosynthetic process | 3 | 0.034162979 | 1.466444268 |
| GO:0019375 | Galactolipid biosynthetic process | 2 | 0.034162979 | 1.466444268 |
| GO:0006682 | Galactosylceramide biosynthetic process | 2 | 0.034162979 | 1.466444268 |
| GO:0046476 | Glycosylceramide biosynthetic process | 2 | 0.0382352 | 1.417536631 |
| GO:0006681 | Galactosylceramide metabolic process | 2 | 0.041917255 | 1.377607169 |
| GO:0030165 | PDZ domain binding | 3 | 0.048747995 | 1.31204324 |
| MT Upregulated Gene Ontology (GO) | ||||
| Pathway ID | Pathway Description | Gene Count | Adjusted p-Value | Log10 |
| GO:0005887 | Integral component of plasma membrane | 43 | 9.77 × 10−17 | 16.01024386 |
| GO:0016323 | Basolateral plasma membrane | 10 | 0.00000211 | 5.675736848 |
| GO:0004713 | Protein tyrosine kinase activity | 7 | 0.001099549 | 2.9587853 |
| GO:0019199 | Transmembrane receptor protein kinase activity | 5 | 0.002610742 | 2.5832361 |
| GO:0004714 | Transmembrane receptor protein tyrosine kinase activity | 5 | 0.002610742 | 2.5832361 |
| GO:0005254 | Chloride channel activity | 5 | 0.00268003 | 2.571860416 |
| GO:0045121 | Membrane raft | 7 | 0.002867281 | 2.542529695 |
| GO:0043005 | Neuron projection | 11 | 0.01394385 | 1.855617311 |
| GO:0007167 | Enzyme linked receptor protein signaling pathway | 7 | 0.024380271 | 1.612961464 |
| AT Upregulated Gene Ontology (GO) | ||||
| Pathway ID | Pathway Description | Gene Count | Adjusted p-Value | Log10 |
| GO:0005887 | Integral component of plasma membrane | 46 | 1.74 × 10−17 | 16.75829697 |
| GO:0030667 | Secretory granule membrane | 13 | 0.00000273 | 5.564593362 |
| GO:0006954 | Inflammatory response | 11 | 0.000308 | 3.511189697 |
| GO:0071345 | Cellular response to cytokine stimulus | 15 | 0.000421 | 3.375939369 |
| GO:0002283 | Neutrophil activation involved in immune response | 13 | 0.00509802 | 2.292598474 |
| GO:0004713 | Protein tyrosine kinase activity | 6 | 0.00518138 | 2.285554596 |
| GO:0070372 | Regulation of ERK1 and ERK2 cascade | 9 | 0.005657676 | 2.247361946 |
| GO:0032103 | Positive regulation of response to external stimulus | 7 | 0.006311353 | 2.199877562 |
| GO:0043410 | Positive regulation of MAPK cascade | 9 | 0.008909311 | 2.050155885 |
| GO:0030198 | Extracellular matrix organization | 9 | 0.012342256 | 1.908605459 |
| GO:0005925 | Focal adhesion | 9 | 0.013649783 | 1.864874257 |
| GO:0005789 | Endoplasmic reticulum membrane | 12 | 0.028337729 | 1.547634959 |
| mPIN Upregulated Gene Ontology (GO) | ||||
| Pathway ID | Pathway Description | Gene Count | Adjusted p-Value | Log10 |
| GO:0019221 | Cytokine-mediated signaling pathway | 10 | 0.0000154 | 4.813747101 |
| GO:0062023 | Collagen-containing extracellular matrix | 7 | 0.0000354 | 4.451038902 |
| GO:0016019 | Peptidoglycan immune receptor activity | 2 | 0.000428 | 3.368891105 |
| GO:1902533 | Positive regulation of intracellular signal transduction | 7 | 0.001214858 | 2.915474599 |
| GO:0042834 | Peptidoglycan binding | 2 | 0.002962979 | 2.528271463 |
| GO:0008284 | Positive regulation of cell population proliferation | 6 | 0.003344125 | 2.475717449 |
| GO:0001774 | Microglial cell activation | 2 | 0.008863214 | 2.052408777 |
| GO:0030855 | Epithelial cell differentiation | 3 | 0.008882339 | 2.051472641 |
| GO:0033628 | Regulation of cell adhesion mediated by integrin | 2 | 0.01496553 | 1.824907884 |
| GO:0030198 | Extracellular matrix organization | 4 | 0.01496553 | 1.824907884 |
| GO:0010811 | Positive regulation of cell-substrate adhesion | 2 | 0.037254018 | 1.428826881 |
| GO:0060252 | Positive regulation of glial cell proliferation | 1 | 0.04201877 | 1.376556662 |
| GO:0033631 | Cell-cell adhesion mediated by integrin | 1 | 0.04201877 | 1.376556662 |
| GO:0071902 | positive regulation of protein serine/threonine kinase activity | 2 | 0.046363298 | 1.333825674 |
| MT Upregulated Gene Ontology (GO) | ||||
| Pathway ID | Pathway Description | Gene Count | Adjusted p-Value | Log10 |
| GO:0034774 | Secretory granule lumen | 14 | 4.44 × 10−12 | 11.35278878 |
| GO:0006954 | Inflammatory response | 13 | 2.06 × 10−11 | 10.6853178 |
| GO:0062023 | Collagen-containing extracellular matrix | 12 | 9.21 × 10−9 | 8.035890283 |
| GO:1905517 | Macrophage migration | 3 | 0.000185 | 3.732564754 |
| GO:0007160 | Cell-matrix adhesion | 3 | 0.011210456 | 1.950376724 |
| GO:0030855 | Epithelial cell differentiation | 3 | 0.011480631 | 1.940034241 |
| GO:0098632 | Cell-cell adhesion mediator activity | 2 | 0.025492739 | 1.5935835 |
| GO:0030203 | Glycosaminoglycan metabolic process | 2 | 0.031920127 | 1.495935395 |
| GO:0045545 | Syndecan binding | 1 | 0.045268101 | 1.344207727 |
| GO:0048708 | Astrocyte differentiation | 1 | 0.048279814 | 1.316234416 |
| GO:0008347 | Glial cell migration | 1 | 0.048279814 | 1.316234416 |
| AT Upregulated Gene Ontology (GO) | ||||
| Pathway ID | Pathway Description | Gene Count | Adjusted p-Value | Log10 |
| GO:0048018 | Receptor ligand activity | 19 | 3.70 × 10−18 | 17.43149267 |
| GO:0019221 | Cytokine-mediated signaling pathway | 23 | 5.23 × 10−16 | 15.28185137 |
| GO:0006954 | Inflammatory response | 15 | 1.64 × 10−13 | 12.78451214 |
| GO:0062023 | Collagen-containing extracellular matrix | 13 | 1.23 × 10−8 | 7.90940544 |
| GO:0008284 | Positive regulation of cell population proliferation | 13 | 2.40 × 10−7 | 6.620158329 |
| GO:0070374 | Positive regulation of ERK1 and ERK2 cascade | 9 | 3.14 × 10−7 | 6.503756602 |
| GO:0030198 | Extracellular matrix organization | 7 | 0.0000857 | 4.066941739 |
| GO:0000165 | MAPK cascade | 7 | 0.0000906 | 4.043108741 |
| GO:1905517 | Macrophage migration | 2 | 0.000194 | 3.712020847 |
| GO:0043062 | Extracellular structure organization | 5 | 0.000676 | 3.170015558 |
| GO:0042834 | Peptidoglycan binding | 1 | 0.006637813 | 2.177975013 |
| mPIN Downregulated Gene Ontology (GO) | ||||
| Pathway ID | Pathway Description | Gene Count | Adjusted p-Value | Log10 |
| GO:0098662 | Inorganic cation transmembrane transport | 11 | 2.10 × 10−8 | 7.677498224 |
| GO:0005887 | Integral component of plasma membrane | 19 | 2.38 × 10−8 | 7.623546608 |
| GO:0016529 | Sarcoplasmic reticulum | 6 | 3.99 × 10−8 | 7.399562462 |
| GO:0042383 | Sarcolemma | 6 | 6.56 × 10−8 | 7.18327432 |
| GO:0051480 | Regulation of cytosolic calcium ion concentration | 8 | 3.05 × 10−7 | 6.516291269 |
| GO:0006874 | Cellular calcium ion homeostasis | 7 | 0.00000312 | 5.505384291 |
| GO:0070588 | Calcium ion transmembrane transport | 5 | 0.0000635 | 4.197087214 |
| GO:0005267 | Potassium channel activity | 4 | 0.002683985 | 2.571219909 |
| GO:0005217 | Intracellular ligand-gated ion channel activity | 2 | 0.004871735 | 2.312316306 |
| GO:0006939 | Smooth muscle contraction | 2 | 0.005857764 | 2.232268112 |
| GO:0005790 | Smooth endoplasmic reticulum | 2 | 0.007141408 | 2.146216169 |
| GO:0015081 | Sodium ion transmembrane transporter activity | 2 | 0.017072923 | 1.76769213 |
| MT Downregulated Gene Ontology (GO) | ||||
| Pathway ID | Pathway Description | Gene Count | Adjusted p-Value | Log10 |
| GO:0005267 | Potassium channel activity | 9 | 3.84 × 10−9 | 8.415844939 |
| GO:0006813 | Potassium ion transport | 10 | 8.24 × 10−9 | 8.084282366 |
| GO:0016529 | Sarcoplasmic reticulum | 7 | 9.40 × 10−9 | 8.027036253 |
| GO:0005887 | Integral component of plasma membrane | 20 | 0.00000197 | 5.704434553 |
| GO:0005789 | Endoplasmic reticulum membrane | 11 | 0.000418 | 3.37836809 |
| GO:0015085 | Calcium ion transmembrane transporter activity | 3 | 0.010602036 | 1.974610737 |
| GO:0005790 | Smooth endoplasmic reticulum | 2 | 0.018509381 | 1.732608106 |
| GO:0005355 | Glucose transmembrane transporter activity | 2 | 0.019938461 | 1.700308368 |
| AT Downregulated Gene Ontology (GO) | ||||
| Pathway ID | Pathway Description | Gene Count | Adjusted p-Value | Log10 |
| GO:0005887 | Integral component of plasma membrane | 25 | 2.28 × 10−10 | 9.642075486 |
| GO:0098662 | Inorganic cation transmembrane transport | 13 | 1.98 × 10−9 | 8.703210974 |
| GO:0006813 | Potassium ion transport | 10 | 2.03 × 10−9 | 8.693400143 |
| GO:0010232 | Vascular transport | 6 | 0.0000439 | 4.357272375 |
| GO:0035725 | Sodium ion transmembrane transport | 6 | 0.0000439 | 4.357272375 |
| GO:0005267 | Potassium channel activity | 6 | 0.0001 | 3.999508817 |
| GO:0016529 | Sarcoplasmic reticulum | 4 | 0.000734 | 3.134317147 |
| GO:0015079 | Potassium ion transmembrane transporter activity | 3 | 0.003470428 | 2.459616983 |
| mPIN Downregulated Gene Ontology (GO) | ||||
| Pathway ID | Pathway Description | Gene Count | Adjusted p-Value | Log10 |
| GO:0062023 | Collagen-containing extracellular matrix | 6 | 0.000153 | 3.814225913 |
| GO:0008237 | Metallopeptidase activity | 3 | 0.008113586 | 2.090787142 |
| GO:0032027 | Myosin light chain binding | 1 | 0.049310437 | 1.307061149 |
| GO:0008270 | Zinc ion binding | 3 | 0.049310437 | 1.307061149 |
| MT Downregulated Gene Ontology (GO) | ||||
| Pathway ID | Pathway Description | Gene Count | Adjusted p-Value | Log10 |
| GO:0062023 | Collagen-containing extracellular matrix | 7 | 0.0000450 | 4.34654643 |
| GO:0005604 | Basement membrane | 2 | 0.037653103 | 1.424199233 |
| GO:1903561 | extracellular vesicle | 2 | 0.037653103 | 1.424199233 |
| AT Downregulated Gene Ontology (GO) | ||||
| Pathway ID | Pathway Description | Gene Count | Adjusted p-Value | Log10 |
| GO:0062023 | Collagen-containing extracellular matrix | 6 | 0.000226 | 3.645111972 |
| GO:0001823 | Mesonephros development | 2 | 0.028927408 | 1.538690472 |
| GO:0004518 | Nuclease activity | 2 | 0.048024251 | 1.318539402 |
| GO:0004540 | Ribonuclease activity | 2 | 0.048024251 | 1.318539402 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, N.J.; Camargo, A.C.L.; Carvalho, H.F.; Justulin, L.A.; Felisbino, S.L. Prostate Cancer Secretome and Membrane Proteome from Pten Conditional Knockout Mice Identify Potential Biomarkers for Disease Progression. Int. J. Mol. Sci. 2022, 23, 9224. https://doi.org/10.3390/ijms23169224
Santos NJ, Camargo ACL, Carvalho HF, Justulin LA, Felisbino SL. Prostate Cancer Secretome and Membrane Proteome from Pten Conditional Knockout Mice Identify Potential Biomarkers for Disease Progression. International Journal of Molecular Sciences. 2022; 23(16):9224. https://doi.org/10.3390/ijms23169224
Chicago/Turabian StyleSantos, Nilton J., Ana Carolina Lima Camargo, Hernandes F. Carvalho, Luis Antonio Justulin, and Sérgio Luis Felisbino. 2022. "Prostate Cancer Secretome and Membrane Proteome from Pten Conditional Knockout Mice Identify Potential Biomarkers for Disease Progression" International Journal of Molecular Sciences 23, no. 16: 9224. https://doi.org/10.3390/ijms23169224
APA StyleSantos, N. J., Camargo, A. C. L., Carvalho, H. F., Justulin, L. A., & Felisbino, S. L. (2022). Prostate Cancer Secretome and Membrane Proteome from Pten Conditional Knockout Mice Identify Potential Biomarkers for Disease Progression. International Journal of Molecular Sciences, 23(16), 9224. https://doi.org/10.3390/ijms23169224