
Unveiling the Ability of Witch Hazel (Hamamelis virginiana L.) Bark Extract to Impair Keratinocyte Inflammatory Cascade Typical of Atopic Eczema

 , , , ,
, , , ,  and
and 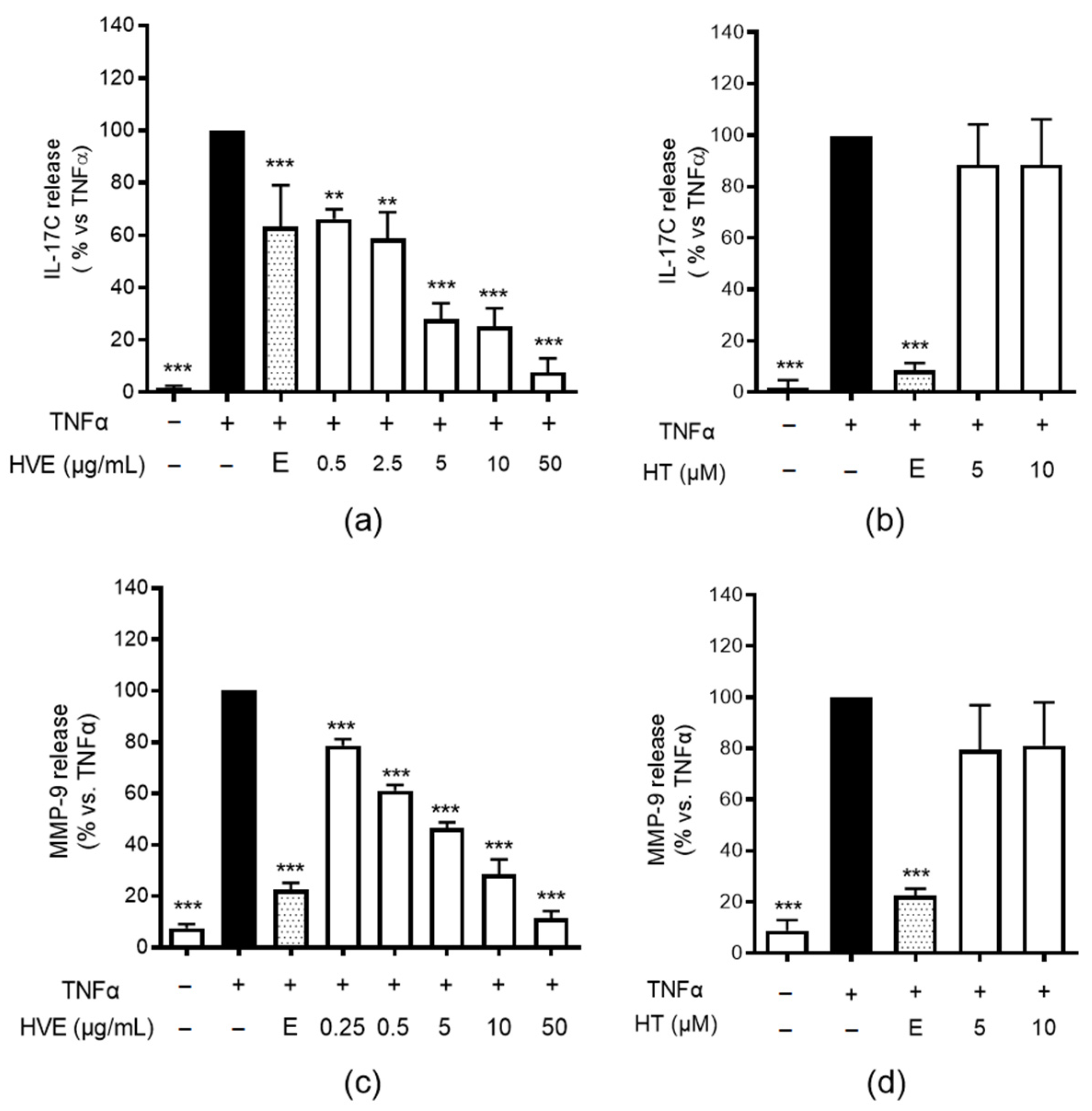{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. HVE, but Not HT, Inhibits IL-17C and MMP-9 Release in HaCaT Cells
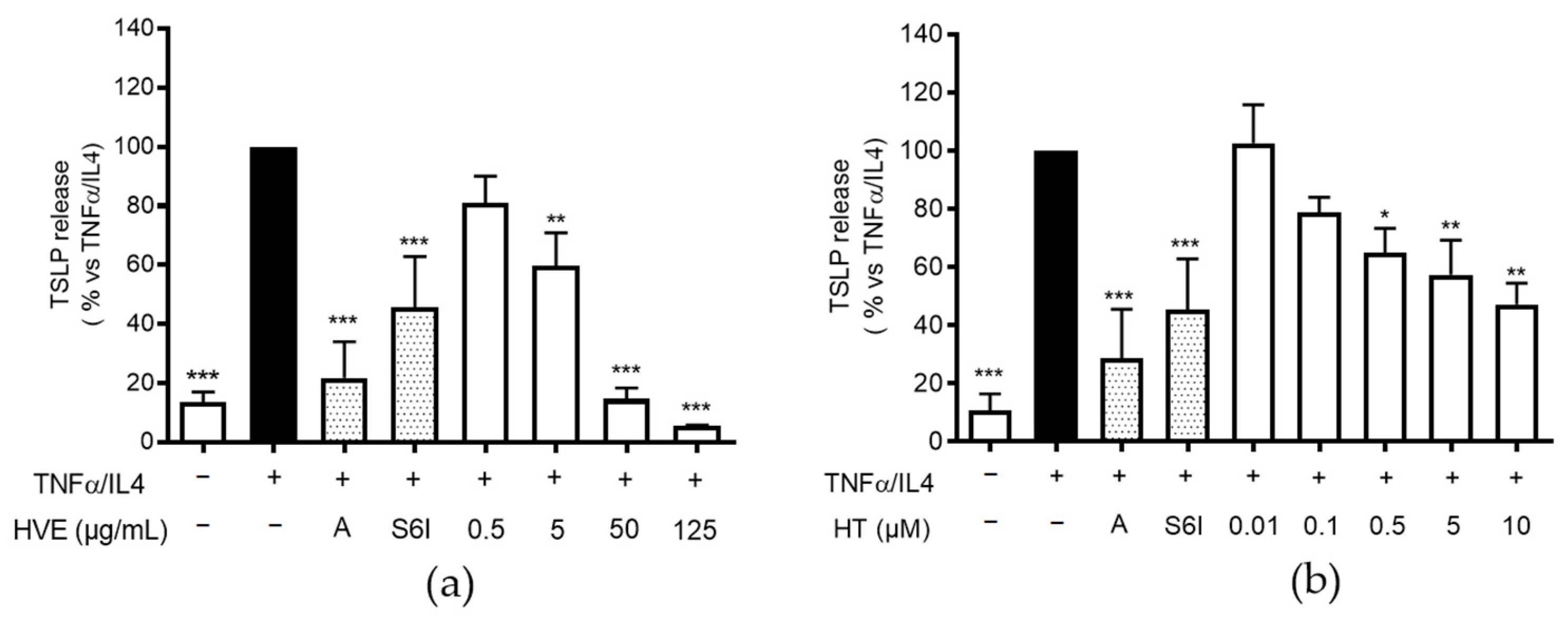
2.2. HVE and HT Inhibit TSLP Release in HaCaT Cells
2.3. HVE, but Not HT, Inhibits IL-6 Release in HaCaT Cells
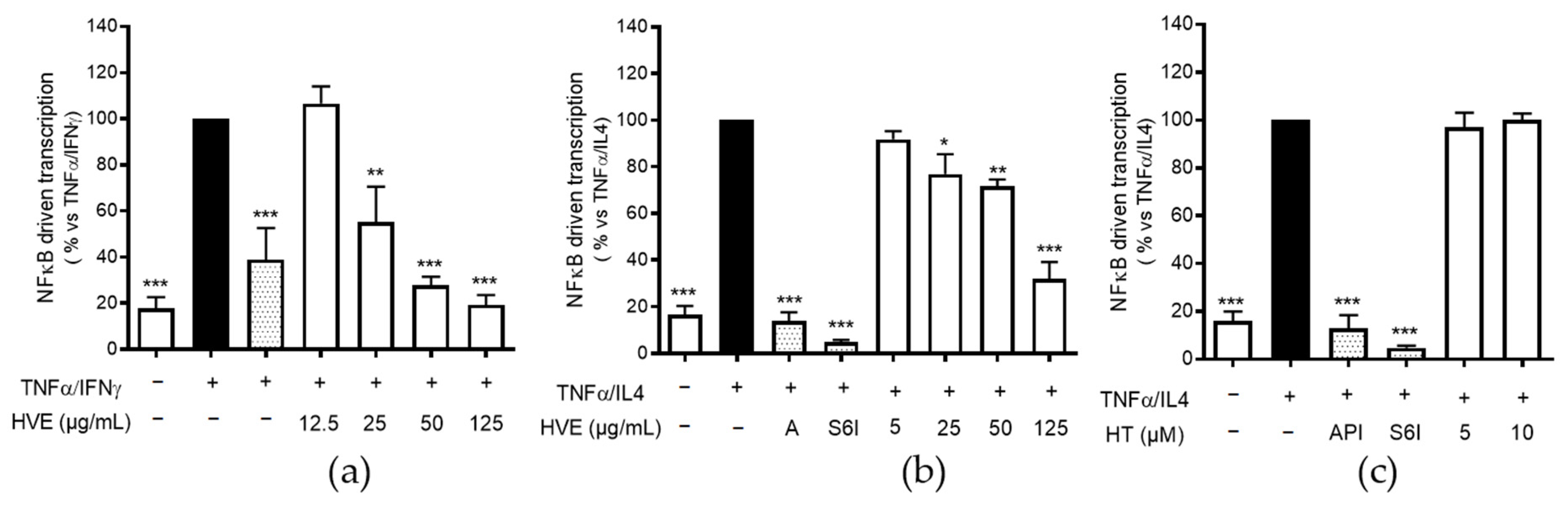
2.4. HVE, but Not HT, Inhibits the NF-κB-Driven Transcription
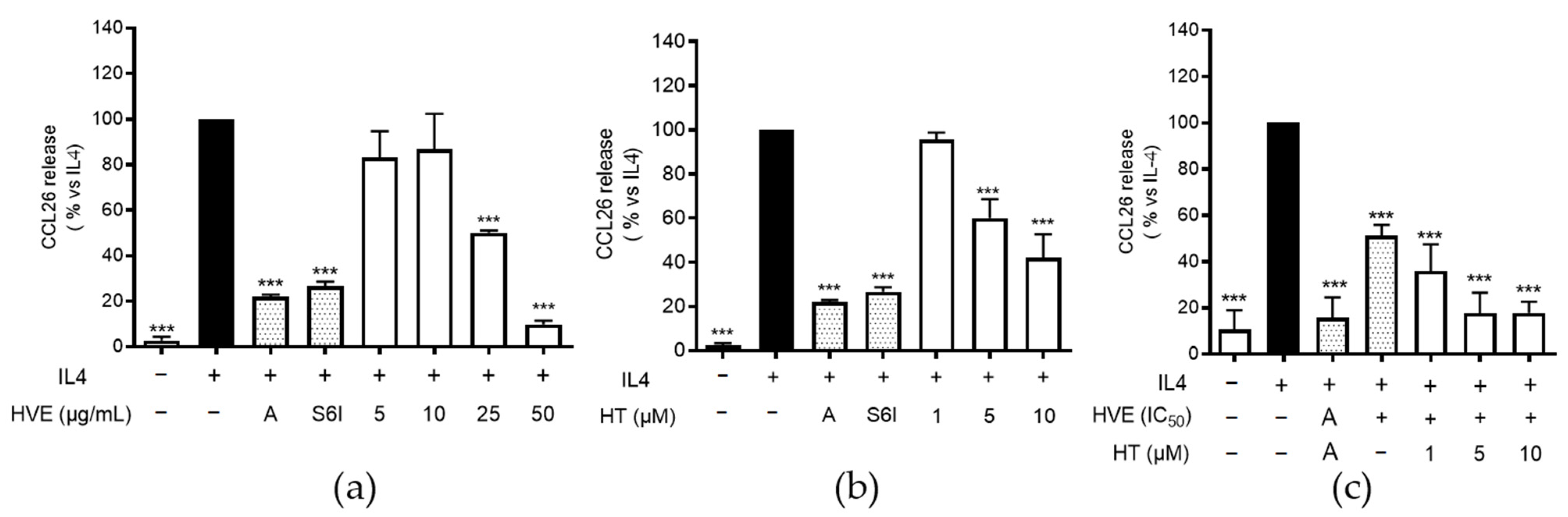
2.5. HVE and HT Inhibit IL-4-Induced CCL26 in HaCaT Cells
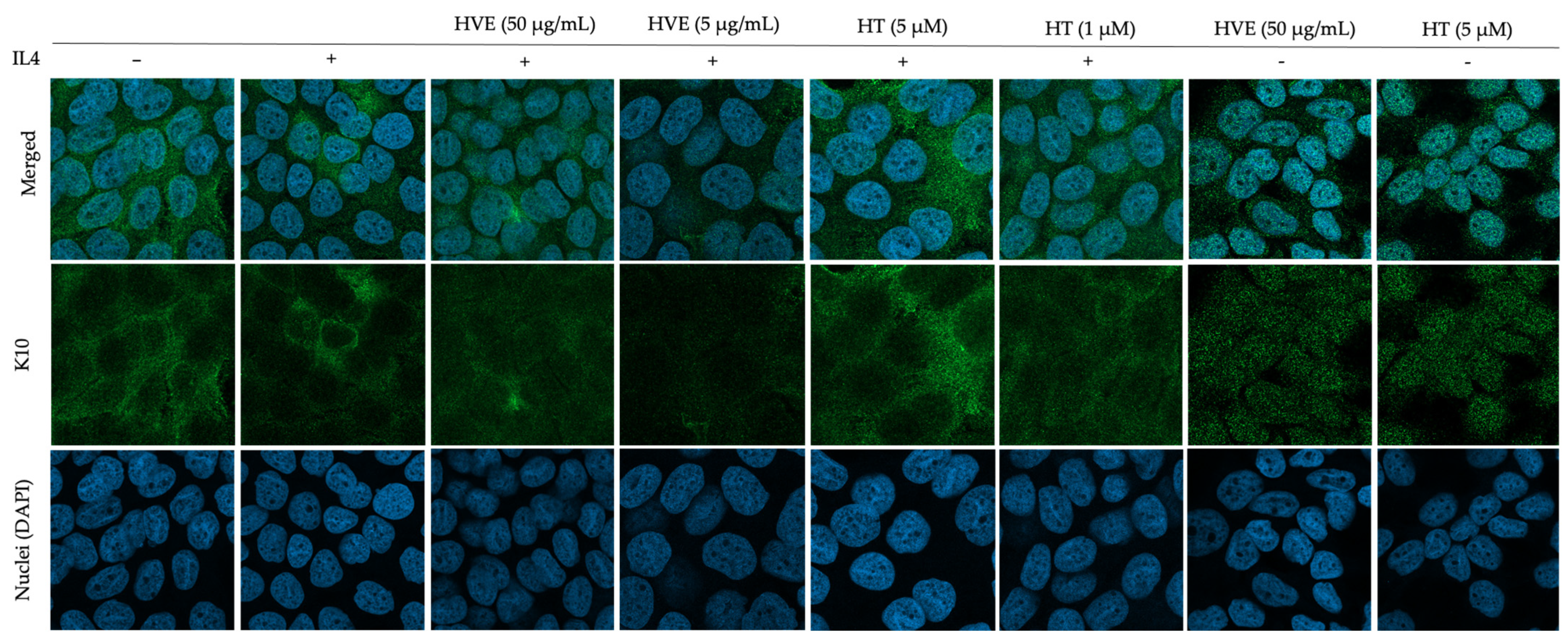
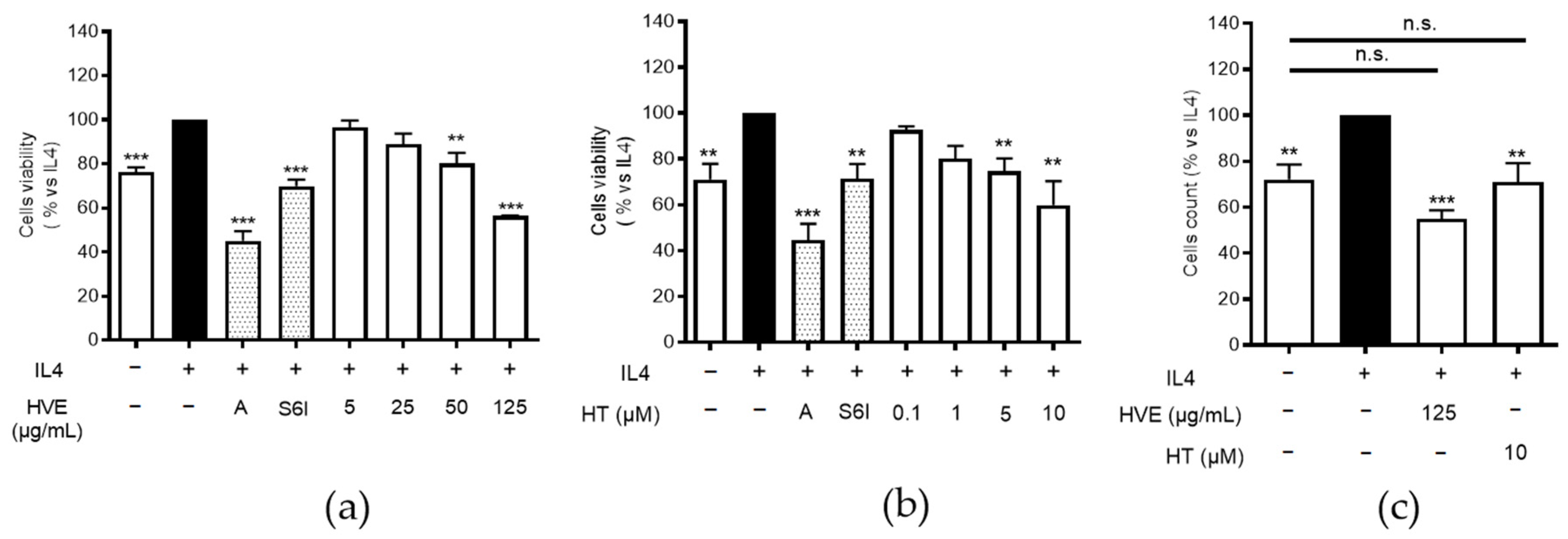
2.6. HVE and HT Inhibit IL4-Induced Proliferation and Recover K10 Expression
3. Discussion
4. Materials and Methods
4.1. HaCaT Cell Culture and Differentiation
4.2. Evaluation of Cell Viability and Proliferation
4.3. Evaluation of Cytokeratin 10 (K10) and Involucrin (IVN) Expression via Immunofluorescence
4.4. Measurement of Cytokine Release with the ELISA Assay
- -
- IL-17C and MMP-9 release was stimulated with TNF-α (10 ng/mL) in undifferentiated HaCaT cells;
- -
- TSLP and IL-6 release was stimulated with TNF-α (10 ng/mL)/IL-4 (100 ng/mL) combination in differentiated HaCaT cells, and IL-6 was also evaluated after TNF-α (10 ng/mL)/IFN-γ (5 ng/mL) stimulation in undifferentiated HaCaT cells, according to our previously published time-course experiments [44];
- -
- CCL26 release was stimulated with IL-4 (100 ng/mL) in differentiated HaCaT cells.
4.5. Measurement of the NF-κB-Driven Transcription
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- European Medicines Agency Committeeon Herbal Medicinal Products (HMPC). Assessment re-port on Hamamelis virginiana L., cortex; Hamamelis virginiana L., folium; Hamamelis virginiana L., folium et cortex aut ramunculus destillatum, EMA/HMPC/114585/2008. Available online: http://www.ema.europa.eu/docs/en_GB/document_library/Herbal_-_HMPC_assessment_report/2010/04/WC500089242.pdf (accessed on 11 July 2022).
- ESCOP. Escop Monographs: The Scientific Foundation for Herbal Medicinal Products, 2nd ed.; Georg Thieme Verlag: New York, NY, USA, 2009. [Google Scholar]
- Hughes-Formella, B.J.; Bohnsack, K.; Rippke, F.; Benner, G.; Rudolph, M.; Tausch, I.; Gassmueller, J. Anti-inflammatory effect of hamamelis lotion in a UVB erythema test. Dermatology 1998, 196, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Korting, H.C.; Schafer-Korting, M.; Klovekorn, W.; Klovekorn, G.; Martin, C.; Laux, P. Comparative efficacy of hamamelis distillate and hydrocortisone cream in atopic eczema. Eur. J. Clin. Pharmacol. 1995, 48, 461–465. [Google Scholar] [CrossRef] [PubMed]
- Wolff, H.H.; Kieser, M. Hamamelis in children with skin disorders and skin injuries: Results of an observational study. Eur. J. Pediatr. 2007, 166, 943–948. [Google Scholar] [CrossRef] [PubMed]
- Duwiejua, M.; Zeitlin, I.J.; Waterman, P.G.; Gray, A.I. Anti-inflammatory activity of Polygonum bistorta, Guaiacum officinale and Hamamelis virginiana in rats. J. Pharm. Pharmacol. 1994, 46, 286–290. [Google Scholar] [CrossRef] [PubMed]
- Erdelmeier, C.A.J.; Cinatl, J.; Rabenau, H.; Doerr, H.W.; Biber, A.; Koch, E. Antiviral and antiphiogistic activities of hamamelis virginiana bark. Planta Med. 1995, 62, 241–245. [Google Scholar] [CrossRef]
- Wang, H.; Provan, G.J.; Helliwell, K. Determination of hamamelitannin, catechins and gallic acid in witch hazel bark, twig and leaf by HPLC. J. Pharm. Biomed. Anal. 2003, 33, 539–544. [Google Scholar] [CrossRef]
- Theisen, L.L.; Erdelmeier, C.A.; Spoden, G.A.; Boukhallouk, F.; Sausy, A.; Florin, L.; Muller, C.P. Tannins from Hamamelis virginiana bark extract: Characterization and improvement of the antiviral efficacy against influenza A virus and human papillomavirus. PLoS ONE 2014, 9, e88062. [Google Scholar] [CrossRef]
- Masaki, H.; Atsumi, T.; Sakurai, H. Protective activity of hamamelitannin on cell damage of murine skin fibroblasts induced by UVB irradiation. J. Dermatol. Sci. 1995, 10, 25–34. [Google Scholar] [CrossRef]
- Cheesman, M.J.; Alcorn, S.; Verma, V.; Cock, I.E. An assessment of the growth inhibition profiles of Hamamelis virginiana L. extracts against Streptococcus and Staphylococcus spp. J. Tradit. Complement. Med. 2021, 11, 457–465. [Google Scholar] [CrossRef]
- Hartisch, C.; Kolodziej, H.; von Bruchhausen, F. Dual inhibitory activities of tannins from Hamamelis virginiana and related polyphenols on 5-lipoxygenase and lyso-PAF: Acetyl-CoA acetyltransferase. Planta Med. 1997, 63, 106–110. [Google Scholar] [CrossRef]
- Piazza, S.; Martinelli, G.; Vrhovsek, U.; Masuero, D.; Fumagalli, M.; Magnavacca, A.; Pozzoli, C.; Canilli, L.; Terno, M.; Angarano, M.; et al. Anti-Inflammatory and Anti-Acne Effects of Hamamelis virginiana Bark in Human Keratinocytes. Antioxidants 2022, 11, 1119. [Google Scholar] [CrossRef] [PubMed]
- Langan, S.M.; Irvine, A.D.; Weidinger, S. Atopic dermatitis. Lancet 2020, 396, 345–360. [Google Scholar] [CrossRef]
- Francuzik, W.; Franke, K.; Schumann, R.R.; Heine, G.; Worm, M. Propionibacterium acnes Abundance Correlates Inversely with Staphylococcus aureus: Data from Atopic Dermatitis Skin Microbiome. Acta Derm.-Venereol. 2018, 98, 490–495. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Kim, J.S.; Cho, D.H.; Park, H.J. Molecular Mechanisms of Cutaneous Inflammatory Disorder: Atopic Dermatitis. Int. J. Mol. Sci. 2016, 17, 1234. [Google Scholar] [CrossRef] [PubMed]
- Corren, J.; Ziegler, S.F. TSLP: From allergy to cancer. Nat. Immunol. 2019, 20, 1603–1609. [Google Scholar] [CrossRef]
- Dao, T.T.P.; Song, K.; Kim, J.Y.; Kim, Y.S. Igalan from Inula helenium (L.) suppresses the atopic dermatitis-like response in stimulated HaCaT keratinocytes via JAK/STAT3 signaling. Inflamm. Res. 2020, 69, 309–319. [Google Scholar] [CrossRef]
- Bao, L.; Alexander, J.B.; Zhang, H.; Shen, K.; Chan, L.S. Interleukin-4 Downregulation of Involucrin Expression in Human Epidermal Keratinocytes Involves Stat6 Sequestration of the Coactivator CREB-Binding Protein. J. Interf. Cytokine Res. 2016, 36, 374–381. [Google Scholar] [CrossRef]
- Gruber, R.; Bornchen, C.; Rose, K.; Daubmann, A.; Volksdorf, T.; Wladykowski, E.; Vidal, Y.S.S.; Peters, E.M.; Danso, M.; Bouwstra, J.A.; et al. Diverse regulation of claudin-1 and claudin-4 in atopic dermatitis. Am. J. Pathol. 2015, 185, 2777–2789. [Google Scholar] [CrossRef]
- Bao, L.; Shi, V.Y.; Chan, L.S. IL-4 up-regulates epidermal chemotactic, angiogenic, and pro-inflammatory genes and down-regulates antimicrobial genes in vivo and in vitro: Relevant in the pathogenesis of atopic dermatitis. Cytokine 2013, 61, 419–425. [Google Scholar] [CrossRef]
- Werfel, T.; Allam, J.P.; Biedermann, T.; Eyerich, K.; Gilles, S.; Guttman-Yassky, E.; Hoetzenecker, W.; Knol, E.; Simon, H.U.; Wollenberg, A.; et al. Cellular and molecular immunologic mechanisms in patients with atopic dermatitis. J. Allerg. Clin. Immunol. 2016, 138, 336–349. [Google Scholar] [CrossRef]
- Danso, M.O.; van Drongelen, V.; Mulder, A.; van Esch, J.; Scott, H.; van Smeden, J.; El Ghalbzouri, A.; Bouwstra, J.A. TNF-alpha and Th2 cytokines induce atopic dermatitis-like features on epidermal differentiation proteins and stratum corneum lipids in human skin equivalents. J. Investig. Dermatol. 2014, 134, 1941–1950. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Baek, J.; Lee, J.R.; Roh, J.Y.; Jung, Y. Optimization of Cytokine Milieu to Reproduce Atopic Dermatitis-related Gene Expression in HaCaT Keratinocyte Cell Line. Immun. Netw. 2018, 18, e9. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.E.; Ha, H.; Shin, H.K.; Seo, C.S. Anti-Allergic and Anti-Inflammatory Effects of Kuwanon G and Morusin on MC/9 Mast Cells and HaCaT Keratinocytes. Molecules 2019, 24, 265. [Google Scholar] [CrossRef]
- Gadina, M.; Gazaniga, N.; Vian, L.; Furumoto, Y. Small molecules to the rescue: Inhibition of cytokine signaling in immune-mediated diseases. J Autoim. 2017, 85, 20–31. [Google Scholar] [CrossRef]
- Piazza, S.; Fumagalli, M.; Khalilpour, S.; Martinelli, G.; Magnavacca, A.; Dell’Agli, M.; Sangiovanni, E. A Review of the Potential Benefits of Plants Producing Berries in Skin Disorders. Antioxidants 2020, 9, 542. [Google Scholar] [CrossRef] [PubMed]
- Park, C.H.; Min, S.Y.; Yu, H.W.; Kim, K.; Kim, S.; Lee, H.J.; Kim, J.H.; Park, Y.J. Effects of Apigenin on RBL-2H3, RAW264.7, and HaCaT Cells: Anti-Allergic, Anti-Inflammatory, and Skin-Protective Activities. Int. J. Mol. Sci. 2020, 21, 4620. [Google Scholar] [CrossRef] [PubMed]
- Pang, L.; Zou, S.; Shi, Y.; Mao, Q.; Chen, Y. Apigenin attenuates PM2.5-induced airway hyperresponsiveness and inflammation by down-regulating NF-kappaB in murine model of asthma. Int. J. Clin. Exp. Pathol. 2019, 12, 3700–3709. [Google Scholar]
- Pappu, R.; Rutz, S.; Ouyang, W. Regulation of epithelial immunity by IL-17 family cytokines. Trends Immunol. 2012, 33, 343–349. [Google Scholar] [CrossRef]
- Gittler, J.K.; Shemer, A.; Suarez-Farinas, M.; Fuentes-Duculan, J.; Gulewicz, K.J.; Wang, C.Q.; Mitsui, H.; Cardinale, I.; de Guzman Strong, C.; Krueger, J.G.; et al. Progressive activation of T(H)2/T(H)22 cytokines and selective epidermal proteins characterizes acute and chronic atopic dermatitis. J. Allerg. Clin. Immunol. 2012, 130, 1344–1354. [Google Scholar] [CrossRef]
- Johnston, A.; Fritz, Y.; Dawes, S.M.; Diaconu, D.; Al-Attar, P.M.; Guzman, A.M.; Chen, C.S.; Fu, W.; Gudjonsson, J.E.; McCormick, T.S.; et al. Keratinocyte overexpression of IL-17C promotes psoriasiform skin inflammation. J. Immunol. 2013, 190, 2252–2262. [Google Scholar] [CrossRef]
- Harper, J.I.; Godwin, H.; Green, A.; Wilkes, L.E.; Holden, N.J.; Moffatt, M.; Cookson, W.O.; Layton, G.; Chandler, S. A study of matrix metalloproteinase expression and activity in atopic dermatitis using a novel skin wash sampling assay for functional biomarker analysis. Br. J. Dermatol. 2010, 162, 397–403. [Google Scholar] [CrossRef]
- Wilson, S.R.; The, L.; Batia, L.M.; Beattie, K.; Katibah, G.E.; McClain, S.P.; Pellegrino, M.; Estandian, D.M.; Bautista, D.M. The epithelial cell-derived atopic dermatitis cytokine TSLP activates neurons to induce itch. Cell 2013, 155, 285–295. [Google Scholar] [CrossRef]
- Worm, M.; Francuzik, W.; Kraft, M.; Alexiou, A. Modern therapies in atopic dermatitis: Biologics and small molecule drugs. J. Dtsch. Dermatol. Ges. 2020, 18, 1085–1092. [Google Scholar] [CrossRef]
- Bogiatzi, S.I.; Fernandez, I.; Bichet, J.C.; Marloie-Provost, M.A.; Volpe, E.; Sastre, X.; Soumelis, V. Cutting Edge: Proinflammatory and Th2 cytokines synergize to induce thymic stromal lymphopoietin production by human skin keratinocytes. J. Immunol. 2007, 178, 3373–3377. [Google Scholar] [CrossRef] [PubMed]
- Kao, J.K.; Lee, C.H.; Lee, M.S.; Hsu, C.S.; Tsao, L.Y.; Tsai, Y.G.; Shieh, J.J.; Yang, R.C. Heat-shock pretreatment reduces expression and release of TSLP from keratinocytes under Th2 environment. Pediatr. Allerg. Immunol. 2016, 27, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Vu, A.T.; Chen, X.; Xie, Y.; Kamijo, S.; Ushio, H.; Kawasaki, J.; Hara, M.; Ikeda, S.; Okumura, K.; Ogawa, H.; et al. Extracellular double-stranded RNA induces TSLP via an endosomal acidification- and NF-kappaB-dependent pathway in human keratinocytes. J. Investig. Dermatol. 2011, 131, 2205–2212. [Google Scholar] [CrossRef] [PubMed]
- Derocq, J.M.; Segui, M.; Poinot-Chazel, C.; Minty, A.; Caput, D.; Ferrara, P.; Casellas, P. Interleukin-13 stimulates interleukin-6 production by human keratinocytes. Similarity with interleukin-4. FEBS Lett. 1994, 343, 32–36. [Google Scholar] [CrossRef]
- Diehl, S.; Rincon, M. The two faces of IL-6 on Th1/Th2 differentiation. Mol. Immunol. 2002, 39, 531–536. [Google Scholar] [CrossRef]
- Son, E.D.; Kim, H.J.; Kim, K.H.; Bin, B.H.; Bae, I.H.; Lim, K.M.; Yu, S.J.; Cho, E.G.; Lee, T.R. S100A7 (psoriasin) inhibits human epidermal differentiation by enhanced IL-6 secretion through IkappaB/NF-kappaB signalling. Exp Dermatol. 2016, 25, 636–641. [Google Scholar] [CrossRef]
- Kohda, F.; Koga, T.; Uchi, H.; Urabe, K.; Furue, M. Histamine-induced IL-6 and IL-8 production are differentially modulated by IFN-gamma and IL-4 in human keratinocytes. J. Dermatol. Sci. 2002, 28, 34–41. [Google Scholar] [CrossRef]
- Beken, B.; Serttas, R.; Yazicioglu, M.; Turkekul, K.; Erdogan, S. Quercetin Improves Inflammation, Oxidative Stress, and Impaired Wound Healing in Atopic Dermatitis Model of Human Keratinocytes. Pediatr. Allerg. Immunol. Pulmonol. 2020, 33, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Magnavacca, A.; Piazza, S.; Cammisa, A.; Fumagalli, M.; Martinelli, G.; Giavarini, F.; Sangiovanni, E.; Dell’Agli, M. Ribes nigrum Leaf Extract Preferentially Inhibits IFN-gamma-Mediated Inflammation in HaCaT Keratinocytes. Molecules 2021, 26, 3044. [Google Scholar] [CrossRef] [PubMed]
- Omori-Miyake, M.; Yamashita, M.; Tsunemi, Y.; Kawashima, M.; Yagi, J. In vitro assessment of IL-4- or IL-13-mediated changes in the structural components of keratinocytes in mice and humans. J. Investig. Dermatol. 2014, 134, 1342–1350. [Google Scholar] [CrossRef]
- Serezani, A.P.M.; Bozdogan, G.; Sehra, S.; Walsh, D.; Krishnamurthy, P.; Sierra Potchanant, E.A.; Nalepa, G.; Goenka, S.; Turner, M.J.; Spandau, D.F.; et al. IL-4 impairs wound healing potential in the skin by repressing fibronectin expression. J. Allerg. Clin. Immunol. 2017, 139, 142–151.e5. [Google Scholar] [CrossRef]
- Kagami, S.; Saeki, H.; Komine, M.; Kakinuma, T.; Tsunemi, Y.; Nakamura, K.; Sasaki, K.; Asahina, A.; Tamaki, K. Interleukin-4 and interleukin-13 enhance CCL26 production in a human keratinocyte cell line, HaCaT cells. Clin. Exp. Immunol. 2005, 141, 459–466. [Google Scholar] [CrossRef]
- Bao, L.; Shi, V.Y.; Chan, L.S. IL-4 regulates chemokine CCL26 in keratinocytes through the Jak1, 2/Stat6 signal transduction pathway: Implication for atopic dermatitis. Mol. Immunol. 2012, 50, 91–97. [Google Scholar] [CrossRef]
- Koller, F.L.; Hwang, D.G.; Dozier, E.A.; Fingleton, B. Epithelial interleukin-4 receptor expression promotes colon tumor growth. Carcinogenesis 2010, 31, 1010–1017. [Google Scholar] [CrossRef]
- Shi, V.Y.; Bao, L.; Chan, L.S. Inflammation-driven dermal lymphangiogenesis in atopic dermatitis is associated with CD11b+ macrophage recruitment and VEGF-C up-regulation in the IL-4-transgenic mouse model. Microcirculation 2012, 19, 567–579. [Google Scholar] [CrossRef]
- Lee, S.; Jegal, H.; Bong, S.K.; Yoon, K.N.; Park, N.J.; Shin, M.S.; Yang, M.H.; Kim, Y.K.; Kim, S.N. Anti-Atopic Effect of Acorn Shell Extract on Atopic Dermatitis-Like Lesions in Mice and Its Active Phytochemicals. Biomolecules 2019, 10, 57. [Google Scholar] [CrossRef]
- Choi, J.; Yang, D.; Moon, M.Y.; Han, G.Y.; Chang, M.S.; Cha, J. The Protective Effect of Hamamelis virginiana Stem and Leaf Extract on Fine Dust-Induced Damage on Human Keratinocytes. Cosmetics 2021, 8, 119. [Google Scholar] [CrossRef]
- Kim, Y.H.; Yoshimoto, M.; Nakayama, K.; Tanino, S.; Fujimura, Y.; Yamada, K.; Tachibana, H. Tannic acid, a higher galloylated pentagalloylglucose, suppresses antigen-specific IgE production by inhibiting varepsilon germline transcription induced by STAT6 activation. FEBS Open Bio 2013, 3, 341–345. [Google Scholar] [CrossRef]
- Coleman, S.L.; Shaw, O.M. Progress in the understanding of the pathology of allergic asthma and the potential of fruit proanthocyanidins as modulators of airway inflammation. Food Funct. 2017, 8, 4315–4324. [Google Scholar] [CrossRef]
- Colombo, I.; Sangiovanni, E.; Maggio, R.; Mattozzi, C.; Zava, S.; Corbett, Y.; Fumagalli, M.; Carlino, C.; Corsetto, P.A.; Scaccabarozzi, D.; et al. HaCaT Cells as a Reliable In Vitro Differentiation Model to Dissect the Inflammatory/Repair Response of Human Keratinocytes. Mediat. Inflamm. 2017, 2017, 7435621. [Google Scholar] [CrossRef]
- Bellosta, S.; Dell’Agli, M.; Canavesi, M.; Mitro, N.; Monetti, M.; Crestani, M.; Verotta, L.; Fuzzati, N.; Bernini, F.; Bosisio, E. Inhibition of metalloproteinase-9 activity and gene expression by polyphenolic compounds isolated from the bark of Tristaniopsis calobuxus (Myrtaceae). Cell. Mol. Life Sci. 2003, 60, 1440–1448. [Google Scholar] [CrossRef]
- Dell’agli, M.; Bellosta, S.; Rizzi, L.; Galli, G.V.; Canavesi, M.; Rota, F.; Parente, R.; Bosisio, E.; Romeo, S. A structure-activity study for the inhibition of metalloproteinase-9 activity and gene expression by analogues of gallocatechin-3-gallate. Cell. Mol. Life Sci. 2005, 62, 2896–2903. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piazza, S.; Martinelli, G.; Magnavacca, A.; Fumagalli, M.; Pozzoli, C.; Terno, M.; Canilli, L.; Angarano, M.; Maranta, N.; Dell’Agli, M.; et al. Unveiling the Ability of Witch Hazel (Hamamelis virginiana L.) Bark Extract to Impair Keratinocyte Inflammatory Cascade Typical of Atopic Eczema. Int. J. Mol. Sci. 2022, 23, 9279. https://doi.org/10.3390/ijms23169279
Piazza S, Martinelli G, Magnavacca A, Fumagalli M, Pozzoli C, Terno M, Canilli L, Angarano M, Maranta N, Dell’Agli M, et al. Unveiling the Ability of Witch Hazel (Hamamelis virginiana L.) Bark Extract to Impair Keratinocyte Inflammatory Cascade Typical of Atopic Eczema. International Journal of Molecular Sciences. 2022; 23(16):9279. https://doi.org/10.3390/ijms23169279
Chicago/Turabian StylePiazza, Stefano, Giulia Martinelli, Andrea Magnavacca, Marco Fumagalli, Carola Pozzoli, Massimo Terno, Luisa Canilli, Marco Angarano, Nicole Maranta, Mario Dell’Agli, and et al. 2022. "Unveiling the Ability of Witch Hazel (Hamamelis virginiana L.) Bark Extract to Impair Keratinocyte Inflammatory Cascade Typical of Atopic Eczema" International Journal of Molecular Sciences 23, no. 16: 9279. https://doi.org/10.3390/ijms23169279
APA StylePiazza, S., Martinelli, G., Magnavacca, A., Fumagalli, M., Pozzoli, C., Terno, M., Canilli, L., Angarano, M., Maranta, N., Dell’Agli, M., & Sangiovanni, E. (2022). Unveiling the Ability of Witch Hazel (Hamamelis virginiana L.) Bark Extract to Impair Keratinocyte Inflammatory Cascade Typical of Atopic Eczema. International Journal of Molecular Sciences, 23(16), 9279. https://doi.org/10.3390/ijms23169279