
Expression of TXNRD1, HSPA4L and ATP1B1 Genes Associated with the Freezability of Boar Sperm

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Semen Quality Assessment
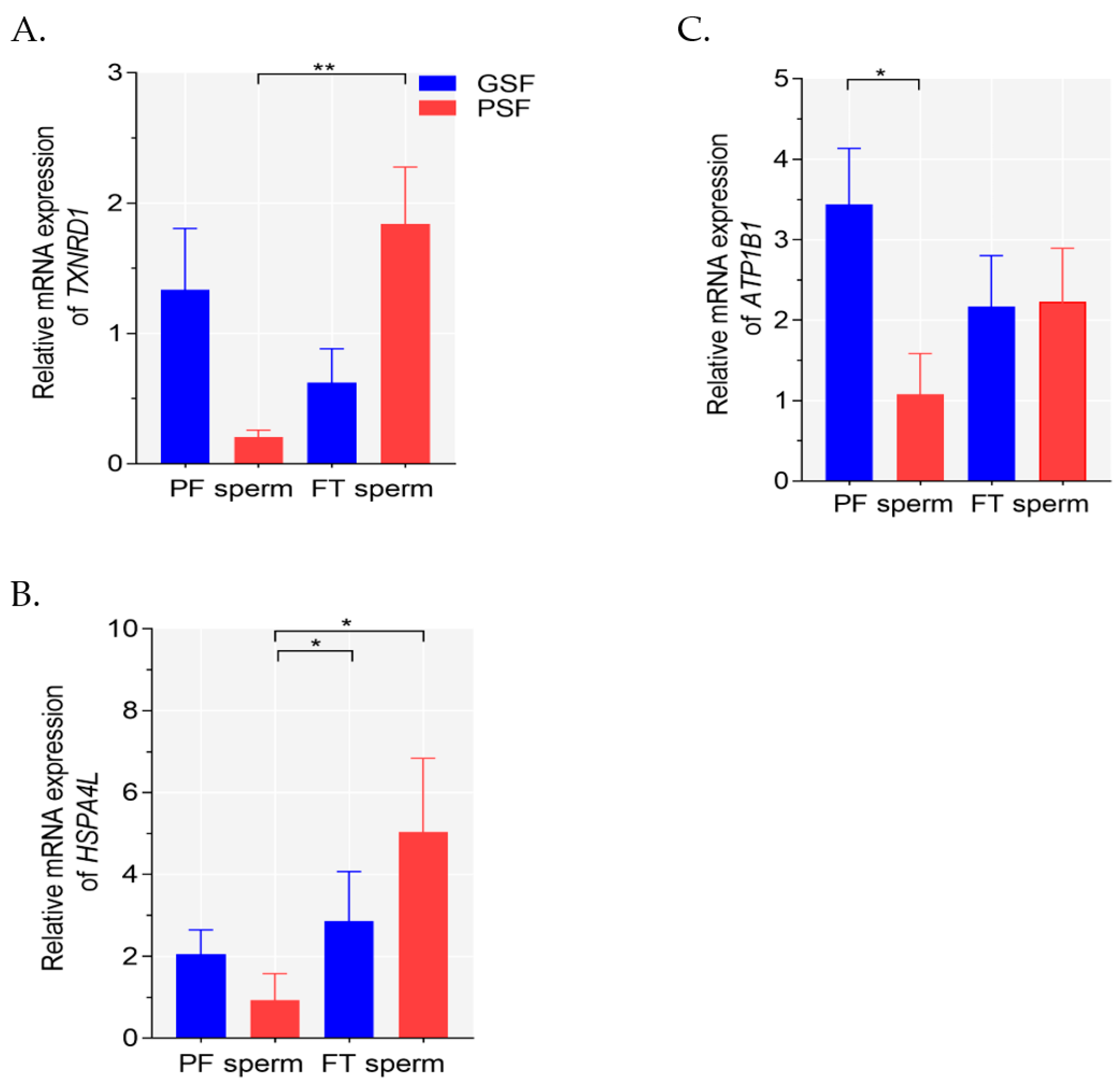
2.2. Analysis of Gene Expression
2.3. Gene Ontology (GO) Enrichment Analysis and KEGG Pathways
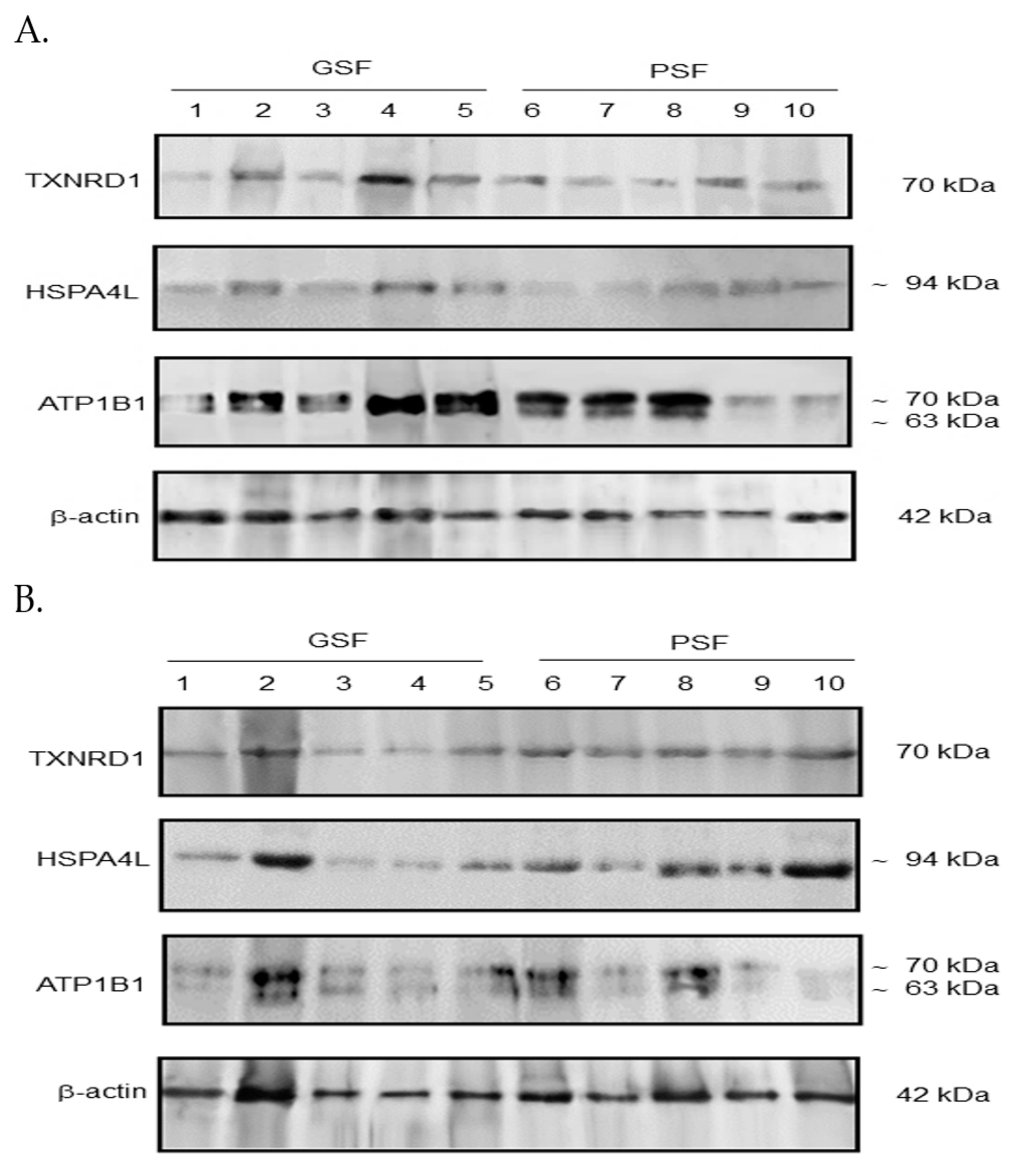
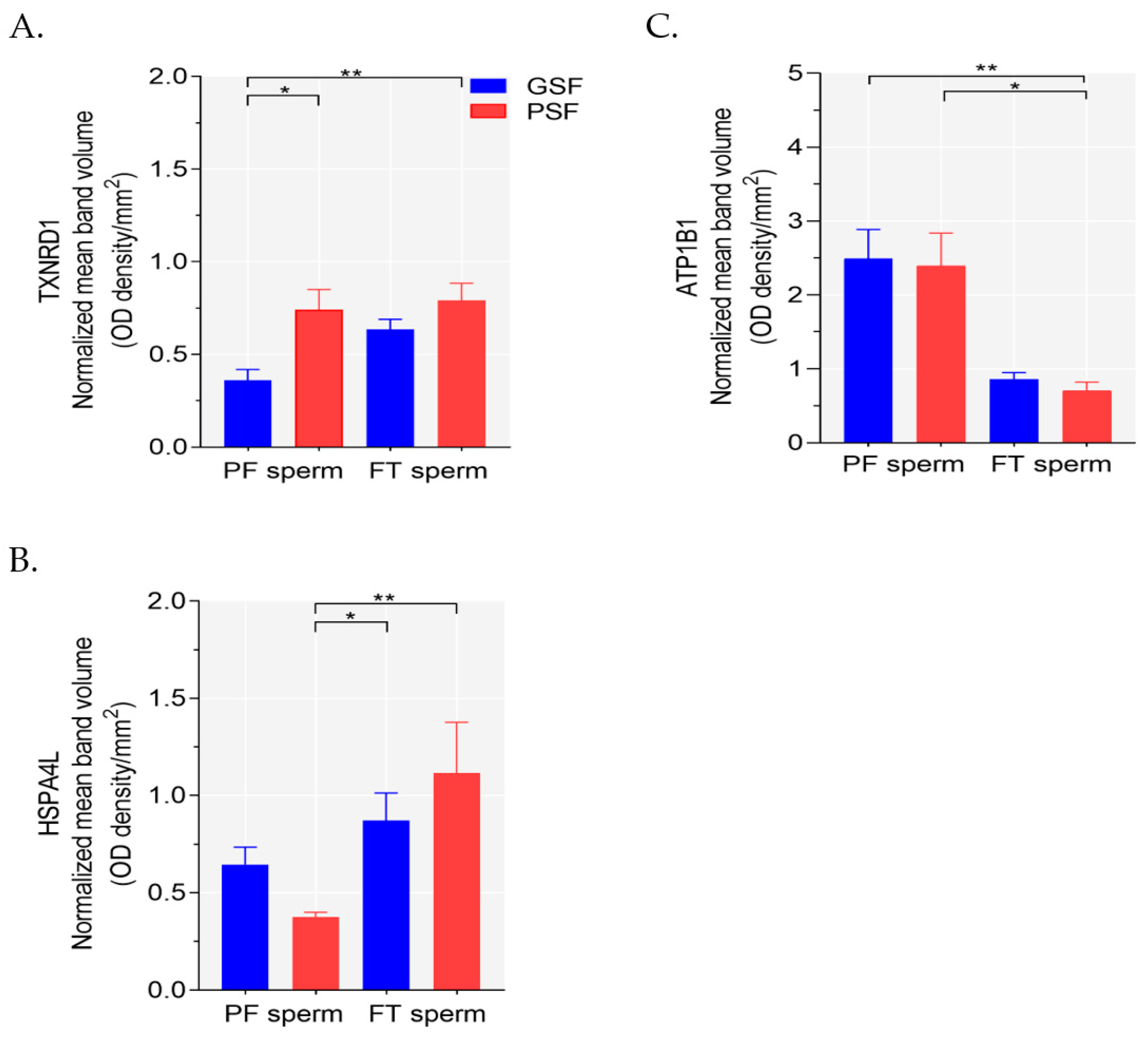
2.4. Western Blotting Analysis
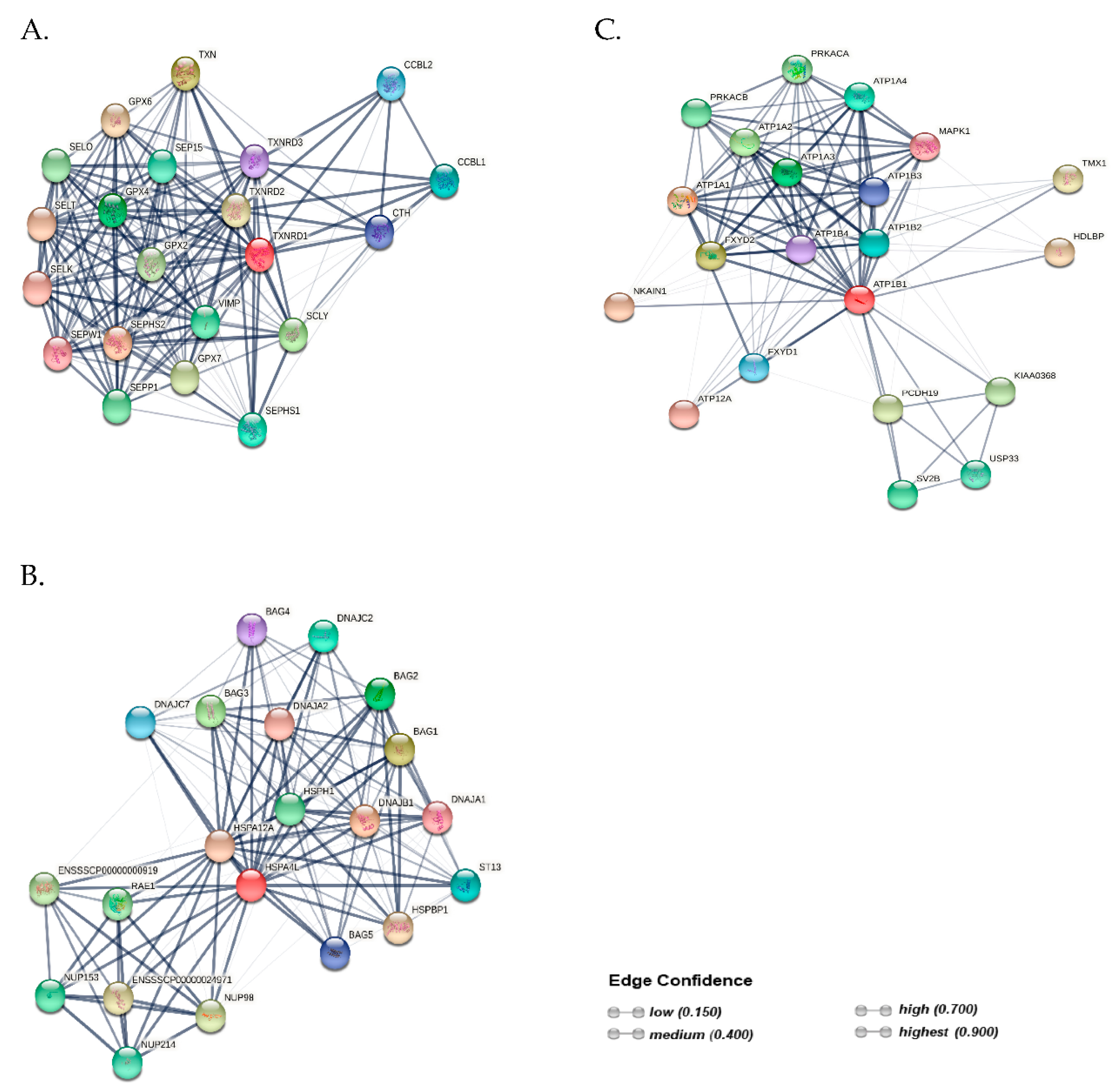
2.5. Protein–Protein Interaction (PPI) Networks
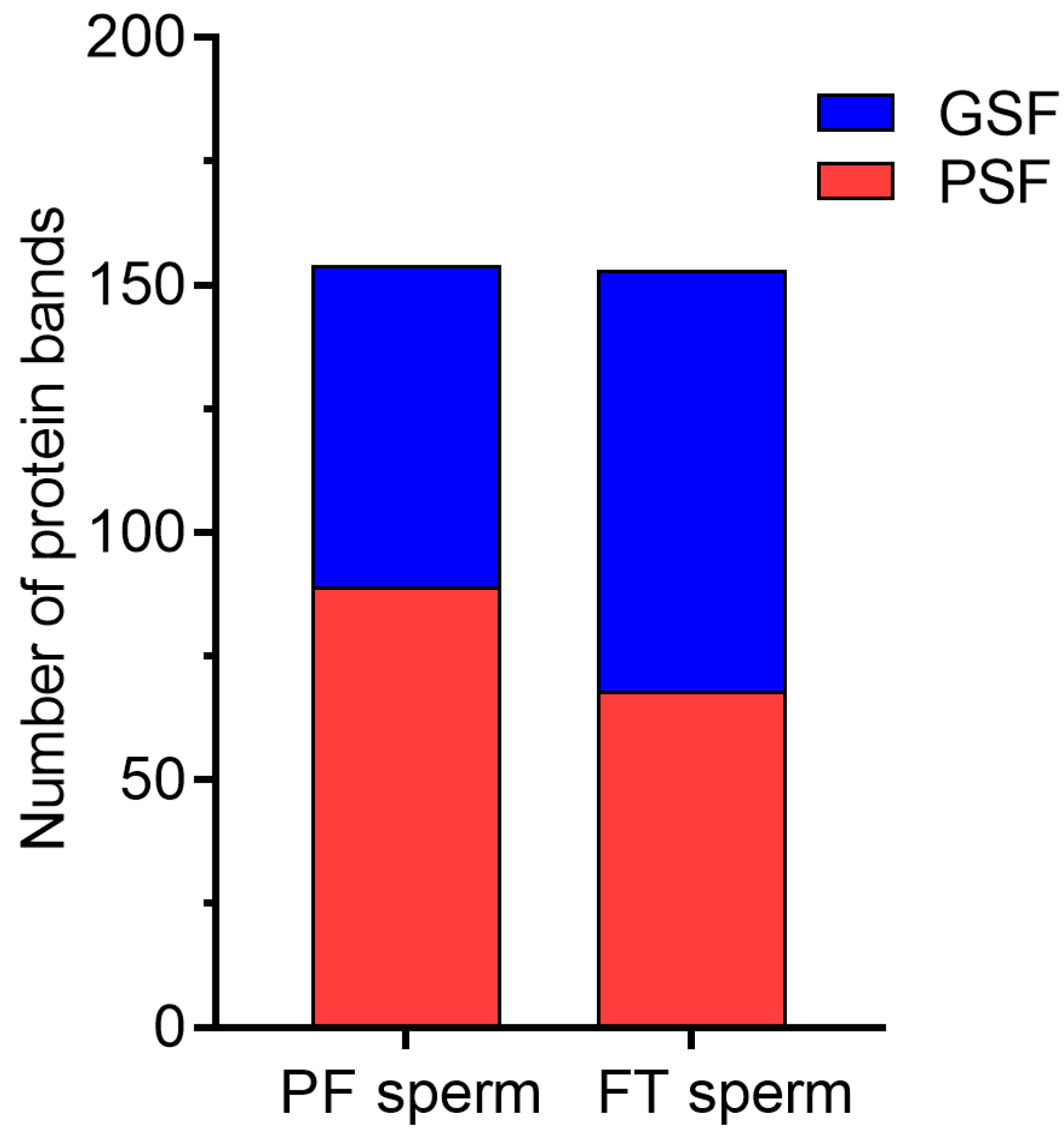
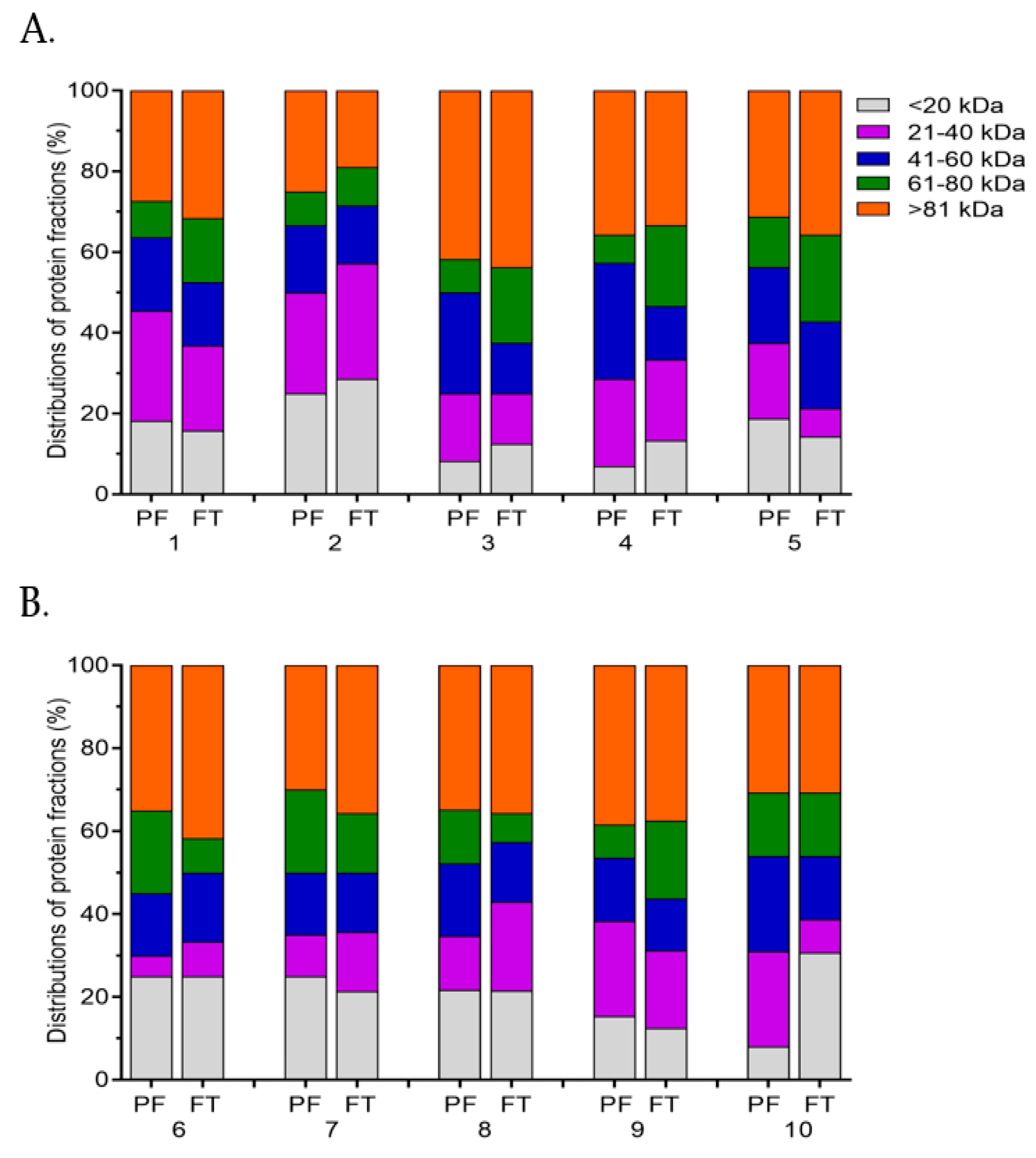
2.6. SDS-PAGE Analysis
3. Discussion
4. Materials and Methods
4.1. Chemicals and Media
4.2. Animals and Semen Collections
4.3. Semen Processing Procedure
4.4. Semen Quality Assessment
4.4.1. CASA Motility and Motion Parameters
4.4.2. Membrane Integrity Characteristics
4.4.3. DNA Fragmentation
4.5. Total RNA Isolation and Quantitative Real-Time PCR (RT-qPCR) Analysis
4.6. GO Enrichment Analysis and KEGG Pathways
4.7. Western Blotting Analysis
4.8. PPI Networks
4.9. Sodium Dodecyl Sulfate–Polyacrylamide Gel Electrophoresis (SDS-PAGE) Analysis
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Holt, W.V.; Medrano, A.; Thurston, L.M.; Watson, P.F. The significance of cooling rates and animal variability for boar sperm cryopreservation: Insights from the cryomicroscope. Theriogenology 2005, 63, 370–382. [Google Scholar] [CrossRef] [PubMed]
- Yeste, M. Sperm cryopreservation update: Cryodamage, markers and factors affecting the sperm freezability in pigs. Theriogenology 2016, 85, 47–64. [Google Scholar] [CrossRef] [PubMed]
- Fraser, L.; Strzeżek, J. Effect of different procedures of ejaculate collection, extenders and packages on DNA integrity of boar spermatozoa following freezing-thawing. Anim. Reprod. Sci. 2007, 99, 317–329. [Google Scholar] [CrossRef]
- Basioura, A.; Tsakmakidis, I.A.; Martinez, E.A.; Roca, J.; Molina, M.F.; Theodoridis, A.; Boscos, C.M.; Parilla, I. Effect of astaxanthin in extenders on sperm quality and functional variables of frozen-thawed boar semen. Anim. Reprod. Sci. 2020, 218, 106478. [Google Scholar] [CrossRef]
- Winn, E.; Whitaker, B.D. Quercetin supplementation to the thawing and incubation media of boar sperm improves post-thaw sperm characteristics and the in vitro production of pig embryos. Reprod. Biol. 2020, 20, 315–320. [Google Scholar] [CrossRef]
- Cappallo-Obermann, H.; Schulze, W.; Jastrow, H.; Baukloh, V.; Spiess, A.N. Highly purified spermatozoal RNA obtained by a novel method indicates an unusual 28S/18S rRNA ratio and suggests impaired ribosome assembly. Mol. Hum. Reprod. 2011, 17, 669–678. [Google Scholar] [CrossRef]
- Hamatani, T. Human spermatozoal RNAs. Fertil. Steril. 2012, 97, 275–281. [Google Scholar] [CrossRef]
- Card, C.J.; Anderson, E.J.; Zamberlan, S.; Krieger, K.E.; Kaproth, M.; Sartini, B.L. Cryopreserved bovine spermatozoal transcript profile as revealed by high-throughput ribonucleic acid sequencing. Biol. Reprod. 2013, 88, 49. [Google Scholar] [CrossRef]
- Jodar, M.; Selvaraju, S.; Sendler, E.; Diamond, M.P.; Krawetz, S.A. The presence, role and clinical use of spermatozoal RNAs. Hum. Reprod. Update 2013, 19, 604–624. [Google Scholar] [CrossRef]
- Valcarce, D.G.; Cartón-García, F.; Herráez, M.P.; Robles, V. Effect of cryopreservation on human sperm messenger RNAs crucial for fertilization and early embryo development. Cryobiology 2013, 67, 84–90. [Google Scholar] [CrossRef]
- Zeng, C.; Peng, W.; Ding, L.; He, L.; Zhang, Y.; Fang, D.; Tang, K. A preliminary study on epigenetic changes during boar spermatozoa cryopreservation. Cryobiology 2014, 69, 119–127. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Y.; Zhu, H.; Hao, H.; Zhao, X.; Qin, T.; Wang, D. Comparative transcript profiling of gene expression of fresh and frozen-thawed bull sperm. Theriogenology 2015, 83, 504–511. [Google Scholar] [CrossRef] [PubMed]
- Yathish, H.M.; Kumar, S.; Dubey, P.P.; Modi, R.P.; Chaudhary, R.; Kumar, A.S.; Ghosh, S.K.; Sarkar, M.; Sivamani, B. Profiling of sperm gene transcripts in crossbred (Bos taurus × Bos indicus) bulls. Anim. Reprod. Sci. 2017, 17, 2534. [Google Scholar] [CrossRef]
- Qin, Z.; Wang, W.; Ali, M.A.; Wang, Y.; Zhang, Y.; Zhang, M.; Zhou, G.; Yang, J.D.; Zeng, C. Transcriptome-wide m6A profiling reveals mRNA post-transcriptional modification of boar sperm during cryopreservation. BMC Genomics 2021, 22, 588. [Google Scholar] [CrossRef] [PubMed]
- Held, T.; Paprotta, I.; Khulan, J.; Hemmerlein, B.; Binder, L.; Wolf, S.; Schubert, S.; Meinhardt, A.; Engel, W.; Adham, I.M. Hspa4l-deficient mice display increased incidence of male infertility and hydronephrosis development. Mol. Cell. Biol. 2006, 26, 8099–8108. [Google Scholar] [CrossRef] [PubMed]
- Held, T.; Barakat, A.Z.; Mohamed, B.A.; Paprotta, I.; Meinhardt, A.; Engel, W.; Adham, I.M. Heat-shock protein HSPA4 is required for progression of spermatogenesis. Reproduction 2011, 142, 133–144. [Google Scholar] [CrossRef]
- Thundathil, J.C.; Anzar, M.; Buhr, M.M. Na/K-ATPase as a signaling molecule during bovine sperm capacitation. Biol. Reprod. 2006, 75, 308–317. [Google Scholar] [CrossRef]
- O’Flaherty, C. The enzymatic antioxidant system of human spermatozoa. Adv. Androl. 2014, 2014, 6263374. [Google Scholar] [CrossRef]
- Liu, X.X.; Shen, X.F.; Liu, F.J. Screening targeted testis-specific genes for molecular assessment of aberrant sperm quality. Mol. Med. Rep. 2016, 14, 1594–1600. [Google Scholar] [CrossRef]
- Liu, X.; Wang, X.; Liu, F. Decreased expression of heat shock protein A4L in spermatozoa is positively related to poor human sperm quality. Mol. Reprod. Dev. 2019, 86, 379–386. [Google Scholar] [CrossRef]
- Fraser, L.; Brym, P.; Pareek, C.S.; Mogielnicka-Brzozowska, M.; Paukszto, Ł.; Jastrzębski, J.P.; Wasilewska-Sakowska, K.; Mańkowska, A.; Sobiech, P.; Żukowski, K. Transcriptome analysis of boar spermatozoa with different freezability using RNA-Seq. Theriogenology 2020, 142, 400–413. [Google Scholar] [CrossRef] [PubMed]
- Corral-Vazquez, C.; Blanco, J.; Aiese Cigliano, R.; Sarrate, Z.; Rivera-Egea, R.; Vidal, F.; Garrido, N.; Daub, C.; Anton, E. The RNA content of human sperm reflects prior events in spermatogenesis and potential post-fertilization effects. Mol. Hum. Reprod. 2021, 27, gaab035. [Google Scholar] [CrossRef] [PubMed]
- Ureña, I.; González, C.; Ramón, M.; Gòdia, M.; Clop, A.; Calvo, J.H.; Carabaño, M.J.; Serrano, M. Exploring the ovine sperm transcriptome by RNAseq techniques. I Effect of seasonal conditions on transcripts abundance. PLoS ONE 2022, 17, e0264978. [Google Scholar] [CrossRef] [PubMed]
- Arnér, E.S.; Holmgren, A. Physiological functions of thioredoxin and thioredoxin reductase. Eur. J. Biochem. 2000, 267, 6102–6109. [Google Scholar] [CrossRef] [PubMed]
- Nordberg, J.; Arnér, E.S. Reactive oxygen species, antioxidants, and the mammalian thioredoxin system. Free Rad. Biol. Med. 2001, 31, 1287–1312. [Google Scholar] [CrossRef]
- Moradi, M.N.; Karimi, J.; Khodadadi, I.; Amiri, I.; Karami, M.; Saidijam, M.; Vatannejad, A.; Tavilani, H. Evaluation of the p53 and Thioredoxin reductase in sperm from asthenozoospermic males in comparison to normozoospermic males. Free Rad. Biol. Med. 2018, 116, 123–128. [Google Scholar] [CrossRef]
- Kalmar, B.; Greensmith, L. Induction of heat shock proteins for protection against oxidative stress. Adv. Drug Deliv. Rev. 2009, 1, 310–318. [Google Scholar] [CrossRef]
- Blanco, G.; Mercer, R.W. Isozymes of the Na-K-ATPase: Heterogeneity in structure, diversity in function. Am. J. Physiol. 1998, 275, 633–650. [Google Scholar] [CrossRef]
- Thundathil, J.C.; Rajamanickam, G.D.; Kastelic, J.P. Na/K-ATPase and regulation of sperm function. Anim. Reprod. 2018, 15, 711–720. [Google Scholar] [CrossRef]
- Ryu, D.Y.; Song, W.H.; Pang, W.K.; Yoon, S.J.; Rahman, M.S.; Pang, M.G. Freezability biomarkers in bull epididymal spermatozoa. Sci. Rep. 2019, 9, 12797. [Google Scholar] [CrossRef]
- Salinas, M.B.S.; Lertwichaikul, T.; Khunkaew, C.; Boonyayatra, S.; Sringarm, K.; Chuammitri, P.; Sathanawongs, A. Freezability biomarkers in the epididymal spermatozoa of swamp buffalo. Cryobiology 2022, 106, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Chen, X.; Wang, Z.; Wang, D. Is transcription in sperm stationary or dynamic? J. Reprod. Dev. 2017, 63, 439–443. [Google Scholar] [CrossRef] [PubMed]
- Riesco, M.F.; Robles, V. Cryopreservation causes genetic and epigenetic changes in zebrafish genital ridges. PLoS ONE 2013, 8, e67614. [Google Scholar] [CrossRef]
- Card, C.J.; Kreiger, K.E.; Kaproth, M.; Sartini, B.L. Oligo-dT selected spermatozoal transcript profiles differ among higher and lower fertility dairy sires. Anim. Reprod. Sci. 2017, 177, 105–123. [Google Scholar] [CrossRef] [PubMed]
- Dai, D.H.; Qazi, I.H.; Ran, M.X.; Liang, K.; Zhang, Y.; Zhang, M.; Zhou, G.B.; Angel, C.; Zeng, C.J. Exploration of miRNA and mRNA profiles in fresh and frozen-thawed boar sperm by transcriptome and small RNA Sequencing. Int. J. Mol. Sci. 2019, 20, 802. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Rodriguez, J.M.; Ortega-Ferrusola, C.; Gil, M.C.; Martín-Cano, F.E.; Gaitskell-Phillips, G.; Rodríguez-Martínez, H.; Hinrichs, K.; Álvarez-Barrientos, A.; Roman, Á.; Peña, F.J. Transcriptome analysis reveals that fertilization with cryopreserved sperm downregulates genes relevant for early embryo development in the horse. PLoS ONE 2019, 14, e0213420. [Google Scholar] [CrossRef]
- Maylem, E.R.S.; Rivera, S.M.; Ramos, G.E.; Atabay, E.C.; Venturina, E.V.; Atabay, E.P. Changes on the heat shock protein 70 (HSP70) in water buffalo spermatozoa revealed the capacitation like event in cryopreservation. Asian J. Agric. & Biol. 2021, 3, 1–7. [Google Scholar] [CrossRef]
- Chen, X.; Zhu, H.; Hu, C.; Hao, H.; Zhang, J.; Li, K.; Zhao, X.; Qin, T.; Zhao, K.; Zhu, H.; et al. Identification of differentially expressed proteins in fresh and frozen–thawed boar spermatozoa by iTRAQ-coupled 2D LC–MS/MS. Reproduction 2014, 147, 321–330. [Google Scholar] [CrossRef]
- Yoon, S.J.; Rahman, M.S.; Kwon, W.S.; Ryu, D.Y.; Park, Y.J.; Pang, M.G. Proteomic identification of cryostress in epididymal spermatozoa. J. Anim. Sci. 2016, 7, 67. [Google Scholar] [CrossRef]
- Davila, M.P.; Muñoz, P.M.; Bolaños, J.M.; Stout, T.A.; Gadella, B.M.; Tapia, J.A.; da Silva, C.B.; Ferrusola, C.O.; Peña, F.J. Mitochondrial ATP is required for the maintenance of membrane integrity in stallion spermatozoa, whereas motility requires both glycolysis and oxidative phosphorylation. Reproduction 2016, 152, 683–694. [Google Scholar] [CrossRef]
- Gur, Y.; Breitbart, H. Protein synthesis in sperm: Dialog between mitochondria and cytoplasm. Mol. Cell Endocrinol. 2008, 282, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Vogel, C.; Marcotte, E. Insights into the regulation of protein abundance from proteomic and transcriptomic analyses. Nat. Rev. Genet. 2012, 13, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wang, W.; Xu, Y.; Tang, M.; Fang, J.; Sun, H. Proteomic characteristics of human sperm cryopreservation. Proteomics 2014, 14, 298–310. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J. Protein degradation and phosphorylation after freeze thawing result in spermatozoon dysfunction. Proteomics 2014, 14, 155–156. [Google Scholar] [CrossRef]
- Bogle, O.A.; Kumar, K.; Attardo-Parrinello, C.; Lewis, S.E.M.; Estanyol, J.M.; Ballesc, J.L.; Oliva, R. Identification of protein changes in human spermatozoa throughout the cryopreservation process. Andrology 2017, 5, 10–22. [Google Scholar] [CrossRef]
- Jia, B.; Larbi, A.; Lv, C.; Liang, J.; Xiang, D.; Zhang, B.; Fang, Y.; Shen, W.; Wu, G.; Quan, G. Identification and validation of ram sperm proteins associated with cryoinjuries caused by cryopreservation process. Theriogenology 2022, 184, 191–203. [Google Scholar] [CrossRef]
- Stephanou, A.; Latchman, D.D. Transcriptional modulation of heat-shock protein gene. Biochem. Res. 2011, 2011, 238601. [Google Scholar] [CrossRef]
- Corcini, C.D.; Varela, A.S., Jr.; Pigozzo, R.; Rambo, G.; Goularte, K.L.; Calderam, K.; Leon, P.M.M.; Bongalhardo, D.C.; Lucia, T., Jr. Pre-freezing and post-thawing quality of boar sperm for distinct portions of the ejaculate and as a function of protein bands present in seminal plasma. Livest. Sci. 2012, 145, 28–33. [Google Scholar] [CrossRef]
- Rego, J.P.; Martins, J.M.; Wolf, C.A.; van Tilburg, M.; Moreno, F.; Monteiro-Moreira, A.C.; Moreira, R.A.; Santos, D.O.; Moura, A.A. Proteomic analysis of seminal plasma and sperm cells and their associations with semen freezability in Guzerat bulls. J. Anim. Sci. 2016, 12, 5308–5320. [Google Scholar] [CrossRef]
- Michos, I.; Tsantarliotou, M.; Boscos, C.M.; Tsousis, G.; Basioura, A.; Tzika, E.D.; Tassis, P.D.; Lymberopoulos, A.G.; Tsakmakidis, I.A. Effect of boar sperm proteins and quality changes on field fertility. Animals 2021, 11, 1813. [Google Scholar] [CrossRef]
- Fraser, L.; Parda, A.; Filipowicz, K.; Strzeżek, J. Comparison of post-thaw DNA integrity of boar spermatozoa assessed with the neutral Comet assay and sperm-Sus Halomax test kit. Reprod. Domest. Anim. 2010, 45, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Wasilewska-Sakowska, K.; Zasiadczyk, Ł.; Fraser, L. Effect of fractionated seminal plasma on sperm characteristics following cryopreservation of boar semen. Ann. Anim. Sci. 2019, 19, 695–712. [Google Scholar] [CrossRef]
- Fraser, L.; Strzeżek, J.; Kordan, W. Post-thaw sperm characteristics following long-term storage of boar semen in liquid nitrogen. Anim. Reprod. Sci. 2014, 147, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Garner, D.L.; Johnson, L.A. Viability assessment of mammalian sperm using SYBR-14 and propidium iodide. Biol. Reprod. 1995, 53, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Fraser, L.; Dziekońska, A.; Strzeżek, R.; Strzeżek, J. Dialysis of boar semen prior to freezing thawing: Its effects on post-thaw sperm characteristics. Theriogenology 2007, 67, 994–1003. [Google Scholar] [CrossRef]
- Wasilewska, K.; Fraser, L. Boar variability in sperm cryo-tolerance after cooling of semen in different long-term extenders at various temperatures. Anim. Reprod. Sci. 2017, 185, 161–173. [Google Scholar] [CrossRef]
- Koziorowska-Gilun, M.; Fraser, L.; Gilun, P.; Koziorowski, M.; Kordan, W. Activity of antioxidant enzymes and their mRNA expression in reproductive tract tissues of the male roe deer (Capreolus capreolus) during pre-rut and rut season. Small Rumin. Res. 2015, 129, 97–103. [Google Scholar] [CrossRef]
- Zeng, C.; He, L.; Peng, W.; Ding, L.; Tang, K.; Fang, D.; Zhang, Y. Selection of optimal reference genes for quantitative RT-PCR studies of boar spermatozoa cryopreservation. Cryobiology 2014, 68, 113–121. [Google Scholar] [CrossRef]
- Zhao, S.; Fernald, R. Comprehensive algorithm for quantitative real-time polymerase chain reaction. J. Comput. Biol. 2005, 12, 1045–1062. [Google Scholar] [CrossRef]
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. g:Profiler: A web server for functional enrichment analysis and conversions of gene lists (2019 update). Nucleic Acids Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Towbin, H.; Staehelin, T.; Gordon, J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. USA 1979, 76, 4350–4354. [Google Scholar] [CrossRef]
- Vilagran, I.; Castillo, J.; Bonet, S.; Sancho, S.; Yeste, M.; Estanyol, J.M.; Oliva, R. Acrosin-binding protein (ACRBP) and triosephosphate isomerase (TPI) are good markers to predict boar sperm freezing capacity. Theriogenology 2013, 80, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING database in 2021: Customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef] [PubMed]
- Blanch, E.; Tomás, C.; Graham, J.K.; Mocé, E. Response of boar sperm to the treatment with cholesterol-loaded cyclodextrins added prior to cryopreservation. Reprod. Domest. Anim. 2012, 47, 959–964. [Google Scholar] [CrossRef] [PubMed]
- Mańkowska, A.; Brym, P.; Sobiech, P.; Fraser, L. Promoter polymorphisms in STK35 and IFT27 genes and their associations with boar sperm freezability. Theriogenology 2022, 189, 199–208. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sperm Parameters | GSF (n = 25) | PSF (n = 25) | p-Value |
|---|---|---|---|
| Total motility (TMOT, %) | 89.39 ± 0.69 | 87.38 ± 0.92 | 0.846 |
| Progressive motility (PMOT, %) | 63.84 ± 1.24 | 65.83 ± 1.06 | 0.231 |
| Velocity straight line (VSL, μm/s) | 75.90 ± 2.10 | 77.18 ± 2.01 | 0.584 |
| Velocity average path (VAP, μm/s) | 91.90 ± 1.76 | 93.58 ± 2.46 | 0.767 |
| Velocity curvilinear (VCL, μm/s) | 127.18 ± 4.20 | 125.18 ± 4.20 | 0.882 |
| Straightness (STR, %) | 81.35 ± 1.85 | 82.73 ± 1.29 | 0.535 |
| Linearity (LIN, %) | 60.32 ± 1.35 | 62.16 ± 1.45 | 0.355 |
| Amplitude of lateral head displacement (ALH, μm) | 7.16 ± 0.24 | 7.44 ± 0.19 | 0.657 |
| Beat cross frequency (BCF, Hz) | 30.40 ± 0.72 | 31.83 ± 0.95 | 0.231 |
| Mitochondrial membrane potential (MMP, %) | 87.51 ± 0.56 | 88.14 ± 0.67 | 0.664 |
| Plasma membrane integrity (PMI, %) | 88.17 ± 0.51 | 87.55 ± 1.05 | 0.721 |
| NAR acrosome integrity (%) | 91.88 ± 0.49 | 90.76 ± 0.71 | 0.185 |
| DNA fragmentation (%) | 2.23 ± 0.30 | 2.38 ± 0.26 | 0.698 |
| Sperm Parameters | GSF (n = 25) | PSF (n = 25) | p-Value |
|---|---|---|---|
| Total motility (TMOT, %) | 51.29 ± 1.47 a | 24.66 ± 0.82 b | 0.001 |
| Progressive motility (PMOT, %) | 40.71 ± 1.28 a | 17.17 ± 1.35 b | 0.001 |
| Velocity straight line (VSL, μm/s) | 60.97 ± 2.39 a | 46.15 ± 3.02 b | 0.001 |
| Velocity average path (VAP, μm/s) | 76.84 ± 2.56 a | 60.48 ± 3.16 b | 0.001 |
| Velocity curvilinear (VCL, μm/s) | 133.12 ± 3.50 a | 112.18 ± 2.79 b | 0.001 |
| Straightness (STR, %) | 79.10 ± 1.22 a | 75.20 ± 1.43 b | 0.042 |
| Linearity (LIN, %) | 46.07 ± 1.56 a | 40.75 ± 1.18 b | 0.049 |
| Amplitude of lateral head displacement (ALH, μm) | 5.49 ± 0.27 a | 6.42 ± 1.09 a | 0.375 |
| Beat cross frequency (BCF, Hz) | 21.70 ± 0.96 a | 28.23 ± 3.12 a | 0.072 |
| Mitochondrial membrane potential (MMP, %) | 52.10 ± 1.24 a | 31.61 ± 1.64 b | 0.001 |
| Plasma membrane integrity (PMI, %) | 51.24 ± 1.09 a | 36.46 ± 1.26 b | 0.001 |
| NAR acrosome integrity (%) | 51.45 ± 1.34 a | 38.50 ± 1.58 b | 0.001 |
| DNA fragmentation (%) | 7.05 ± 0.48 a | 12.90 ± 0.67 b | 0.001 |
| Gene | Ensemble Accession No. | Primer Sequence | Start (bp) | Stop (bp) | Amplicon Size (bp) |
|---|---|---|---|---|---|
| TXNRD1 | NM_214154.3 | F: 5′-CCAAACCCAAGGCGAAGTTT-3′ R: 5′-GTGTAAGCACGGGACACGC-3′ | 1589 1654 | 1608 1636 | 65 |
| HSPA4L | XM_021101609.1 | F: 5′-GGAGGTTGCGGCGCAG-3′ R: 5′-CTAGGGCAGTGCGTTGGG-3′ | 35 93 | 50 76 | 58 |
| ATP1B1 | NM_001001542.1 | F: 5′-CCATCTTCAATCCCCGCA-3′ R: 5′-GCTTTTCCGCGGGCC-3′ | 89 143 | 106 129 | 54 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mańkowska, A.; Gilun, P.; Zasiadczyk, Ł.; Sobiech, P.; Fraser, L. Expression of TXNRD1, HSPA4L and ATP1B1 Genes Associated with the Freezability of Boar Sperm. Int. J. Mol. Sci. 2022, 23, 9320. https://doi.org/10.3390/ijms23169320
Mańkowska A, Gilun P, Zasiadczyk Ł, Sobiech P, Fraser L. Expression of TXNRD1, HSPA4L and ATP1B1 Genes Associated with the Freezability of Boar Sperm. International Journal of Molecular Sciences. 2022; 23(16):9320. https://doi.org/10.3390/ijms23169320
Chicago/Turabian StyleMańkowska, Anna, Przemysław Gilun, Łukasz Zasiadczyk, Przemysław Sobiech, and Leyland Fraser. 2022. "Expression of TXNRD1, HSPA4L and ATP1B1 Genes Associated with the Freezability of Boar Sperm" International Journal of Molecular Sciences 23, no. 16: 9320. https://doi.org/10.3390/ijms23169320
APA StyleMańkowska, A., Gilun, P., Zasiadczyk, Ł., Sobiech, P., & Fraser, L. (2022). Expression of TXNRD1, HSPA4L and ATP1B1 Genes Associated with the Freezability of Boar Sperm. International Journal of Molecular Sciences, 23(16), 9320. https://doi.org/10.3390/ijms23169320