
Tubular IKKβ Deletion Alleviates Acute Ischemic Kidney Injury and Facilitates Tissue Regeneration

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
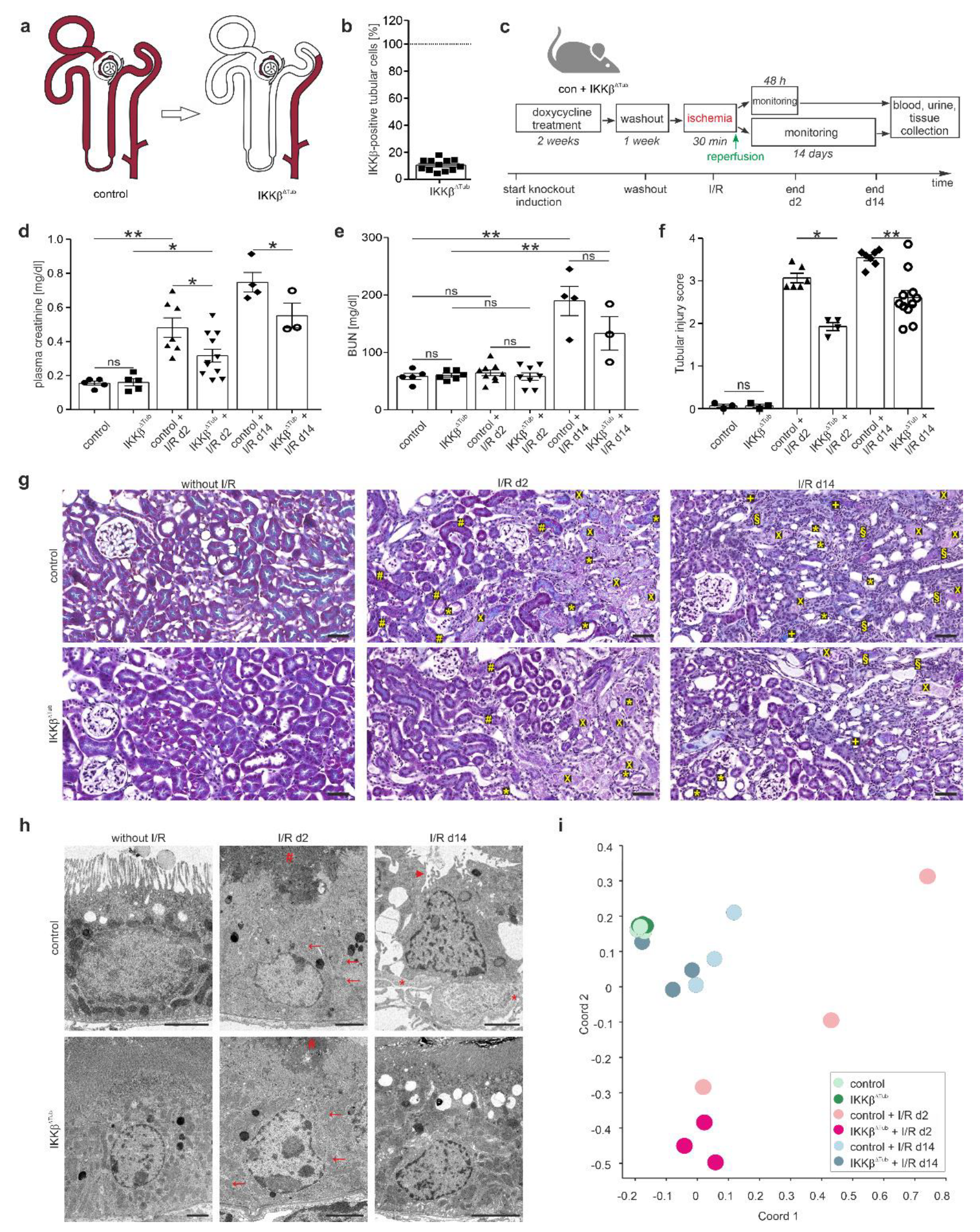
2.1. Genetic Tubular IKKβ-Deletion Ameliorates I/R Injury and Reinforces Tissue Regeneration
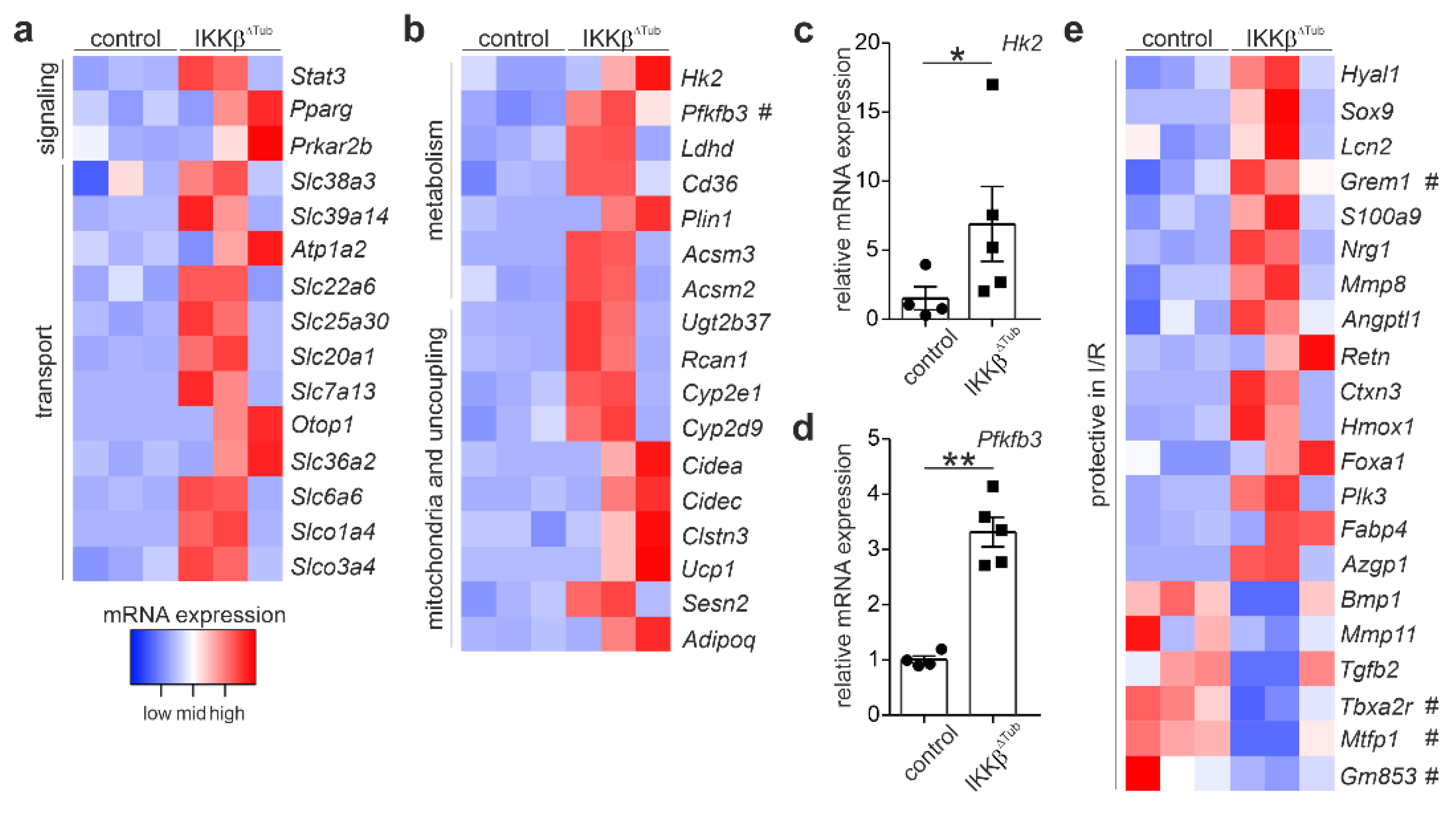
2.2. Comparison of Gene Expression Levels of IKKβ∆Tub Mice Compared to Control at Baseline
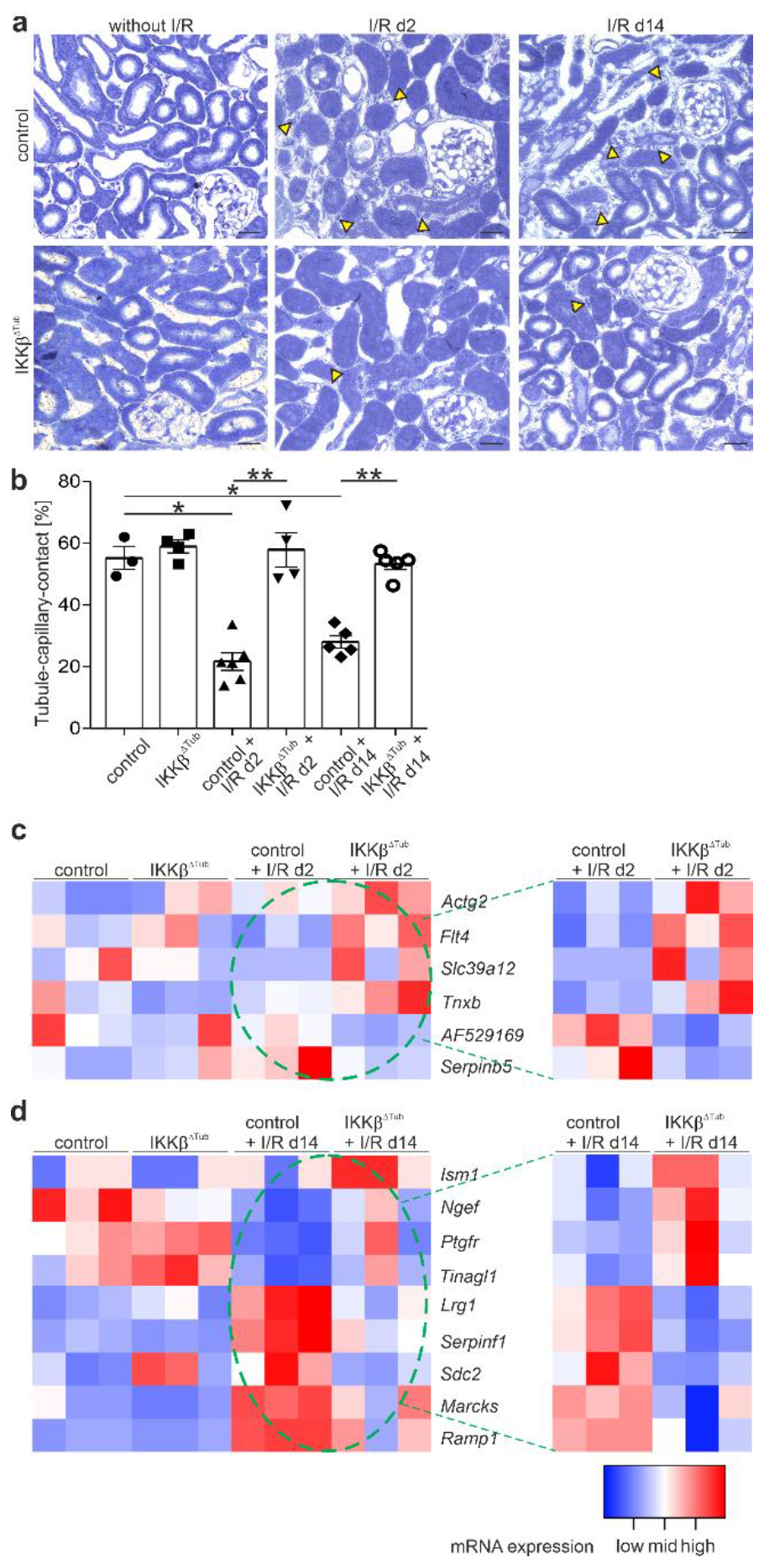
2.3. Tubular Deletion of IKKβ Promotes Angiogenesis after I/R Injury and Preserves Tubulointerstitial Architecture
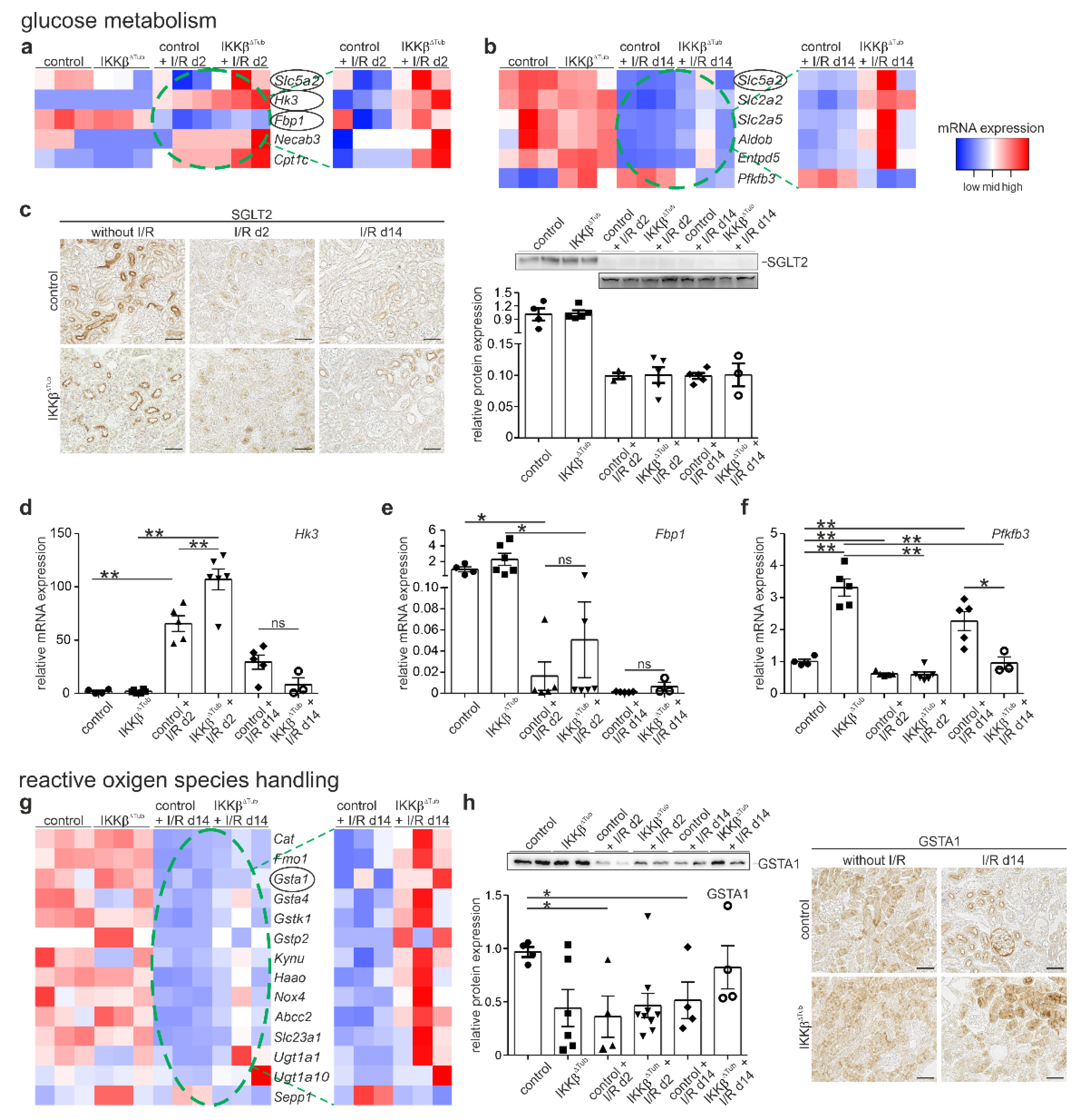
2.4. Tubular Deletion of IKKβ Affect Glucose Metabolism after I/R Injury
2.5. Tubular Deletion of IKKβ Improves Detoxification and Reactive Oxygen Species Handling after I/R Injury
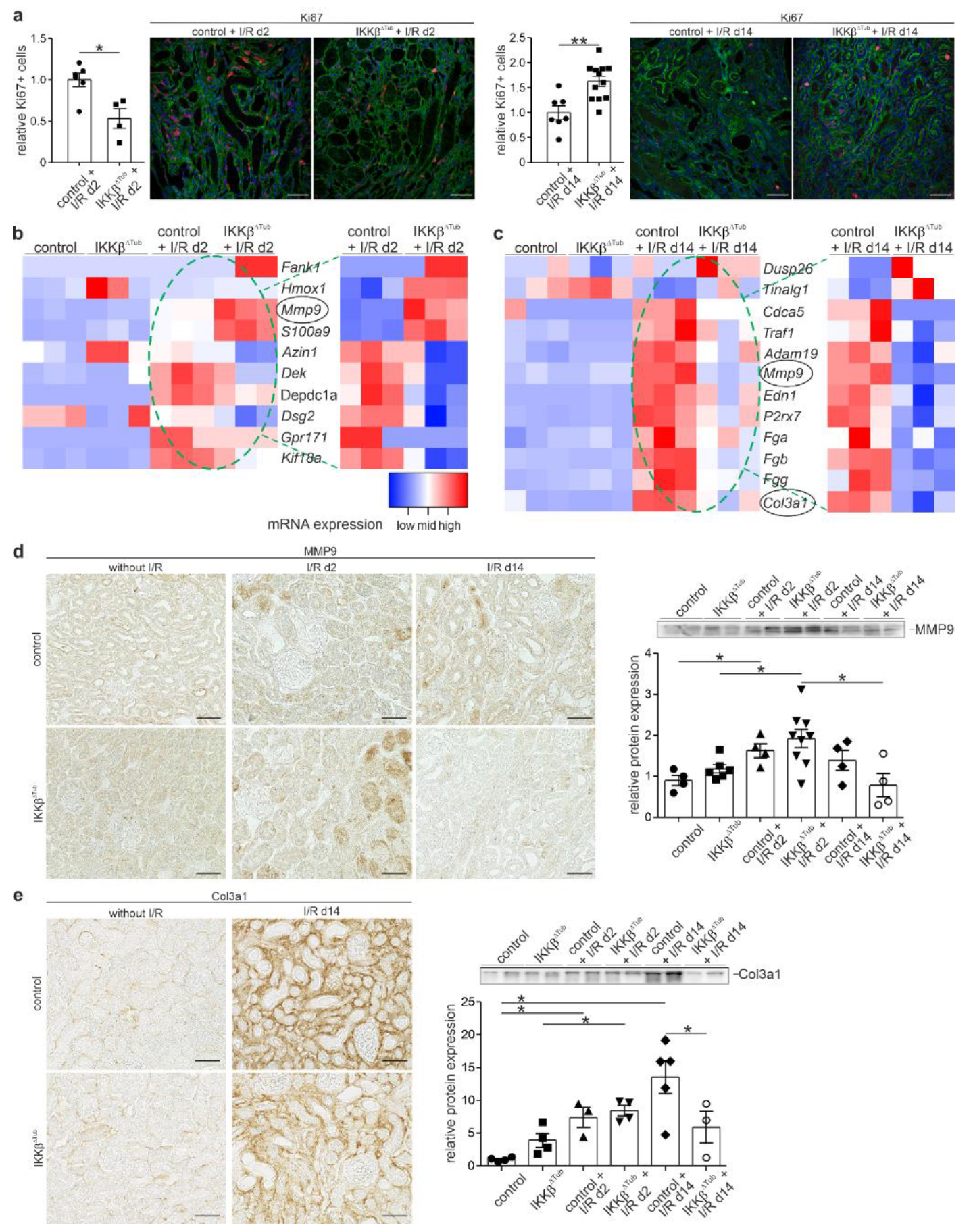
2.6. Tubular Deletion of IKKβ Improves Proliferation, Tissue Regeneration and Reduces Fibrosis
3. Discussion
3.1. Tubular IKKβ Deletion Induced Angiogenesis
3.2. Tubular IKKβ Deletion Improves Glucose Metabolism
3.3. Tubular IKKβ Deletion Improves Kidney Antioxidant Defense
4. Materials and Methods
4.1. Animal Experimentation
4.2. Blood and Urine Collection/Analysis
4.3. Fixation and Tissue Processing for Immunohistochemistry and Immunoblotting
4.4. RNA Isolation, Reverse Transcription, Real-Time PCR and RNA Sequencing
4.5. RNA-Seq, Sequence Details
4.6. RNA In Situ Hybridisation
4.7. SDS-PAGE and Immunoblotting
4.8. Immunohistochemistry
4.9. Cellular Proliferation
4.10. Antibodies
4.11. Histology and Tubular Injury Score
4.12. Electron Microscopy and Morphometry
4.13. Statistical Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rewa, O.; Bagshaw, S.M. Acute kidney injury-epidemiology, outcomes and economics. Nat. Rev. Nephrol. 2014, 10, 193–207. [Google Scholar] [CrossRef]
- Reid, S.; Scholey, J.W. Recent Approaches to Targeting Canonical NFkappaB Signaling in the Early Inflammatory Response to Renal IRI. J. Am. Soc. Nephrol. 2021, 32, 2117–2124. [Google Scholar] [CrossRef]
- Slegtenhorst, B.R.; Dor, F.J.; Rodriguez, H.; Voskuil, F.J.; Tullius, S.G. Ischemia/reperfusion Injury and its Consequences on Immunity and Inflammation. Curr. Transplant. Rep. 2014, 1, 147–154. [Google Scholar] [CrossRef]
- Marko, L.; Vigolo, E.; Hinze, C.; Park, J.K.; Roel, G.; Balogh, A.; Choi, M.; Wubken, A.; Cording, J.; Blasig, I.E.; et al. Tubular Epithelial NF-kappaB Activity Regulates Ischemic AKI. J. Am. Soc. Nephrol. 2016, 27, 2658–2669. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Lee, H.H.; Noriega, M.L.; Paust, H.J.; Zahner, G.; Thaiss, F. Lymphocyte-specific deletion of IKK2 or NEMO mediates an increase in intrarenal Th17 cells and accelerates renal damage in an ischemia-reperfusion injury mouse model. Am. J. Physiol. Ren. Physiol. 2016, 311, F1005–F1014. [Google Scholar] [CrossRef] [PubMed]
- Song, N.; Thaiss, F.; Guo, L. NFkappaB and Kidney Injury. Front. Immunol. 2019, 10, 815. [Google Scholar] [CrossRef]
- Muller, I.I.; Schneider, M.; Muller, K.A.L.; Lunov, O.; Borst, O.; Simmet, T.; Gawaz, M. Protective role of Gremlin-1 in myocardial function. Eur, J. Clin. Investig. 2021, 51, e13539. [Google Scholar] [CrossRef] [PubMed]
- Nagatani, Y.; Higashino, T.; Kinoshita, K.; Higashino, H. Thromboxane A2 receptor antagonist (ONO-8809) attenuates renal disorders caused by salt overload in stroke-prone spontaneously hypertensive rats. Clin. Exp. Pharmacol. Physiol. 2021, 48, 1391–1401. [Google Scholar] [CrossRef] [PubMed]
- Basile, D.P.; Collett, J.A.; Yoder, M.C. Endothelial colony-forming cells and pro-angiogenic cells: Clarifying definitions and their potential role in mitigating acute kidney injury. Acta Physiol. 2018, 222, e12914. [Google Scholar] [CrossRef]
- Antonia, R.J.; Hagan, R.S.; Baldwin, A.S. Expanding the View of IKK: New Substrates and New Biology. Trends Cell Biol. 2021, 31, 166–178. [Google Scholar] [CrossRef]
- Legouis, D.; Ricksten, S.E.; Faivre, A.; Verissimo, T.; Gariani, K.; Verney, C.; Galichon, P.; Berchtold, L.; Feraille, E.; Fernandez, M.; et al. Altered proximal tubular cell glucose metabolism during acute kidney injury is associated with mortality. Nat. Metab. 2020, 2, 732–743. [Google Scholar] [CrossRef] [PubMed]
- Reid, M.A.; Lowman, X.H.; Pan, M.; Tran, T.Q.; Warmoes, M.O.; Ishak Gabra, M.B.; Yang, Y.; Locasale, J.W.; Kong, M. IKKbeta promotes metabolic adaptation to glutamine deprivation via phosphorylation and inhibition of PFKFB3. Genes Dev. 2016, 30, 1837–1851. [Google Scholar] [CrossRef]
- Gao, X.; Han, L.; Yao, X.; Ma, L. Gremlin1 and TGF-beta1 protect kidney tubular epithelial cells from ischemia-reperfusion injury through different pathways. Int. Urol. Nephrol. 2022, 54, 1311–1321. [Google Scholar] [CrossRef] [PubMed]
- Nossent, A.Y.; Hansen, J.L.; Doggen, C.; Quax, P.H.; Sheikh, S.P.; Rosendaal, F.R. SNPs in microRNA binding sites in 3′-UTRs of RAAS genes influence arterial blood pressure and risk of myocardial infarction. Am. J. Hypertens 2011, 24, 999–1006. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Zhou, Y.; Li, S. Association of TBXA2R, P2Y12 and ADD1 genes polymorphisms with ischemic stroke susceptibility: A metaanalysis. Clin. Investig. Med. 2020, 43, E33–E43. [Google Scholar] [CrossRef]
- Long, R.T.; Peng, J.B.; Huang, L.L.; Jiang, G.P.; Liao, Y.J.; Sun, H.; Hu, Y.D.; Liao, X.H. Augmenter of Liver Regeneration Alleviates Renal Hypoxia-Reoxygenation Injury by Regulating Mitochondrial Dynamics in Renal Tubular Epithelial Cells. Mol. Cells 2019, 42, 893–905. [Google Scholar] [CrossRef]
- Tanaka, S.; Tanaka, T.; Nangaku, M. Hypoxia and Dysregulated Angiogenesis in Kidney Disease. Kidney Dis. 2015, 1, 80–89. [Google Scholar] [CrossRef]
- Wen, L.; Li, Y.; Li, S.; Hu, X.; Wei, Q.; Dong, Z. Glucose Metabolism in Acute Kidney Injury and Kidney Repair. Front. Med. 2021, 8, 744122. [Google Scholar] [CrossRef]
- Legouis, D.; Faivre, A.; Cippa, P.E.; de Seigneux, S. Renal gluconeogenesis: An underestimated role of the kidney in systemic glucose metabolism. Nephrol. Dial. Transplant. 2020. [Google Scholar] [CrossRef]
- Coerver, K.A.; Gray, S.M.; Barnes, J.E.; Armstrong, D.L.; McCabe, E.R. Developmental expression of hexokinase 1 and 3 in rats. Histochem. Cell Biol. 1998, 109, 75–86. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, X.; Guan, X.; Ma, X.; Chen, H.; Huang, B.; Chen, D. YAF2 exerts anti-apoptotic effect in human tumor cells in a FANK1- and phosphorylation-dependent manner. Biochem. Biophys. Res. Commun. 2021, 554, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Ferenbach, D.A.; Kluth, D.C.; Hughes, J. Hemeoxygenase-1 and renal ischaemia-reperfusion injury. Nephron Exp. Nephrol. 2010, 115, e33–e37. [Google Scholar] [CrossRef]
- Wozniak, J.; Floege, J.; Ostendorf, T.; Ludwig, A. Key metalloproteinase-mediated pathways in the kidney. Nat. Rev. Nephrol. 2021, 17, 513–527. [Google Scholar] [CrossRef] [PubMed]
- Tulluri, V.; Nemmara, V.V. Role of Antizyme Inhibitor Proteins in Cancers and Beyond. Onco Targets Ther. 2021, 14, 667–682. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Jiang, Y.; Jiang, M.; Zhang, J.; Yang, B.; She, Y.; Wang, W.; Deng, Y.; Ye, Y. Protocadherin 10 inhibits cell proliferation and induces apoptosis via regulation of DEP domain containing 1 in endometrial endometrioid carcinoma. Exp. Mol. Pathol. 2016, 100, 344–352. [Google Scholar] [CrossRef]
- Schlegel, N.; Boerner, K.; Waschke, J. Targeting desmosomal adhesion and signalling for intestinal barrier stabilization in inflammatory bowel diseases-Lessons from experimental models and patients. Acta Physiol. 2021, 231, e13492. [Google Scholar] [CrossRef]
- Dho, S.H.; Lee, K.P.; Jeong, D.; Kim, C.J.; Chung, K.S.; Kim, J.Y.; Park, B.C.; Park, S.S.; Kim, S.Y.; Kwon, K.S. GPR171 expression enhances proliferation and metastasis of lung cancer cells. Oncotarget 2016, 7, 7856–7865. [Google Scholar] [CrossRef]
- Chen, Q.I.; Cao, B.; Nan, N.; Wang, Y.U.; Zhai, X.U.; Li, Y.; Chong, T. Elevated expression of KIF18A enhances cell proliferation and predicts poor survival in human clear cell renal carcinoma. Exp. Ther. Med. 2016, 12, 377–383. [Google Scholar] [CrossRef]
- Shan, Z.G.; Sun, Z.W.; Zhao, L.Q.; Gou, Q.; Chen, Z.F.; Zhang, J.Y.; Chen, W.; Su, C.Y.; You, N.; Zhuang, Y.; et al. Upregulation of Tubulointerstitial nephritis antigen like 1 promotes gastric cancer growth and metastasis by regulating multiple matrix metallopeptidase expression. J. Gastroenterol. Hepatol. 2021, 36, 196–203. [Google Scholar] [CrossRef]
- Wang, Y.; Yao, J.; Zhu, Y.; Zhao, X.; Lv, J.; Sun, F. Knockdown of CDCA5 suppresses malignant progression of breast cancer cells by regulating PDS5A. Mol. Med. Rep. 2022, 25, 209. [Google Scholar] [CrossRef]
- Lee, K.; Gusella, G.L.; He, J.C. Epithelial proliferation and cell cycle dysregulation in kidney injury and disease. Kidney Int. 2021, 100, 67–78. [Google Scholar] [CrossRef]
- Thompson, E.M.; Stoker, A.W. A Review of DUSP26: Structure, Regulation and Relevance in Human Disease. Int. J. Mol. Sci. 2021, 22, 776. [Google Scholar] [CrossRef] [PubMed]
- Baeza-Raja, B.; Goodyear, A.; Liu, X.; Lam, K.; Yamamoto, L.; Li, Y.; Dodson, G.S.; Takeuchi, T.; Kisseleva, T.; Brenner, D.A.; et al. Pharmacological inhibition of P2RX7 ameliorates liver injury by reducing inflammation and fibrosis. PLoS ONE 2020, 15, e0234038. [Google Scholar] [CrossRef] [PubMed]
- Craciun, F.L.; Ajay, A.K.; Hoffmann, D.; Saikumar, J.; Fabian, S.L.; Bijol, V.; Humphreys, B.D.; Vaidya, V.S. Pharmacological and genetic depletion of fibrinogen protects from kidney fibrosis. Am. J. Physiol. Ren. Physiol. 2014, 307, F471–F484. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zheng, C.; Lu, Y.; Jiang, Q.; Yin, R.; Zhu, P.; Zhou, M.; Liu, Z. Urinary Fibrinogen as a Predictor of Progression of CKD. Clin. J. Am. Soc. Nephrol. 2017, 12, 1922–1929. [Google Scholar] [CrossRef] [PubMed]
- Silva, V.R.R.; Molinaro, A.; Gaudi, A.U.; Fryk, E.; Sardi, C.; Hammarlund, M.; Mjornstedt, F.; Johansson, M.E.; Becattini, B.; Jansson, P.A.; et al. Somatic ablation of IKKbeta in liver and leukocytes is not tolerated in obese mice but hepatic IKKbeta deletion improves fatty liver and insulin sensitivity. FASEB J. 2022, 36, e22512. [Google Scholar] [CrossRef]
- Traykova-Brauch, M.; Schonig, K.; Greiner, O.; Miloud, T.; Jauch, A.; Bode, M.; Felsher, D.W.; Glick, A.B.; Kwiatkowski, D.J.; Bujard, H.; et al. An efficient and versatile system for acute and chronic modulation of renal tubular function in transgenic mice. Nat. Med. 2008, 14, 979–984. [Google Scholar] [CrossRef]
- Kastner, C.; Pohl, M.; Sendeski, M.; Stange, G.; Wagner, C.A.; Jensen, B.; Patzak, A.; Bachmann, S.; Theilig, F. Effects of receptor-mediated endocytosis and tubular protein composition on volume retention in experimental glomerulonephritis. Am. J. Physiol. Ren. Physiol. 2009, 296, F902–F911. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data; Babraham Bioinformatics, Babraham Institute: Cambridge, UK, 2010. [Google Scholar]
- Krueger, F. Trim Galore. A Wrapper Tool around Cutadapt and FastQC to Consistently Apply Quality and Adapter Trimming to FastQ Files; Babraham Bioinformatics, Babraham Institute: Cambridge, UK, 2015. [Google Scholar]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, S.; Li, W. RSeQC: Quality control of RNA-seq experiments. Bioinformatics 2012, 28, 2184–2185. [Google Scholar] [CrossRef] [PubMed]
- Sayols, S.; Scherzinger, D.; Klein, H. dupRadar: A Bioconductor package for the assessment of PCR artifacts in RNA-Seq data. BMC Bioinform. 2016, 17, 428. [Google Scholar] [CrossRef] [PubMed]
- Daley, T.; Smith, A.D. Predicting the molecular complexity of sequencing libraries. Nat. Methods 2013, 10, 325–327. [Google Scholar] [CrossRef]
- Ewels, P.; Magnusson, M.; Lundin, S.; Kaller, M. MultiQC: Summarize analysis results for multiple tools and samples in a single report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Rigillo, A.; Fuchs-Baumgartinger, A.; Sabattini, S.; Skor, O.; Agnoli, C.; Schwendenwein, I.; Bettini, G.; Rutgen, B.C. Ki-67 assessment-agreeability between immunohistochemistry and flow cytometry in canine lymphoma. Vet. Comp. Oncol. 2021, 19, 551–566. [Google Scholar] [CrossRef]
- Mazzoli, A.; Sardi, C.; Breasson, L.; Theilig, F.; Becattini, B.; Solinas, G. JNK1 ablation improves pancreatic beta-cell mass and function in db/db diabetic mice without affecting insulin sensitivity and adipose tissue inflammation. FASEB Bioadv. 2021, 3, 94–107. [Google Scholar] [CrossRef]
- Theilig, F.; Kriz, W.; Jerichow, T.; Schrade, P.; Hahnel, B.; Willnow, T.; Le Hir, M.; Bachmann, S. Abrogation of protein uptake through megalin-deficient proximal tubules does not safeguard against tubulointerstitial injury. J. Am. Soc. Nephrol. 2007, 18, 1824–1834. [Google Scholar] [CrossRef]
- Theilig, F.; Enke, A.K.; Scolari, B.; Polzin, D.; Bachmann, S.; Koesters, R. Tubular deficiency of von Hippel-Lindau attenuates renal disease progression in anti-GBM glomerulonephritis. Am. J. Pathol. 2011, 179, 2177–2188. [Google Scholar] [CrossRef]
- Sachs, L. Angewandte Statistik, 7th ed.; Springer: Berlin/Heidelberg, Germany, 1992; pp. 361–362. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dahlke, E.; Engmann, T.; Anan, Y.; Häsler, R.; Solinas, G.; Theilig, F. Tubular IKKβ Deletion Alleviates Acute Ischemic Kidney Injury and Facilitates Tissue Regeneration. Int. J. Mol. Sci. 2022, 23, 10199. https://doi.org/10.3390/ijms231710199
Dahlke E, Engmann T, Anan Y, Häsler R, Solinas G, Theilig F. Tubular IKKβ Deletion Alleviates Acute Ischemic Kidney Injury and Facilitates Tissue Regeneration. International Journal of Molecular Sciences. 2022; 23(17):10199. https://doi.org/10.3390/ijms231710199
Chicago/Turabian StyleDahlke, Eileen, Toni Engmann, Yaman Anan, Robert Häsler, Giovanni Solinas, and Franziska Theilig. 2022. "Tubular IKKβ Deletion Alleviates Acute Ischemic Kidney Injury and Facilitates Tissue Regeneration" International Journal of Molecular Sciences 23, no. 17: 10199. https://doi.org/10.3390/ijms231710199
APA StyleDahlke, E., Engmann, T., Anan, Y., Häsler, R., Solinas, G., & Theilig, F. (2022). Tubular IKKβ Deletion Alleviates Acute Ischemic Kidney Injury and Facilitates Tissue Regeneration. International Journal of Molecular Sciences, 23(17), 10199. https://doi.org/10.3390/ijms231710199