
Cyclophilin A/CD147 Interaction: A Promising Target for Anticancer Therapy


Abstract
:1. Introduction
2. Biological Functions of CypA and CD147
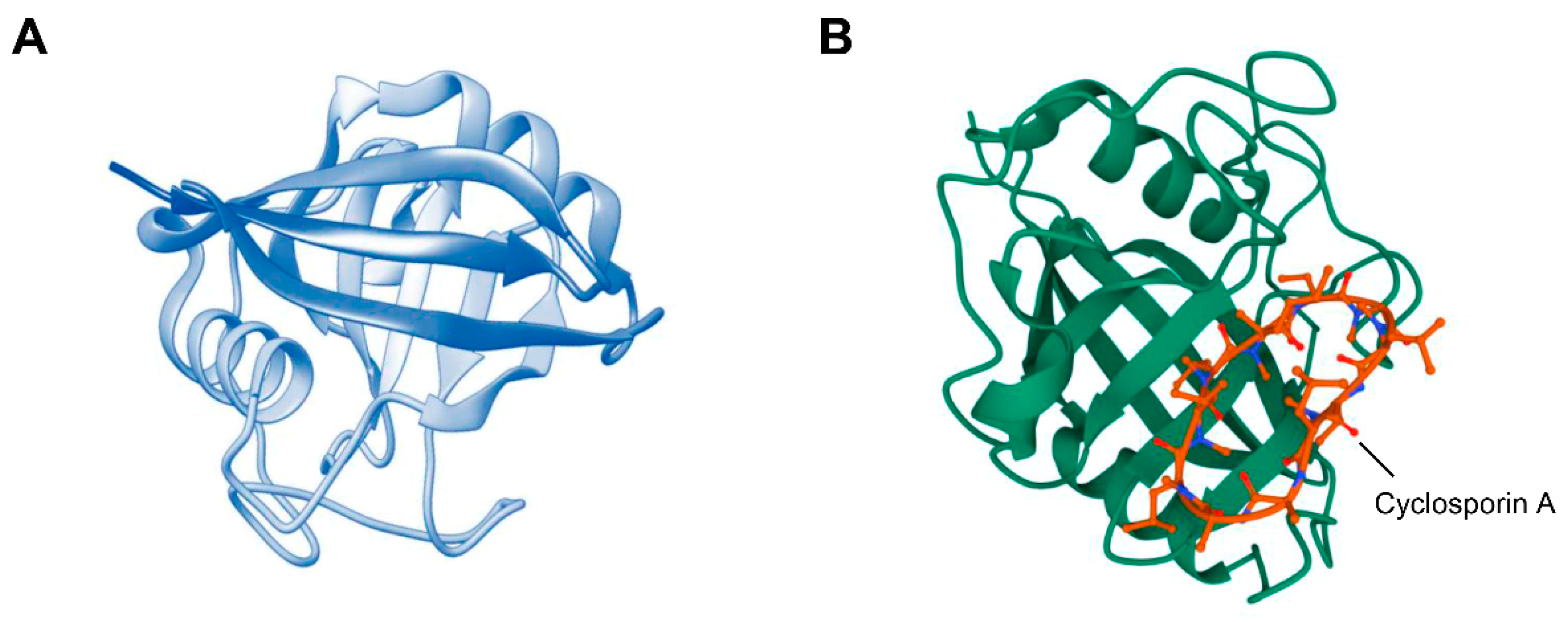
2.1. Biology of CypA
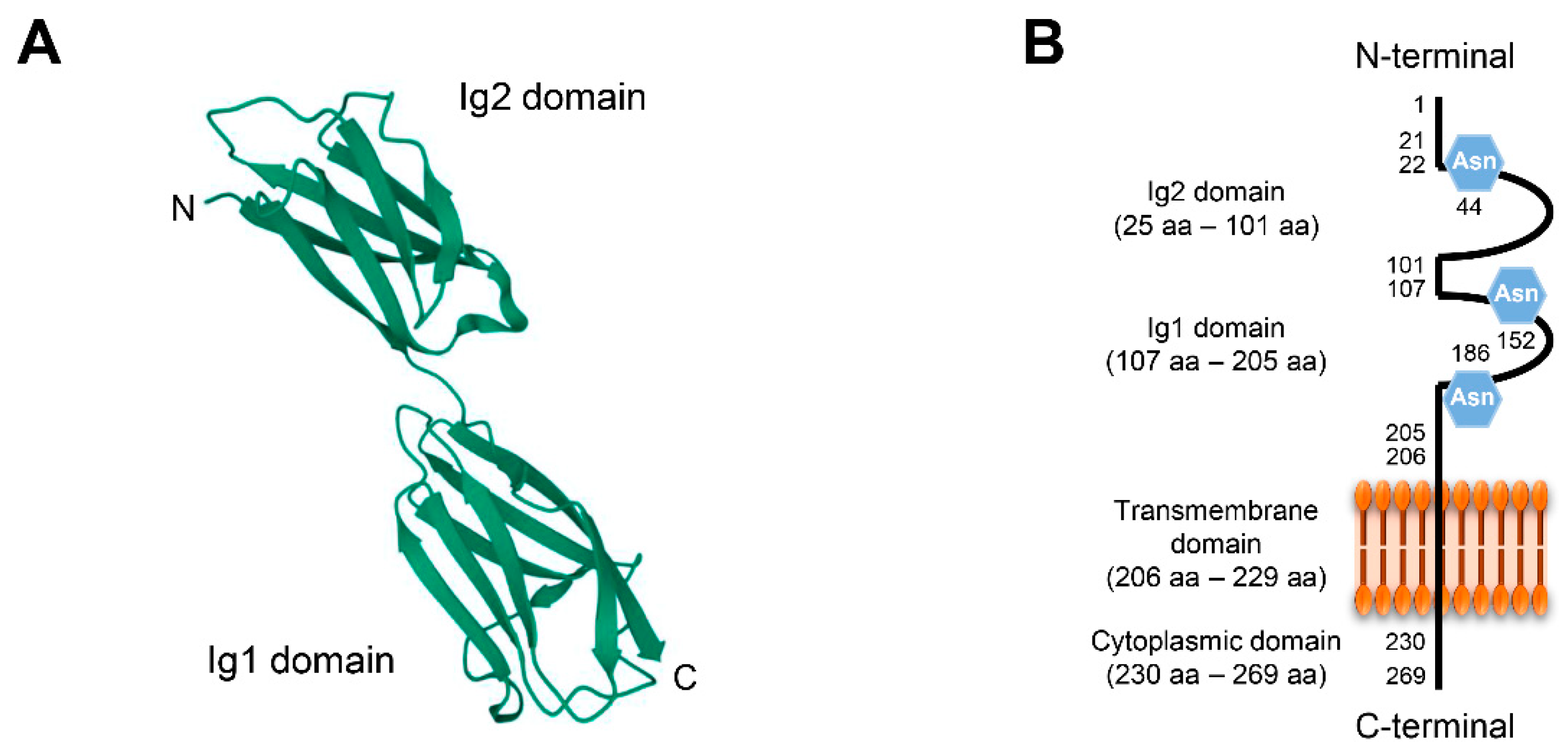
2.2. Biology of CD147
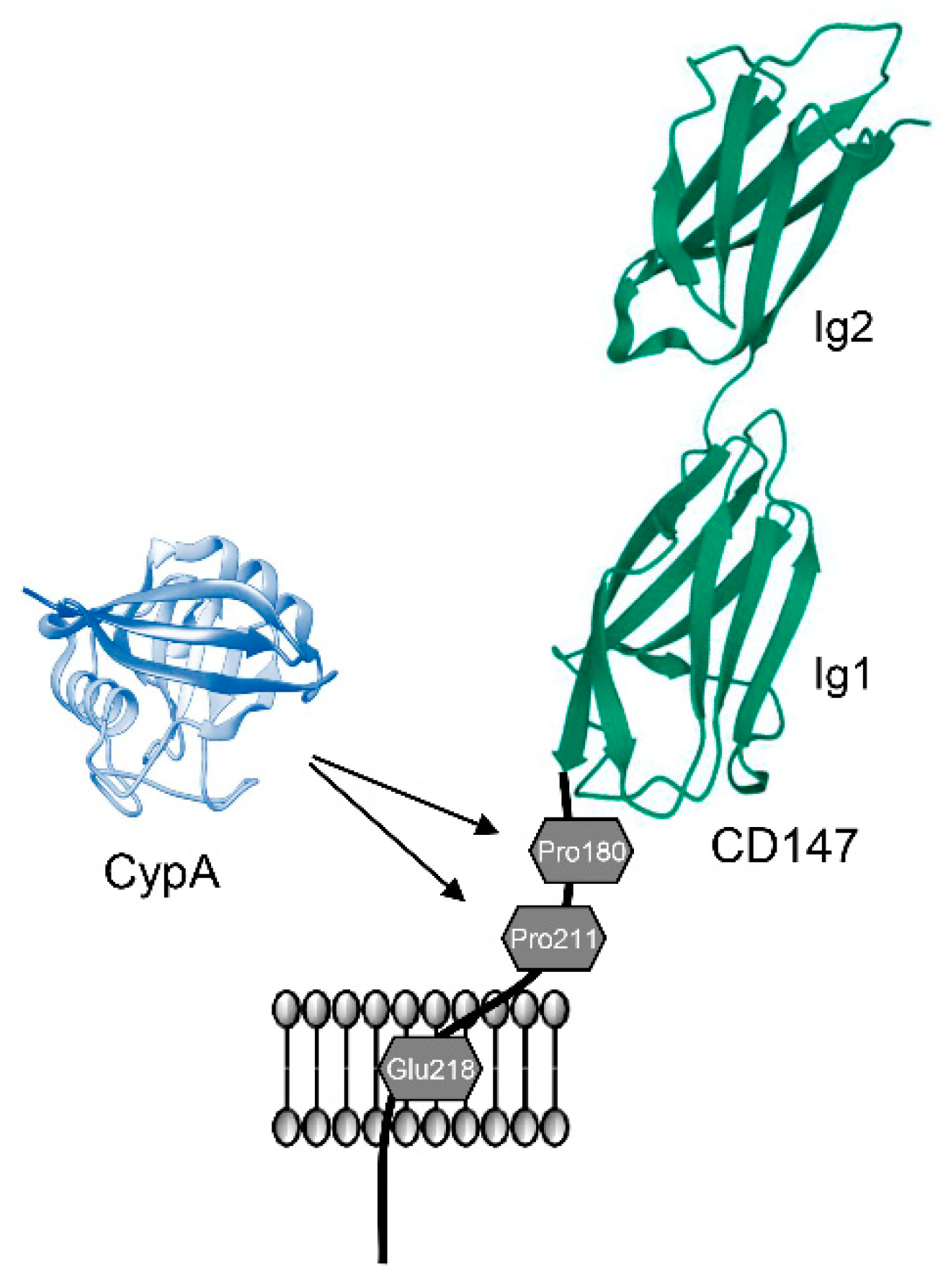
2.3. Interaction between CypA and CD147
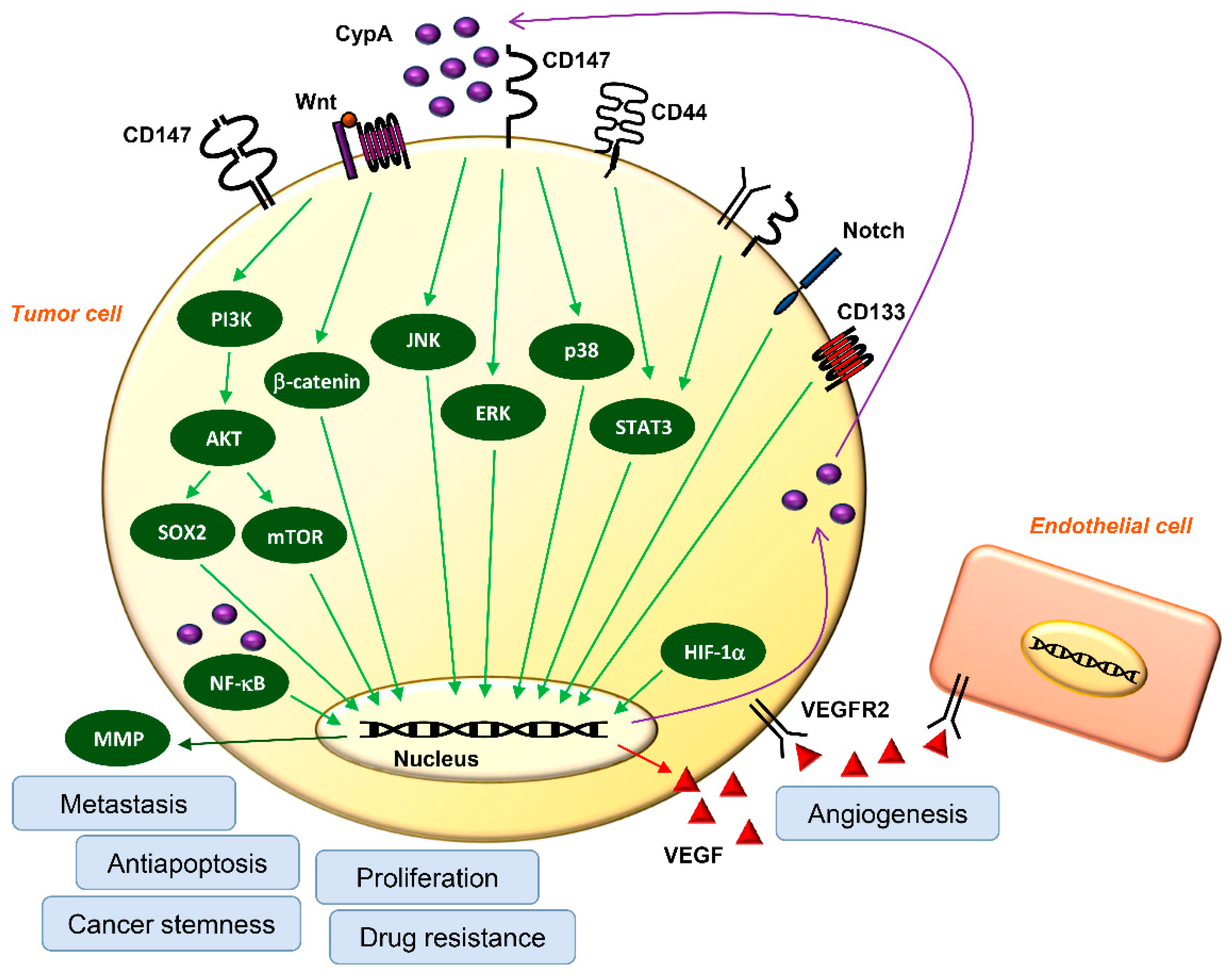
3. Roles of CypA/CD147 Interaction in Cancer
3.1. Proliferation
3.2. Metastasis
3.3. Antiapoptosis
3.4. Resistance to Chemotherapy and Radiation Therapy
3.5. Cancer Stem Cells (CSCs)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| The Key Processes of Cancer Progression | Protein | The Roles of CypA and CD147 in Cancer | References |
|---|---|---|---|
| Proliferation | CypA | Overexpression and autocrine secretion by activation of HIF-1α | [36,56] |
| IL-8 mediated proliferation by stabilization and nuclear accumulation of NF-κB p65 | [34,61] | ||
| Promotion of the cell cycle transition from G1 to S phase | [62] | ||
| Causes of higher tumor malignancy | [62] | ||
| Activation of JAK2/STAT5 signaling pathway | [65] | ||
| CD147 | Promotion of tumorigenesis with CD44 | [66] | |
| Induction of Ki-67 expression | [67] | ||
| CypA and CD147 | Activation of JNK/ERK1/2/p38 MAPK signaling pathways | [34,54,58,59,60] | |
| Overexpression in human tumor tissues than in normal tissues | [58] | ||
| Induction of the secretion of IL-5 and IL-17 | [58] | ||
| Activation of PI3K/AKT/mTOR signaling pathway | [13,54,59,60] | ||
| Poor prognosis | [63,64,67] | ||
| Stimulation of cyclin D1/survivin by activation of STAT3 | [66] | ||
| Metastasis | CypA | Induction of migration through p38 MAPK activation | [68] |
| Positive correlation with MMP | [69] | ||
| Local recurrence and distant metastasis | [73] | ||
| CD147 | Promotion of metastasis with CD44 | [72] | |
| Induction of angiogenesis by increasing VEGF production | [74] | ||
| CypA and CD147 | Promotion of the invasion and migration by induction of MMP-2/MMP-9 expression | [13,69,70,71] | |
| Antiapoptosis | CypA | Inhibition of hypoxia- and cisplatin-induced apoptosis | [56] |
| Upregulation after radiation therapy | [80] | ||
| CD147 | Promotion of anticancer efficacy by trastuzumab | [81] | |
| CypA and CD147 | Activation of PI3K/AKT/mTOR signaling pathway | [13,78,79] | |
| Modulation of Bcl-2 family | [13,78,79] | ||
| Inhibition of caspase cascades | [13,78,79] | ||
| Contribution to cancer aggressiveness | [83,84] | ||
| Resistance to chemotherapy and radiation therapy | CypA | Inhibition of cisplatin-induced apoptosis and causes of drug resistance | [56] |
| Upregulation of IL6, MRP2, MGST1, and GSTZ1 by increasing the expression of ABC transporter | [85] | ||
| Causes of chemoresistance through redox modification | [86] | ||
| Causes of resistance to ER stress inducer-caused apoptosis | [64] | ||
| CD147 | Causes of radiation resistance by interacting with integrin β1 | [87] | |
| Promotion of tumor chemotolerance through interactions with CD44 and HA receptor | [70] | ||
| Cancer stem cells | CypA | Promotion of self-renewal, proliferation, and radiotherapy resistance through Wnt/β-catenin signaling | [14] |
| The most stably expressed essential gene in CSC | [89] | ||
| Induction of sphere formation, self-renewal, and metastasis through Notch signaling | [90] | ||
| CD147 | Release of small extracellular vesicles for invasion | [49] | |
| Induction of self-renewal capacity, differentiation, and in vivo tumorigenic potential | [91] | ||
| Activation of SLC34A2/PI3K/AKT/SOX2 signaling | [91] | ||
| CypA and CD147 | Promotion of CD44+/CD133+ CSCs through the activation of PTEN/PI3K/AKT | [88] | |
| Induction of CSC features through STAT3 signaling | [15] |
4. Therapeutic Potential of Targeting CypA/CD147 in Cancer
| Target Protein | Inhibitors | Mechanism | Therapeutic Potential | Cancer | References |
|---|---|---|---|---|---|
| CypA | Cyclosporin A | PPIase activity | Interference of CypA and CD147 binding, Induction of apoptosis | Breast cancer, Lung adenocarcinoma | [20,51,77,82] |
| Sanglifehrin A | PPIase activity | Binding to CypA with about 60-fold higher affinity than CsA | T cells, B cells, Glioblastoma multiforme | [92,93,94] | |
| Debio-025 (Alisporivir) | Crk signaling | Nonimmunosuppressive analogue of CsA, Potent antitumor and antimetastatic activity, Enhancing of tumor immunogenicity and anti-PD-1 therapy | Breast cancer, Hepatitis C-hepatocellular carcinoma | [96] | |
| SCY-635, NIM811 | NS5A | Nonimmunosuppressive activity, Inhibition of hepatocarcinogenesis | Hepatitis C-hepatocellular carcinoma | [97] | |
| NV651 | PPIase activity | Nonimmunosuppressive activity, More potent PPIase activity than CsA and SfA, Inhibition of cell proliferation and tumor growth in vivo | Hepatocellular carcinoma | [98] | |
| 23-demethyl 8,13-deoxynargenicin (compound 9) | MAPK signaling | Inhibition of proliferation, migration, invasion, and angiogenesis | Gastric cancer | [13,83,99] | |
| Melittin | MMP-9 | Inhibition of Metastasis | Breast cancer | [100] | |
| RNA interference | PPIase activity | Inhibition of tumor growth, Enhancing of radiosensitivity | Lung adenocarcinoma | [54,80] | |
| NF-κB signaling | Inhibition of glioblastoma growth | Glioblastoma | [61] | ||
| CD147 | AC-73 | CD147 dimerization, MAPK/STAT3 signaling, MMP-2 | Inhibition of metastasis and growth, Activation of autophagy, Increase of chemotherapy sensitivity | Hepatocellular carcinoma, Acute myeloid leukemia | [101,102] |
| Metuximab (Licartin) | CD44/STAT3 signaling | Prevention of tumor recurrence, Increase of sensitivity to chemoradiation therapy | Hepatocellular carcinoma, Pancreatic cancer | [48,103,104] | |
| RNA interference | Stemness markers | Suppression of the proliferation and invasion of CSCs, Promotion of chemosensitivity | Colorectal adenocarcinoma | [105] | |
| Caspase-3/9, MAPK pathway, AKT pathway | Promotion of anticancer activity of trastuzumab | HER2-positive breast cancer | [81] | ||
| XIAP | Induction of apoptosis | Oral squamous carcinoma, Multidrug-resistant cancer | [76] | ||
| Caspase-3, MMP-9 | Induction of apoptosis, Inhibition of invasion | Triple-negative breast cancer | [75] |
5. Expression and Clinical Significance of CypA and CD147 in Cancer
6. Conclusions and Future Perspectives
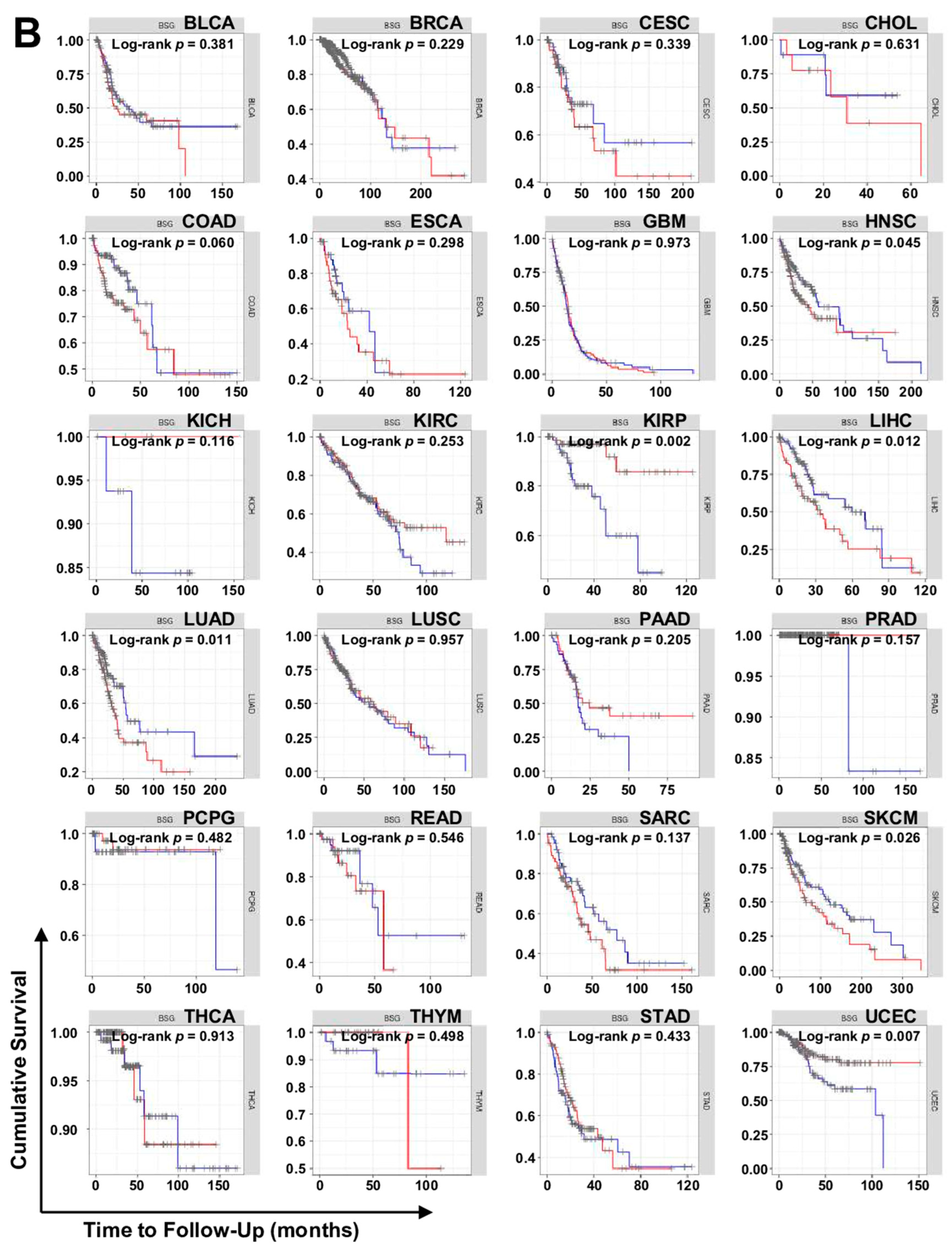
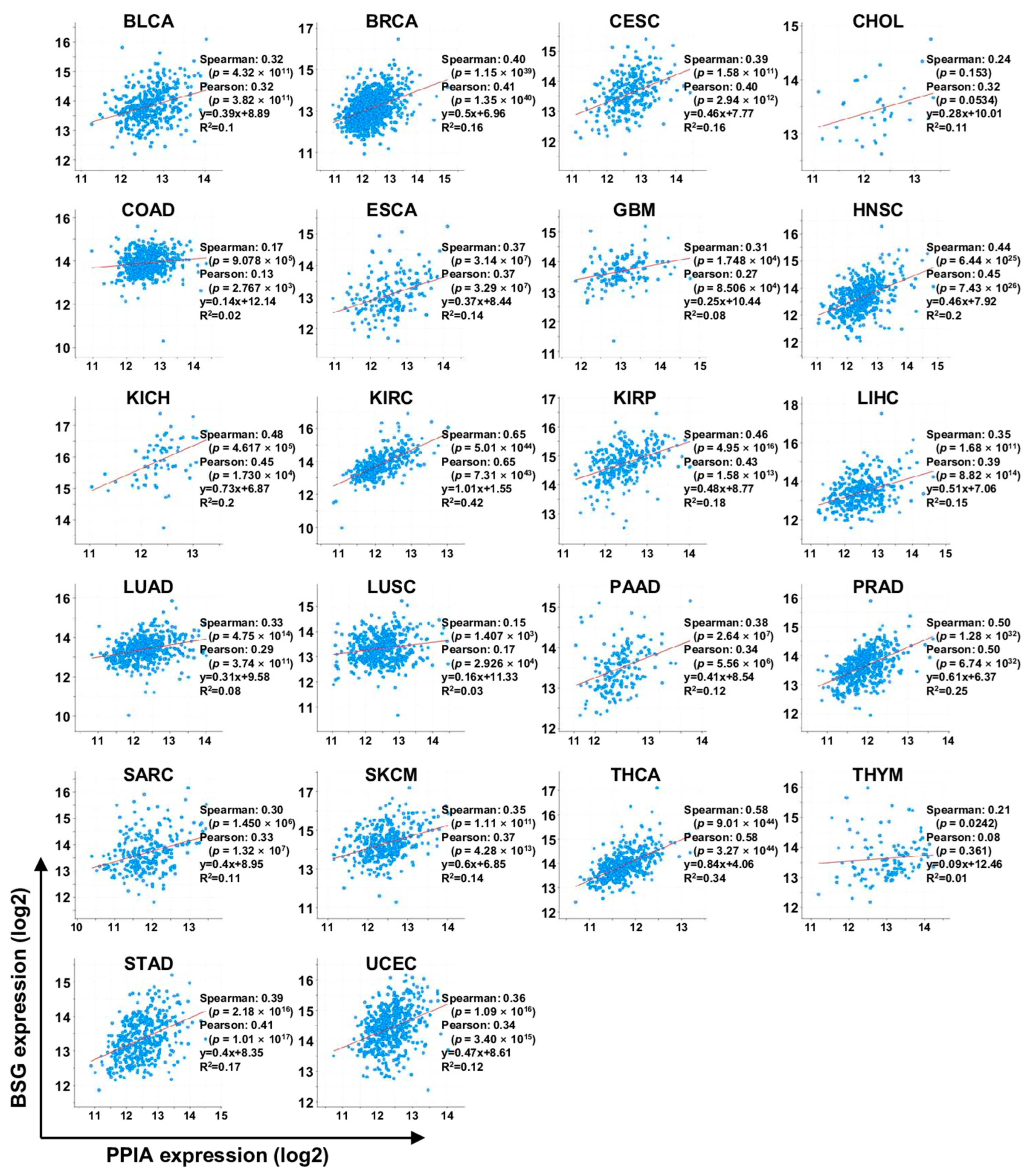
7. TCGA Database Analysis
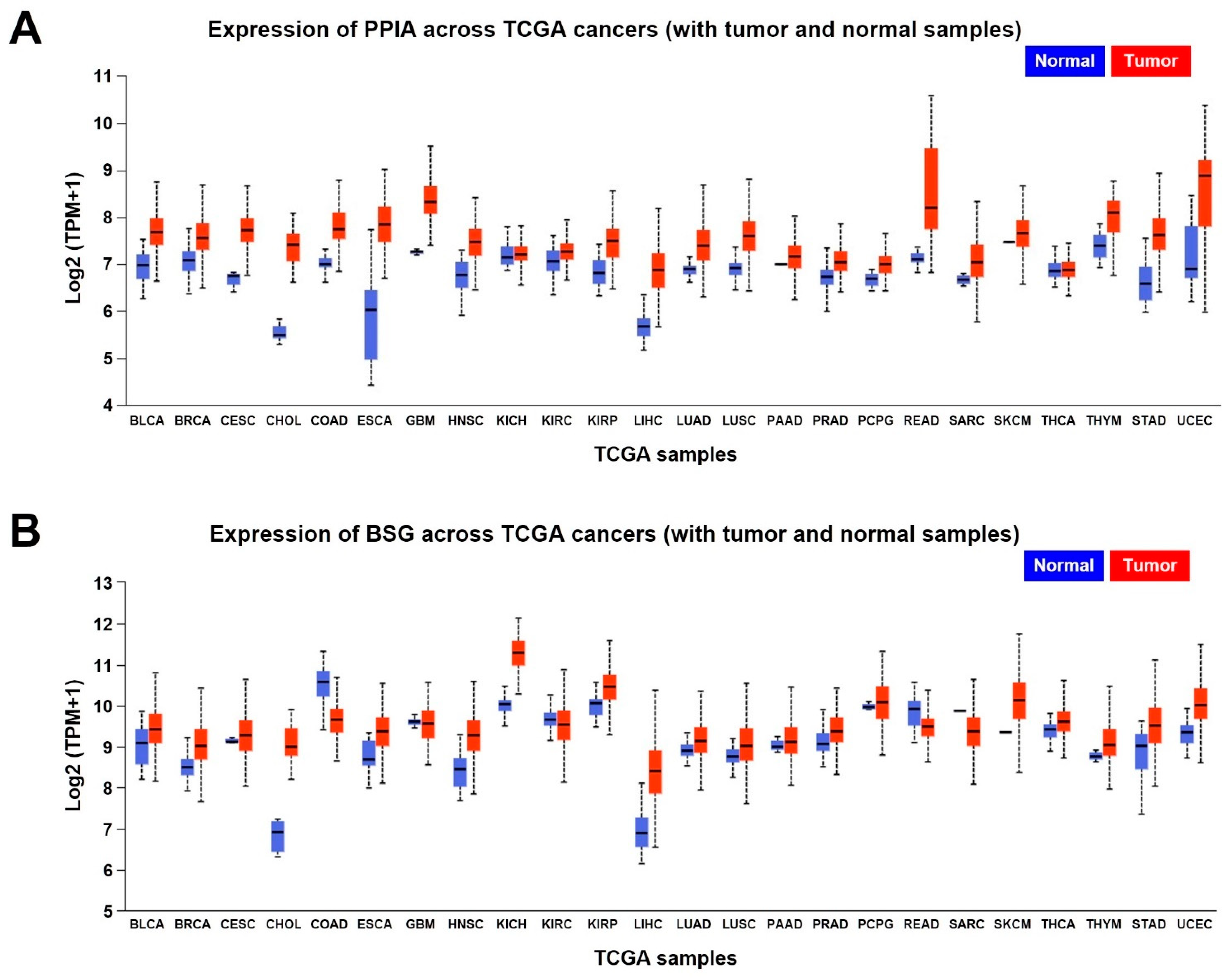
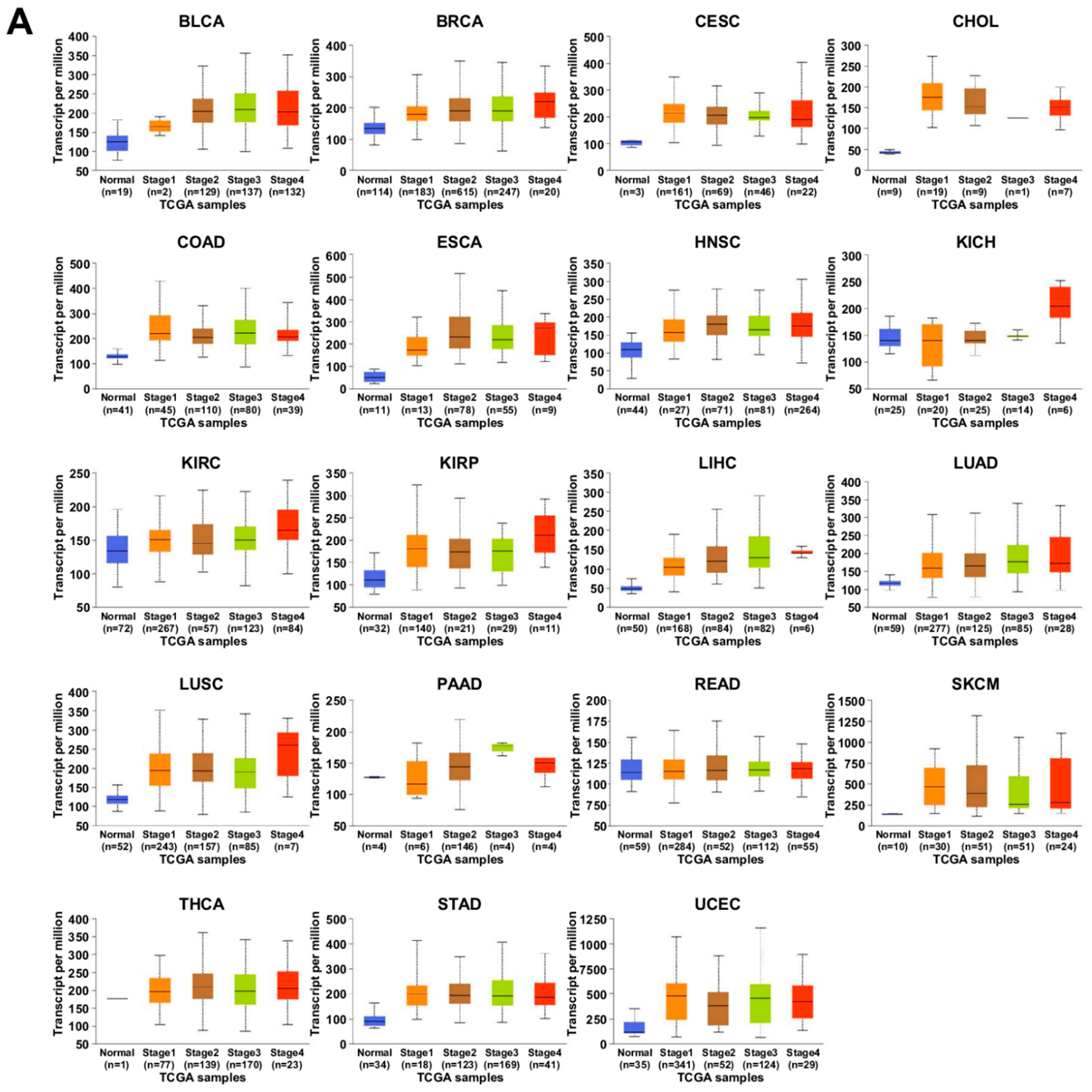
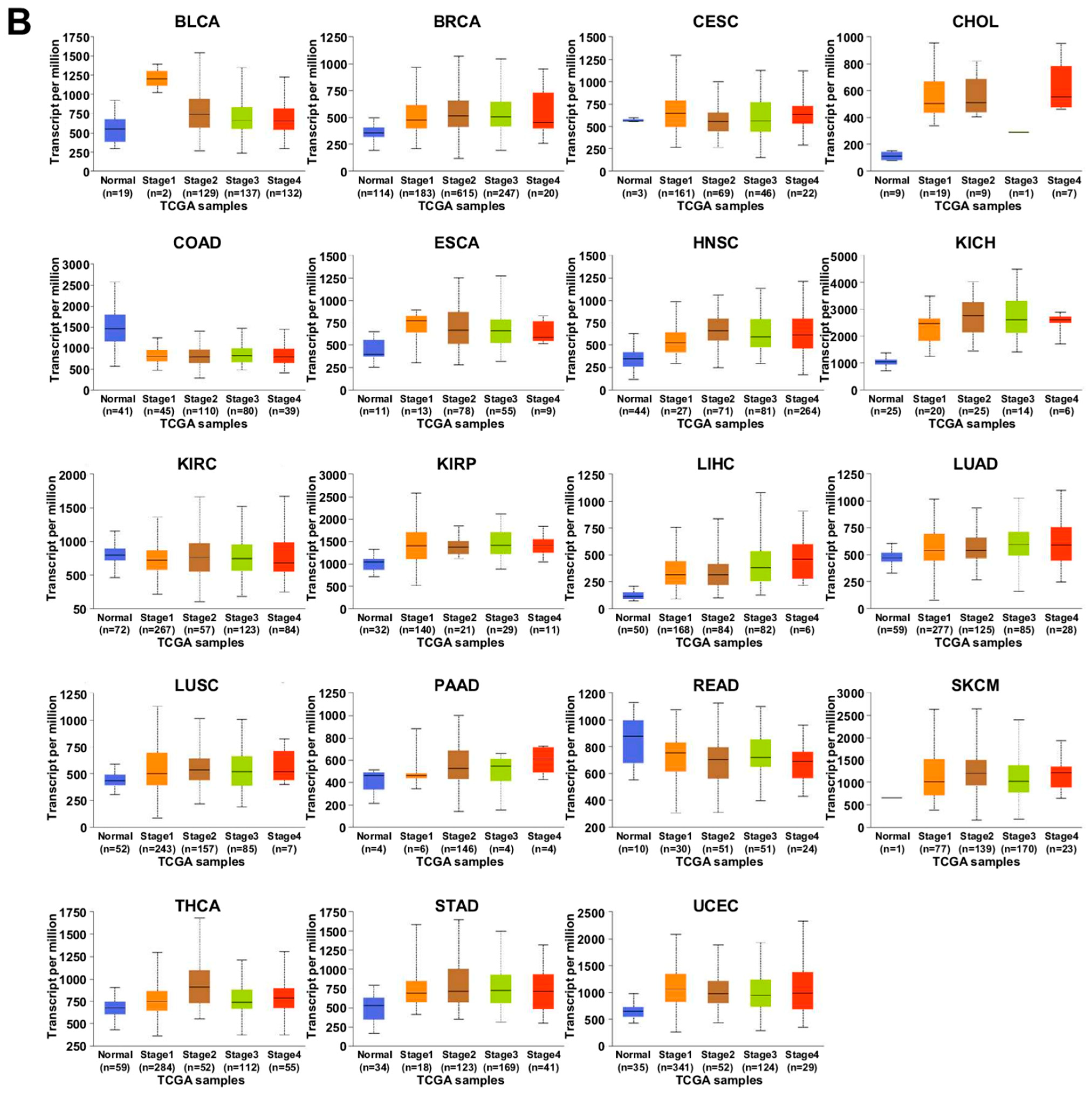
7.1. UALCAN Analysis
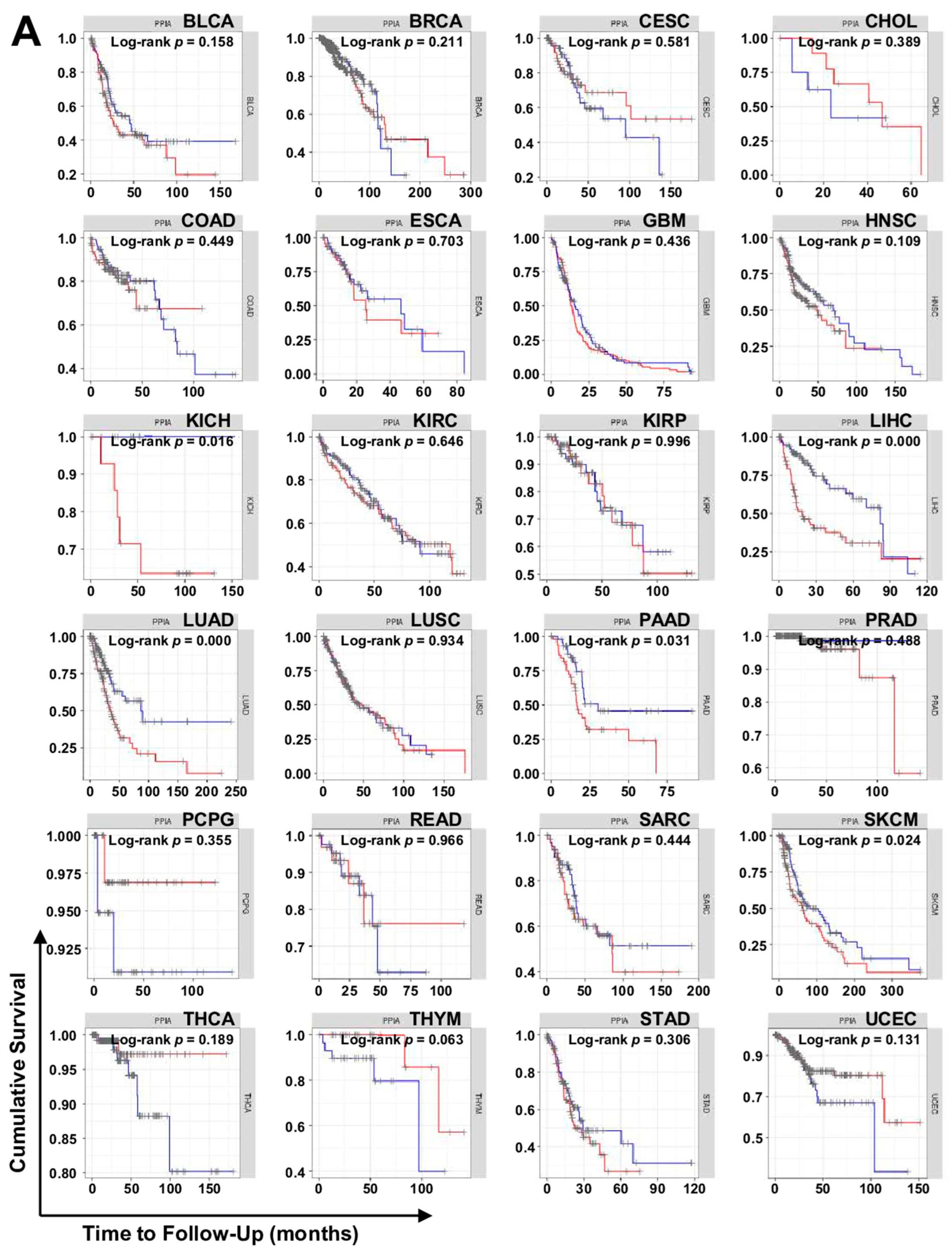
7.2. TIMER Analysis
7.3. cBioPortal Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [PubMed]
- Sun, Y. Translational horizons in the tumor microenvironment: Harnessing breakthroughs and targeting cures. Med. Res. Rev. 2015, 35, 408–436. [Google Scholar] [PubMed]
- Batlle, E.; Clevers, H. Cancer stem cells revisited. Nat. Med. 2017, 23, 1124–1134. [Google Scholar] [PubMed]
- Reya, T.; Morrison, S.J.; Clarke, M.F.; Weissman, I.L. Stem cells, cancer, and cancer stem cells. Nature 2001, 414, 105–111. [Google Scholar]
- Chen, W.; Dong, J.; Haiech, J.; Kilhoffer, M.C.; Zeniou, M. Cancer stem cell quiescence and plasticity as major challenges in cancer therapy. Stem Cells Int. 2016, 2016, 1740936. [Google Scholar] [PubMed]
- Visvader, J.E.; Lindeman, G.J. Cancer stem cells: Current status and evolving complexities. Cell Stem Cell. 2012, 10, 717–728. [Google Scholar]
- Ajani, J.A.; Song, S.; Hochster, H.S.; Steinberg, I.B. Cancer stem cells: The promise and the potential. Semin. Oncol. 2015, 42 (Suppl. S1), S3–S17. [Google Scholar]
- Yurchenko, V.; Constant, S.; Eisenmesser, E.; Bukrinsky, M. Cyclophilin–CD147 interactions: A new target for anti-inflammatory therapeutics. Clin. Exp. Immunol. 2010, 160, 305–317. [Google Scholar] [PubMed]
- Liu, C.; von Brunn, A.; Zhu, D. Cyclophilin A and CD147: Novel therapeutic targets for the treatment of COVID-19. Med. Drug Discov. 2020, 7, 100056. [Google Scholar]
- Hu, X.; Su, J.; Zhou, Y.; Xie, X.; Peng, C.; Yuan, Z.; Chen, X. Repressing CD147 is a novel therapeutic strategy for malignant melanoma. Oncotarget 2017, 8, 25806–25813. [Google Scholar]
- Obchoei, S.; Wongkhan, S.; Wongkham, C.; Li, M.; Yao, Q.; Chen, C. Cyclophilin A: Potential functions and therapeutic target for human cancer. Med. Sci. Monit. 2009, 15, RA221–RA232. [Google Scholar] [PubMed]
- Chu, M.Y.; Huang, H.C.; Li, E.M.; Xu, L.Y. CypA: A potential target of tumor radiotherapy and/or chemotherapy. Curr. Med. Chem. 2021, 28, 3787–3802. [Google Scholar]
- Han, J.M.; Sohng, J.K.; Lee, W.H.; Oh, T.J.; Jung, H.J. Identification of cyclophilin A as a potential anticancer target of novel nargenicin A1 analog in AGS gastric cancer cells. Int. J. Mol. Sci. 2021, 22, 2473. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Shen, J.; Sun, J.; Jiang, Z.; Fan, J.; Wang, H.; Yu, S.; Long, Y.; Liu, Y.; Bao, H.; et al. Cyclophilin A maintains glioma-Initiating cell stemness by regulating Wnt/β-catenin signaling. Clin. Cancer Res. 2017, 23, 6640–6649. [Google Scholar]
- Meng, Y.; Fan, X.Y.; Yang, L.J.; Xu, B.Q.; He, D.; Xu, Z.; Wu, D.; Wang, B.; Cui, H.Y.; Wang, S.J.; et al. Detachment activated CypA and CD147 induces cancer stem cell potential in non-stem breast cancer cells. Front. Cell Dev. Biol. 2020, 8, 543856. [Google Scholar] [PubMed]
- Kang, M.J.; Kim, H.P.; Lee, K.S.; Yoo, Y.D.; Kwon, Y.T.; Kim, K.M.; Kim, T.Y.; Yi, E.C. Proteomic analysis reveals that CD147/EMMPRIN confers chemoresistance in cancer stem cell-like cells. Proteomics 2013, 13, 1714–1725. [Google Scholar] [CrossRef]
- Wang, P.; Heitman, J. The cyclophilins. Genome Biol. 2005, 6, 226. [Google Scholar] [CrossRef]
- Lang, K.; Schmid, F.X.; Fischer, G. Catalysis of protein folding by prolyl isomerase. Nature 1987, 329, 268–270. [Google Scholar] [CrossRef]
- Schiene, C.; Fischer, G. Enzymes that catalyse the restructuring of proteins. Curr. Opin. Struct. Biol. 2000, 10, 40–45. [Google Scholar]
- Handschumacher, R.E.; Harding, M.W.; Rice, J.; Drugge, R.J.; Speicher, D.W. Cyclophilin: A specific cytosolic binding protein for cyclosporin A. Science 1984, 226, 544–547. [Google Scholar]
- Jin, Z.G.; Melaragno, M.G.; Liao, D.F.; Yan, C.; Haendeler, J.; Suh, Y.A.; Lambeth, J.D.; Berk, B.C. Cyclophilin A is a secreted growth factor induced by oxidative stress. Circ. Res. 2000, 87, 789–796. [Google Scholar] [CrossRef] [PubMed]
- Seko, Y.; Fujimura, T.; Taka, H.; Mineki, R.; Murayama, K.; Nagai, R. Hypoxia followed by reoxygenation induces secretion of cyclophilin A from cultured rat cardiac myocytes. Biochem. Biophys. Res. Commun. 2004, 317, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Sherry, B.; Yarlett, N.; Strupp, A.; Cerami, A. Identification of cyclophilin as a proinflammatory secretory product of lipopolysaccharide-activated macrophages. Proc. Natl. Acad. Sci. USA 1992, 89, 3511–3515. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, J.; Jin, Z.G.; Meoli, D.F.; Matoba, T.; Berk, B.C. Cyclophilin A is secreted by a vesicular pathway in vascular smooth muscle cells. Circ. Res. 2006, 98, 811–817. [Google Scholar] [CrossRef] [PubMed]
- Satoh, K.; Matoba, T.; Suzuki, J.; O’Dell, M.R.; Nigro, P.; Cui, Z.; Mohan, A.; Pan, S.; Li, L.; Jin, Z.G.; et al. Cyclophilin A mediates vascular remodeling by promoting inflammation and vascular smooth muscle cell proliferation. Circulation 2008, 117, 3088–3098. [Google Scholar] [CrossRef]
- Luban, J. Absconding with the chaperone: Essential cyclophilin-Gag interaction in HIV-1 virions. Cell 1996, 87, 1157–1159. [Google Scholar] [CrossRef]
- Bosco, D.A.; Eisenmesser, E.Z.; Pochapsky, S.; Sundquist, W.I.; Kern, D. Catalysis of cis/trans isomerization in native HIV-1 capsid by human cyclophilin A. Proc. Natl. Acad. Sci. USA 2002, 99, 5247–5252. [Google Scholar] [CrossRef]
- Colgan, J.; Asmal, M.; Yu, B.; Luban, J. Cyclophilin A-deficient mice are resistant to immunosuppression by cyclosporine. J. Immunol. 2005, 174, 6030–6038. [Google Scholar] [CrossRef]
- Melo, A.; Monteiro, L.; Lima, R.M.; Oliveira, D.M.; Cerqueira, M.D.; El-Bachá, R.S. Oxidative stress in neurodegenerative diseases: Mechanisms and therapeutic perspectives. Oxidative Med. Cell. Longev. 2011, 2011, 467180. [Google Scholar] [CrossRef]
- Billich, A.; Winkler, G.; Aschauer, H.; Rot, A.; Peichl, P. Presence of cyclophilin A in synovial fluids of patients with rheumatoid arthritis. J. Exp. Med. 1997, 185, 975–980. [Google Scholar] [CrossRef]
- Tegeder, I.; Schumacher, A.; John, S.; Geiger, H.; Geisslinger, G.; Bang, H.; Brune, K. Elevated serum cyclophilin levels in patients with severe sepsis. J. Clin. Immunol. 1997, 17, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Xie, H.; Yi, M.; Peng, L.; Lei, D.; Chen, X.; Jian, D. Expression of cyclophilin A and CD147 during skin aging. Zhong Nan Da Xue Xue Bao Yi Xue Ban 2011, 36, 203–211. [Google Scholar] [PubMed]
- Kalinina, A.; Golubeva, I.; Kudryavtsev, I.; Khromova, N.; Antoshina, E.; Trukhanova, L.; Gorkova, T.; Kazansky, D.; Khromykh, L. Cyclophilin A is a factor of antitumor defense in the early stages of tumor development. Int. Immunopharmacol. 2021, 94, 107470. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Chen, J.; Yang, J.; Qiao, S.; Zhao, S.; Yu, L. Cyclophilin A is upregulated in small cell lung cancer and activates ERK1/2 signal. Biochem. Biophys. Res. Commun. 2007, 361, 763–767. [Google Scholar] [CrossRef]
- Li, Z.; Zhao, X.; Bai, S.; Wang, Z.; Chen, L.; Wei, Y.; Huang, C. Proteomics identification of cyclophilin a as a potential prognostic factor and therapeutic target in endometrial carcinoma. Mol. Cell. Proteom. 2008, 7, 1810–1823. [Google Scholar] [CrossRef]
- Nigro, P.; Pompilio, G.; Capogrossi, M.C. Cyclophilin A: A key player for human disease. Cell Death Dis. 2013, 4, e888. [Google Scholar] [CrossRef]
- Yan, L.; Zucker, S.; Toole, B.P. Roles of the multifunctional glycoprotein, emmprin (basigin; CD147), in tumour progression. Thromb. Haemost. 2005, 93, 199–204. [Google Scholar] [CrossRef]
- Biswas, C.; Zhang, Y.; DeCastro, R.; Guo, H.; Nakamura, T.; Kataoka, H.; Nabeshima, K. The human tumor cell-derived collagenase stimulatory factor (renamed EMMPRIN) is a member of the immunoglobulin superfamily. Cancer Res. 1995, 55, 434–439. [Google Scholar]
- Sameshima, T.; Nabeshima, K.; Toole, B.P.; Yokogami, K.; Okada, Y.; Goya, T.; Koono, M.; Wakisaka, S. Expression of emmprin (CD147), a cell surface inducer of matrix metalloproteinases, in normal human brain and gliomas. Int. J. Cancer 2000, 88, 21–27. [Google Scholar] [CrossRef]
- Chen, X.; Kanekura, T.; Kanzaki, T. Expression of Basigin in human fetal, infantile and adult skin and in basal cell carcinoma. J. Cutan. Pathol. 2001, 28, 184–190. [Google Scholar] [CrossRef]
- Yu, X.L.; Hu, T.; Du, J.M.; Ding, J.P.; Yang, X.M.; Zhang, J.; Yang, B.; Shen, X.; Zhang, Z.; Zhong, W.D.; et al. Crystal structure of HAb18G/CD147: Implications for immunoglobulin superfamily homophilic adhesion. J. Biol. Chem. 2008, 283, 18056–18065. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, S.; Shibata, M.; Yamamoto, S.; Hagihara, M.; Asai, N.; Takahashi, M.; Mizutani, S.; Muramatsu, T.; Kadomatsu, K. Homo-oligomer formation by basigin, an immunoglobulin superfamily member, via its N-terminal immunoglobulin domain. Eur. J. Biochem. 2000, 267, 4372–4380. [Google Scholar] [CrossRef] [PubMed]
- Knutti, N.; Kuepper, M.; Friedrich, K. Soluble extracellular matrix metalloproteinase inducer (EMMPRIN, EMN) regulates cancer-related cellular functions by homotypic interactions with surface CD147. FEBS J. 2015, 282, 4187–4200. [Google Scholar] [CrossRef] [PubMed]
- Landras, A.; Reger de Moura, C.; Jouenne, F.; Lebbe, C.; Menashi, S.; Mourah, S. CD147 is a promising target of tumor progression and a prognostic biomarker. Cancers 2019, 11, 1803. [Google Scholar] [CrossRef]
- Lv, J.J.; Wang, H.; Cui, H.Y.; Liu, Z.K.; Zhang, R.Y.; Lu, M.; Li, C.; Yong, Y.L.; Liu, M.; Zhang, H.; et al. Blockade of macrophage CD147 protects against foam cell formation in atherosclerosis. Front. Cell Dev. Biol. 2021, 8, 609090. [Google Scholar] [CrossRef]
- Luan, J.; Zhang, K.; Yang, P.; Zhang, Y.; Feng, F.; Zhu, Y.M.; Zhu, P.; Chen, Z.N. The combination of FK506 and an anti-CD147 mAb exerts potential therapeutic effects on a mouse model of collagen-induced arthritis. Mol. Immunol. 2018, 101, 1–9. [Google Scholar] [CrossRef]
- Zhou, Y.Q.; Wang, K.; Wang, X.Y.; Cui, H.Y.; Zhao, Y.; Zhu, P.; Chen, Z.N. SARS-CoV-2 pseudovirus enters the host cells through spike protein-CD147 in an Arf6-dependent manner. Emerg. Microbes. Infect. 2022, 11, 1135–1144. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.Y.; He, D.; Sheng, C.B.; Wang, B.; Wang, L.J.; Wu, X.Q.; Xu, L.; Jiang, J.L.; Li, L.; Chen, Z.N. Therapeutic anti-CD147 antibody sensitizes cells to chemoradiotherapy via targeting pancreatic cancer stem cells. Am. J. Transl. Res. 2019, 11, 3543–3554. [Google Scholar] [PubMed]
- Lucchetti, D.; Colella, F.; Perelli, L.; Ricciardi-Tenore, C.; Calapà, F.; Fiori, M.E.; Carbone, F.; De Maria, R.; Sgambato, A. CD147 promotes cell small extracellular vesicles release during colon cancer stem cells differentiation and triggers cellular changes in recipient cells. Cancers 2020, 12, 260. [Google Scholar] [CrossRef]
- Pushkarsky, T.; Zybarth, G.; Dubrovsky, L.; Yurchenko, V.; Tang, H.; Guo, H.; Toole, B.; Sherry, B.; Bukrinsky, M. CD147 facilitates HIV-1 infection by interacting with virus-associated cyclophilin A. Proc. Natl. Acad. Sci. USA 2001, 98, 6360–6365. [Google Scholar] [CrossRef]
- Yurchenko, V.; Zybarth, G.; O’Connor, M.; Dai, W.W.; Franchin, G.; Hao, T.; Guo, H.; Hung, H.C.; Toole, B.; Gallay, P.; et al. Active site residues of cyclophilin A are crucial for its signaling activity via CD147. J. Biol. Chem. 2002, 277, 22959–22965. [Google Scholar] [CrossRef] [PubMed]
- Schlegel, J.; Redzic, J.S.; Porter, C.C.; Yurchenko, V.; Bukrinsky, M.; Labeikovsky, W.; Armstrong, G.S.; Zhang, F.; Isern, N.G.; DeGregori, J.; et al. Solution characterization of the extracellular region of CD147 and its interaction with its enzyme ligand cyclophilin A. J. Mol. Biol. 2009, 391, 518–535. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Zang, Y.; Wang, H.; Kang, Y.; Zhang, J.; Li, X.; Zhang, L.; Zhang, S. Recognition between CD147 and cyclophilin A deciphered by accelerated molecular dynamics simulations. Phys. Chem. Chem. Phys. 2022, 24, 18905–18914. [Google Scholar] [CrossRef] [PubMed]
- Howard, B.A.; Furumai, R.; Campa, M.J.; Rabbani, Z.N.; Vujaskovic, Z.; Wang, X.F.; Patz, E.F., Jr. Stable RNA interference-mediated suppression of cyclophilin A diminishes non-small-cell lung tumor growth in vivo. Cancer Res. 2005, 65, 8853–8860. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Harris, S.L.; Levine, A.J. The regulation of exosome secretion: A novel function of the p53 protein. Cancer Res. 2006, 66, 4795–4801. [Google Scholar] [CrossRef]
- Choi, K.J.; Piao, Y.J.; Lim, M.J.; Kim, J.H.; Ha, J.; Choe, W.; Kim, S.S. Overexpressed cyclophilin A in cancer cells renders resistance to hypoxia- and cisplatin-induced cell death. Cancer Res. 2007, 67, 3654–3662. [Google Scholar] [CrossRef]
- Obchoei, S.; Weakley, S.M.; Wongkham, S.; Wongkham, C.; Sawanyawisuth, K.; Yao, Q.; Chen, C. Cyclophilin A enhances cell proliferation and tumor growth of liver fluke-associated cholangiocarcinoma. Mol. Cancer 2011, 10, 102. [Google Scholar] [CrossRef]
- Li, M.; Zhai, Q.; Bharadwaj, U.; Wang, H.; Li, F.; Fisher, W.E.; Chen, C.; Yao, Q. Cyclophilin A is overexpressed in human pancreatic cancer cells and stimulates cell proliferation through CD147. Cancer 2006, 106, 2284–2294. [Google Scholar] [CrossRef]
- Sakamoto, M.; Miyagaki, T.; Kamijo, H.; Oka, T.; Boki, H.; Takahashi-Shishido, N.; Suga, H.; Sugaya, M.; Sato, S. CD147-cyclophilin a interactions promote proliferation and survival of cutaneous T-cell lymphoma. Int. J. Mol. Sci. 2021, 22, 7889. [Google Scholar] [CrossRef]
- Sun, S.; Guo, M.; Zhang, J.B.; Ha, A.; Yokoyama, K.K.; Chiu, R.H. Cyclophilin A (CypA) interacts with NF-κB subunit, p65/RelA, and contributes to NF-κB activation signaling. PLoS ONE 2014, 9, e96211. [Google Scholar] [CrossRef]
- Sun, S.; Wang, Q.; Giang, A.; Cheng, C.; Soo, C.; Wang, C.Y.; Liau, L.M.; Chiu, R. Knockdown of CypA inhibits interleukin-8 (IL-8) and IL-8-mediated proliferation and tumor growth of glioblastoma cells through down-regulated NF-κB. J. Neurooncol. 2011, 101, 1–14. [Google Scholar] [PubMed]
- Gong, Z.; Chi, C.; Huang, X.; Chu, H.; Wang, J.; Du, F.; Jiang, L.; Chen, J. Cyclophilin A is overexpressed in hepatocellular carcinoma and is associated with the cell cycle. Anticancer Res. 2017, 37, 4443–4447. [Google Scholar] [PubMed]
- Ramadori, G.; Ioris, R.M.; Villanyi, Z.; Firnkes, R.; Panasenko, O.O.; Allen, G.; Konstantinidou, G.; Aras, E.; Brenachot, X.; Biscotti, T.; et al. FKBP10 regulates protein translation to sustain lung cancer growth. Cell Rep. 2020, 30, 3851–3863. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Liu, P.; Yan, Y.; Fang, K.; Liang, D.; Hou, X.; Zhang, X.; Wu, S.; Ma, J.; Wang, R.; et al. FKBP9 promotes the malignant behavior of glioblastoma cells and confers resistance to endoplasmic reticulum stress inducers. J. Exp. Clin. Cancer Res. 2020, 39, 44. [Google Scholar]
- Volker, S.E.; Hedrick, S.E.; Feeney, Y.B.; Clevenger, C.V. Cyclophilin A function in mammary epithelium impacts Jak2/Stat5 signaling, morphogenesis, differentiation, and tumorigenesis in the mammary gland. Cancer Res. 2018, 78, 3877–3887. [Google Scholar]
- Li, L.; Tang, W.; Wu, X.; Karnak, D.; Meng, X.; Thompson, R.; Hao, X.; Li, Y.; Qiao, X.T.; Lin, J.; et al. HAb18G/CD147 promotes pSTAT3-mediated pancreatic cancer development via CD44s. Clin. Cancer Res. 2013, 19, 6703–6715. [Google Scholar]
- Peng, J.; Jiang, H.; Guo, J.; Huang, J.; Yuan, Q.; Xie, J.; Xiao, K. CD147 expression is associated with tumor proliferation in bladder cancer via GSDMD. BioMed Res. Int. 2020, 2020, 7638975. [Google Scholar]
- Guo, Y.; Jiang, M.; Zhao, X.; Gu, M.; Wang, Z.; Xu, S.; Yue, W. Cyclophilin A promotes non-small cell lung cancer metastasis via p38 MAPK. Thorac. Cancer 2018, 9, 120–128. [Google Scholar] [CrossRef]
- Li, Y.; Guo, H.; Dong, D.; Wu, H.; Li, E. Expression and prognostic relevance of Cyclophilin A and matrix metalloproteinase 9 in esophageal squamous cell carcinoma. Diagn. Pathol. 2013, 8, 207. [Google Scholar] [CrossRef]
- Chen, X.; Lin, J.; Kanekura, T.; Su, J.; Lin, W.; Xie, H.; Wu, Y.; Li, J.; Chen, M.; Chang, J. A small interfering CD147-targeting RNA inhibited the proliferation, invasiveness, and metastatic activity of malignant melanoma. Cancer Res. 2006, 66, 11323–11330. [Google Scholar] [CrossRef]
- Long, T.; Su, J.; Tang, W.; Luo, Z.; Liu, S.; Liu, Z.; Zhou, H.; Qi, M.; Zeng, W.; Zhang, J.; et al. A novel interaction between calcium-modulating cyclophilin ligand and Basigin regulates calcium signaling and matrix metalloproteinase activities in human melanoma cells. Cancer Lett. 2013, 339, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Hao, J.; Madigan, M.C.; Khatri, A.; Power, C.A.; Hung, T.T.; Beretov, J.; Chang, L.; Xiao, W.; Cozzi, P.J.; Graham, P.H.; et al. In vitro and in vivo prostate cancer metastasis and chemoresistance can be modulated by expression of either CD44 or CD147. PLoS ONE 2012, 7, e40716. [Google Scholar] [CrossRef] [PubMed]
- Grigoryeva, E.S.; Cherdyntseva, N.V.; Karbyshev, M.S.; Volkomorov, V.V.; Stepanov, I.V.; Zavyalova, M.V.; Perelmuter, V.M.; Buldakov, M.A.; Afanasjev, S.G.; Tuzikov, S.A.; et al. Expression of cyclophilin A in gastric adenocarcinoma patients and its inverse association with local relapses and distant metastasis. Pathol. Oncol. Res. 2014, 20, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Chen, X.; Zhang, Y.; Gu, H.; Bai, Y. CD147 and VEGF co-expression predicts prognosis in patients with acute myeloid leukemia. Jpn. J. Clin. Oncol. 2010, 40, 1046–1052. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Xu, C.; Niu, R.; Hu, G.; Gu, Z.; Zhuang, Z. MiR-890 inhibits proliferation and invasion and induces apoptosis in triple-negative breast cancer cells by targeting CD147. BMC Cancer 2019, 19, 577. [Google Scholar] [CrossRef] [PubMed]
- Kuang, Y.H.; Chen, X.; Su, J.; Wu, L.S.; Liao, L.Q.; Li, D.; Chen, Z.S.; Kanekura, T. RNA interference targeting the CD147 induces apoptosis of multi-drug resistant cancer cells related to XIAP depletion. Cancer Lett. 2009, 276, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Tsujino, I.; Fukunaga, M.; Mizumura, K.; Gon, Y.; Takahashi, N.; Hashimoto, S. Cyclosporine A induces apoptosis of human lung adenocarcinoma cells via caspase-dependent pathway. Anticancer Res. 2011, 31, 2129–2134. [Google Scholar]
- Franke, T.F.; Hornik, C.P.; Segev, L.; Shostak, G.A.; Sugimoto, C. PI3K/Akt and apoptosis: Size matters. Oncogene 2003, 22, 8983–8998. [Google Scholar] [CrossRef]
- Ma, Z.; Zhang, W.; Wu, Y.; Zhang, M.; Wang, L.; Wang, Y.; Wang, Y.; Liu, W. Cyclophilin A inhibits A549 cell oxidative stress and apoptosis by modulating the PI3K/Akt/mTOR signaling pathway. Biosci. Rep. 2021, 41, BSR20203219. [Google Scholar] [CrossRef]
- Jiang, X.; Zhang, Q.L.; Tian, Y.H.; Huang, J.C.; Ma, G.L. RNA interference-mediated gene silencing of cyclophilin A enhances the radiosensitivity of PAa human lung adenocarcinoma cells in vitro. Oncol. Lett. 2017, 13, 1619–1624. [Google Scholar] [CrossRef]
- Xiong, L.; Ding, L.; Ning, H.; Wu, C.; Fu, K.; Wang, Y.; Zhang, Y.; Liu, Y.; Zhou, L. CD147 knockdown improves the antitumor efficacy of trastuzumab in HER2-positive breast cancer cells. Oncotarget 2016, 7, 57737–57751. [Google Scholar] [CrossRef] [PubMed]
- Jiang, K.; He, B.; Lai, L.; Chen, Q.; Liu, Y.; Guo, Q.; Wang, Q. Cyclosporine A inhibits breast cancer cell growth by downregulating the expression of pyruvate kinase subtype M2. Int. J. Mol. Med. 2012, 30, 302–308. [Google Scholar] [CrossRef]
- Dhakal, D.; Han, J.M.; Mishra, R.; Pandey, R.P.; Kim, T.S.; Rayamajhi, V.; Jung, H.J.; Yamaguchi, T.; Sohng, J.K. Characterization of tailoring steps of nargenicin A1 biosynthesis reveals a novel analogue with anticancer activities. ACS Chem. Biol. 2020, 15, 1370–1380. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Hu, C.; Xiao, Z.; Luo, C.; Liu, Z. Downregulation of CyclophilinA/CD147 axis induces cell apoptosis and inhibits glioma aggressiveness. BioMed Res. Int. 2020, 2020, 7035847. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhang, M.; Ma, H.; Saiyin, H.; Shen, S.; Xi, J.; Wan, B.; Yu, L. Oligo-microarray analysis reveals the role of cyclophilin A in drug resistance. Cancer Chemother. Pharmacol. 2008, 61, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Jiang, J.; Chen, H.N.; Zhou, L.; Huang, Z.; Qin, S.; Jin, P.; Luo, M.; Li, B.; Shi, J.; et al. Redox-sensitive cyclophilin A elicits chemoresistance through realigning cellular oxidative status in colorectal cancer. Cell Rep. 2021, 37, 110069. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Li, Y.; Dang, Y.Z.; Gao, H.X.; Jiang, J.L.; Chen, Z.N. HAb18G/CD147 promotes radioresistance in hepatocellular carcinoma cells: A potential role for integrin β1 signaling. Mol. Cancer Ther. 2015, 14, 553–563. [Google Scholar] [CrossRef]
- Dubrovska, A.; Kim, S.; Salamone, R.J.; Walker, J.R.; Maira, S.M.; García-Echeverría, C.; Schultz, P.G.; Reddy, V.A. The role of PTEN/Akt/PI3K signaling in the maintenance and viability of prostate cancer stem-like cell populations. Proc. Natl. Acad. Sci. USA 2009, 106, 268–273. [Google Scholar] [CrossRef]
- Lemma, S.; Avnet, S.; Salerno, M.; Chano, T.; Baldini, N. Identification and validation of housekeeping genes for gene expression analysis of cancer stem cells. PLoS ONE 2016, 11, e0149481. [Google Scholar] [CrossRef]
- Rustighi, A.; Zannini, A.; Tiberi, L.; Sommaggio, R.; Piazza, S.; Sorrentino, G.; Nuzzo, S.; Tuscano, A.; Eterno, V.; Benvenuti, F.; et al. Prolyl-isomerase Pin1 controls normal and cancer stem cells of the breast. EMBO Mol. Med. 2013, 6, 99–119. [Google Scholar] [CrossRef]
- Lv, Y.; Wang, T.; Fan, J.; Zhang, Z.; Zhang, J.; Xu, C.; Li, Y.; Zhao, G.; He, C.; Meng, H.; et al. The effects and mechanisms of SLC34A2 on maintaining stem cell-like phenotypes in CD147+ breast cancer stem cells. Tumour Biol. 2017, 39, 1010428317695927. [Google Scholar] [CrossRef] [PubMed]
- Zenke, G.; Strittmatter, U.; Fuchs, S.; Quesniaux, V.F.; Brinkmann, V.; Schuler, W.; Zurini, M.; Enz, A.; Billich, A.; Sanglier, J.J.; et al. Sanglifehrin A, a novel cyclophilin-binding compound showing immunosuppressive activity with a new mechanism of action. J. Immunol. 2001, 166, 7165–7171. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.S.; Jia, P.F.; Zhang, Z.Q.; Zhang, S.M. ROS-p53-cyclophilin-D signaling mediates salinomycin-induced glioma cell necrosis. J. Exp. Clin. Cancer Res. 2015, 34, 57. [Google Scholar] [CrossRef] [PubMed]
- Kallen, J.; Sedrani, R.; Zenke, G.; Wagner, J. Structure of Human Cyclophilin A in Complex with the Novel Immunosuppressant Sanglifehrin A at 1.6 Å Resolution. J. Biol. Chem. 2005, 280, 21965–21971. [Google Scholar] [CrossRef]
- Stähelin, H.F. The history of cyclosporin A (Sandimmune) revisited: Another point of view. Experientia 1996, 52, 5–13. [Google Scholar] [CrossRef]
- Davra, V.; Saleh, T.; Geng, K.; Kimani, S.; Mehta, D.; Kasikara, C.; Smith, B.; Colangelo, N.W.; Ciccarelli, B.; Li, H.; et al. Cyclophilin A inhibitor Debio-025 targets Crk, reduces metastasis, and induces tumor immunogenicity in breast cancer. Mol. Cancer Res. 2020, 18, 1189–1201. [Google Scholar] [CrossRef]
- Lee, J. Cyclophilin A as a new therapeutic target for hepatitis C virus-induced hepatocellular carcinoma. Korean J. Physiol. Pharmacol. 2013, 17, 375–383. [Google Scholar] [CrossRef]
- Simón Serrano, S.; Tavecchio, M.; Grönberg, A.; Sime, W.; Jemaà, M.; Moss, S.; Gregory, M.A.; Gallay, P.; Elmér, E.; Hansson, M.J.; et al. Novel cyclophilin inhibitor decreases cell proliferation and tumor growth in models of hepatocellular carcinoma. Cancers 2021, 13, 3041. [Google Scholar] [CrossRef]
- Han, J.M.; Choi, Y.S.; Dhakal, D.; Sohng, J.K.; Jung, H.J. Novel nargenicin A1 analog inhibits angiogenesis by downregulating the endothelial VEGF/VEGFR2 signaling and tumoral HIF-1α/VEGF pathway. Biomedicines 2020, 8, 252. [Google Scholar] [CrossRef]
- Wang, J.; Li, F.; Tan, J.; Peng, X.; Sun, L.; Wang, P.; Jia, S.; Yu, Q.; Huo, H.; Zhao, H. Melittin inhibits the invasion of MCF-7 cells by downregulating CD147 and MMP-9 expression. Oncol. Lett. 2017, 13, 599–604. [Google Scholar] [CrossRef]
- Fu, Z.G.; Wang, L.; Cui, H.Y.; Peng, J.L.; Wang, S.J.; Geng, J.J.; Liu, J.D.; Feng, F.; Song, F.; Li, L.; et al. A novel small-molecule compound targeting CD147 inhibits the motility and invasion of hepatocellular carcinoma cells. Oncotarget 2016, 7, 9429–9447. [Google Scholar] [CrossRef] [PubMed]
- Spinello, I.; Saulle, E.; Quaranta, M.T.; Pasquini, L.; Pelosi, E.; Castelli, G.; Ottone, T.; Voso, M.T.; Testa, U.; Labbaye, C. The small-molecule compound AC-73 targeting CD147 inhibits leukemic cell proliferation, induces autophagy and increases the chemotherapeutic sensitivity of acute myeloid leukemia cells. Haematologica 2019, 104, 973–985. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Shen, Z.Y.; Chen, X.G.; Zhang, Q.; Bian, H.J.; Zhu, P.; Xu, H.Y.; Song, F.; Yang, X.M.; Mi, L.; et al. A randomized controlled trial of Licartin for preventing hepatoma recurrence after liver transplantation. Hepatology 2007, 45, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Bian, H.; Zheng, J.S.; Nan, G.; Li, R.; Chen, C.; Hu, C.X.; Zhang, Y.; Sun, B.; Wang, X.L.; Cui, S.C.; et al. Randomized trial of [131I] metuximab in treatment of hepatocellular carcinoma after percutaneous radiofrequency ablation. J. Natl. Cancer Inst. 2014, 106, dju239. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Pan, Y.; He, B.; Ying, H.; Wang, F.; Sun, H.; Deng, Q.; Liu, X.; Lin, K.; Peng, H.; et al. Inhibition of CD147 expression by RNA interference reduces proliferation, invasion and increases chemosensitivity in cancer stem cell-like HT-29 cells. Int. J. Oncol. 2015, 47, 1476–1484. [Google Scholar] [CrossRef]
- Haendler, B.; Hofer, E. Characterization of the human cyclophilin gene and of related processed pseudogenes. Eur. J. Biochem. 1990, 190, 477–482. [Google Scholar] [CrossRef]
- Xin, X.; Zeng, X.; Gu, H.; Li, M.; Tan, H.; Jin, Z.; Hua, T.; Shi, R.; Wang, H. CD147/EMMPRIN overexpression and prognosis in cancer: A systematic review and meta-analysis. Sci. Rep. 2016, 6, 32804. [Google Scholar] [CrossRef]
- Van Calster, B.; Van Hoorde, K.; Valentin, L.; Testa, A.C.; Fischerova, D.; Van Holsbeke, C.; Savelli, L.; Franchi, D.; Epstein, E.; Kaijser, J.; et al. Evaluating the risk of ovarian cancer before surgery using the ADNEX model to differentiate between benign, borderline, early and advanced stage invasive, and secondary metastatic tumours: Prospective multicentre diagnostic study. BMJ 2014, 349, g5920. [Google Scholar] [CrossRef]
- Chandrashekar, D.S.; Bashel, B.; Balasubramanya, S.A.H.; Creighton, C.J.; Ponce-Rodriguez, I.; Chakravarthi, B.V.S.K.; Varambally, S. UALCAN: A portal for facilitating tumor subgroup gene expression and survival analyses. Neoplasia 2017, 19, 649–658. [Google Scholar] [CrossRef]
- Li, T.; Fan, J.; Wang, B.; Traugh, N.; Chen, Q.; Liu, J.S.; Li, B.; Liu, X.S. TIMER: A web server for comprehensive analysis of tumor-infiltrating immune cells. Cancer Res. 2017, 77, e108–e110. [Google Scholar] [CrossRef]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci. Signal. 2013, 6, pl1. [Google Scholar] [CrossRef] [PubMed]










| TCGA Samples | Expression of PPIA | |||
|---|---|---|---|---|
| Normal Samples | Tumor Samples | |||
| Number of Samples | Median Expression (Log2) | Number of Samples | Median Expression (Log2) | |
| BLCA | 19 | 6.985▼ | 408 | 7.691▲ |
| BRCA | 114 | 7.077▼ | 1097 | 7.572▲ |
| CESC | 3 | 6.748▼ | 305 | 7.728▲ |
| CHOL | 9 | 5.485▼ | 36 | 7.41▲ |
| COAD | 41 | 7.013▼ | 286 | 7.756▲ |
| ESCA | 11 | 6.031▼ | 184 | 7.842▲ |
| GBM | 5 | 7.263▼ | 156 | 8.328▲ |
| HNSC | 44 | 6.78▼ | 520 | 7.474▲ |
| KICH | 25 | 7.143▼ | 67 | 7.217▲ |
| KIRC | 72 | 7.074▼ | 533 | 7.27▲ |
| KIRP | 32 | 6.807▼ | 290 | 7.504▲ |
| LIHC | 50 | 5.672▼ | 371 | 6.883▲ |
| LUAD | 59 | 6.89▼ | 515 | 7.387▲ |
| LUSC | 52 | 6.916▼ | 503 | 7.613▲ |
| PAAD | 4 | 7.009▼ | 178 | 7.178▲ |
| PRAD | 52 | 6.729▼ | 497 | 7.035▲ |
| PCPG | 3 | 6.686▼ | 179 | 6.994▲ |
| READ | 10 | 7.106▼ | 166 | 8.21▲ |
| SARC | 2 | 6.672▼ | 260 | 7.044▲ |
| SKCM | 1 | 7.477▼ | 472 | 7.662▲ |
| THCA | 59 | 6.85▼ | 505 | 6.876▲ |
| THYM | 2 | 7.393▼ | 120 | 8.092▲ |
| STAD | 34 | 6.592▼ | 415 | 7.623▲ |
| UCEC | 35 | 6.905▼ | 546 | 8.881▲ |
| TCGA Samples | Expression of BSG | |||
|---|---|---|---|---|
| Normal Samples | Tumor Samples | |||
| Number of Samples | Median Expression (Log2) | Number of Samples | Median Expression (Log2) | |
| BLCA | 19 | 9.097▼ | 408 | 9.437▲ |
| BRCA | 114 | 8.505▼ | 1097 | 9.027▲ |
| CESC | 3 | 9.126▼ | 305 | 9.276▲ |
| CHOL | 9 | 6.909▼ | 36 | 9.00▲ |
| COAD | 41 | 10.581▲ | 286 | 9.653▼ |
| ESCA | 11 | 8.682▼ | 184 | 9.39▲ |
| GBM | 5 | 9.608▲ | 156 | 9.575▼ |
| HNSC | 44 | 8.455▼ | 520 | 9.286▲ |
| KICH | 25 | 10.039▼ | 67 | 11.292▲ |
| KIRC | 72 | 9.674▲ | 533 | 9.535▼ |
| KIRP | 32 | 10.062▼ | 290 | 10.475▲ |
| LIHC | 50 | 6.894▼ | 371 | 8.409▲ |
| LUAD | 59 | 8.916▼ | 515 | 9.144▲ |
| LUSC | 52 | 8.76▼ | 503 | 9.033▲ |
| PAAD | 4 | 9.01▼ | 178 | 9.12▲ |
| PRAD | 52 | 9.073▼ | 497 | 9.368▲ |
| PCPG | 3 | 9.959▼ | 179 | 10.077▲ |
| READ | 10 | 9.925▲ | 166 | 9.487▼ |
| SARC | 2 | 9.879▲ | 260 | 9.384▼ |
| SKCM | 1 | 9.364▼ | 472 | 10.145▲ |
| THCA | 59 | 9.421▼ | 505 | 9.61▲ |
| THYM | 2 | 8.774▼ | 120 | 9.056▲ |
| STAD | 34 | 9.015▼ | 415 | 9.514▲ |
| UCEC | 35 | 9.347▼ | 546 | 10.024▲ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, J.M.; Jung, H.J. Cyclophilin A/CD147 Interaction: A Promising Target for Anticancer Therapy. Int. J. Mol. Sci. 2022, 23, 9341. https://doi.org/10.3390/ijms23169341
Han JM, Jung HJ. Cyclophilin A/CD147 Interaction: A Promising Target for Anticancer Therapy. International Journal of Molecular Sciences. 2022; 23(16):9341. https://doi.org/10.3390/ijms23169341
Chicago/Turabian StyleHan, Jang Mi, and Hye Jin Jung. 2022. "Cyclophilin A/CD147 Interaction: A Promising Target for Anticancer Therapy" International Journal of Molecular Sciences 23, no. 16: 9341. https://doi.org/10.3390/ijms23169341
APA StyleHan, J. M., & Jung, H. J. (2022). Cyclophilin A/CD147 Interaction: A Promising Target for Anticancer Therapy. International Journal of Molecular Sciences, 23(16), 9341. https://doi.org/10.3390/ijms23169341