
Proteomics Profiling of Osteoporosis and Osteopenia Patients and Associated Network Analysis


 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Clinical Characteristics and Demographics of Study Population
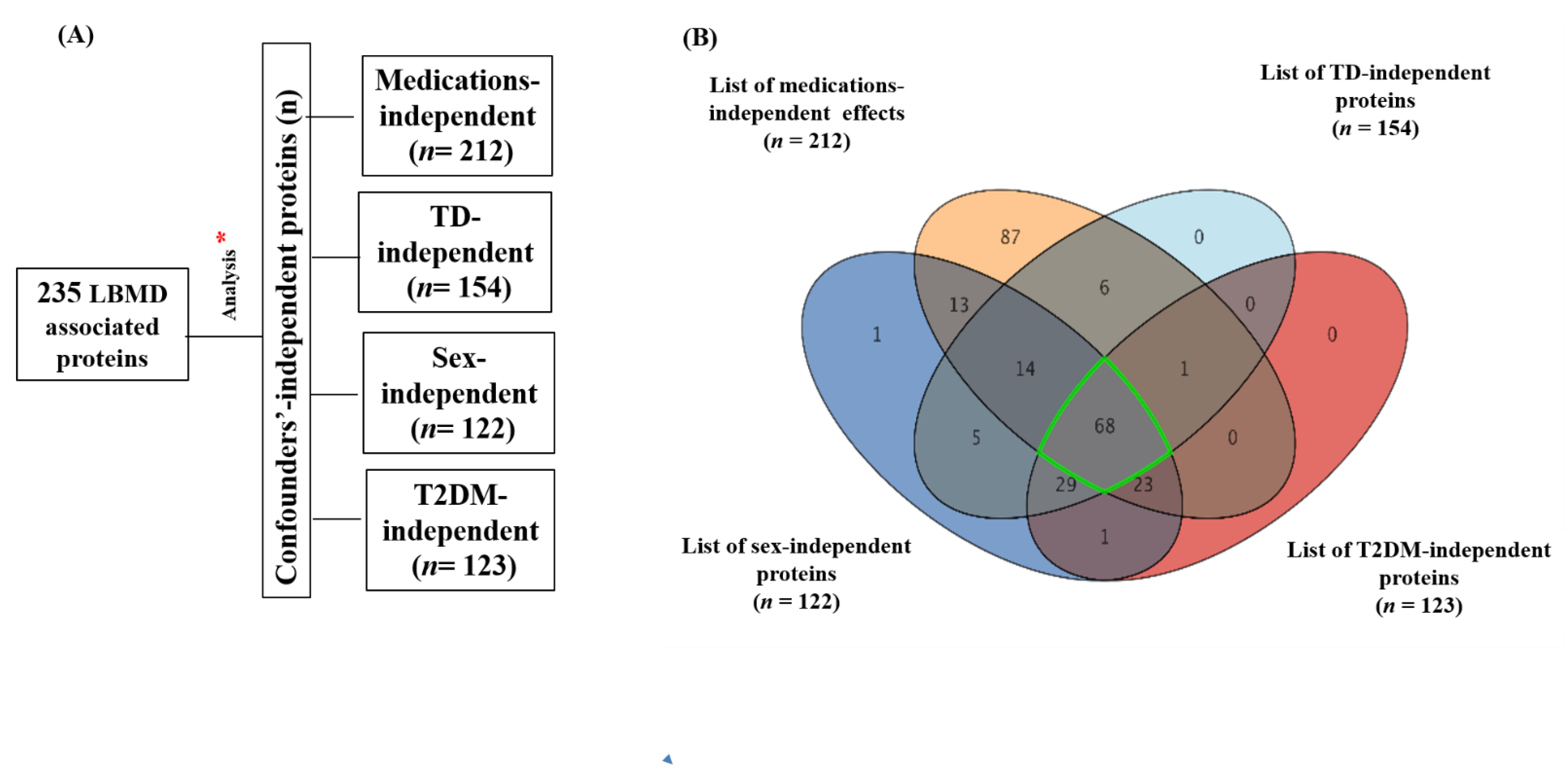
2.2. Results of Overall Proteomics Analysis and Exclusion of Confounder-Associated Proteins
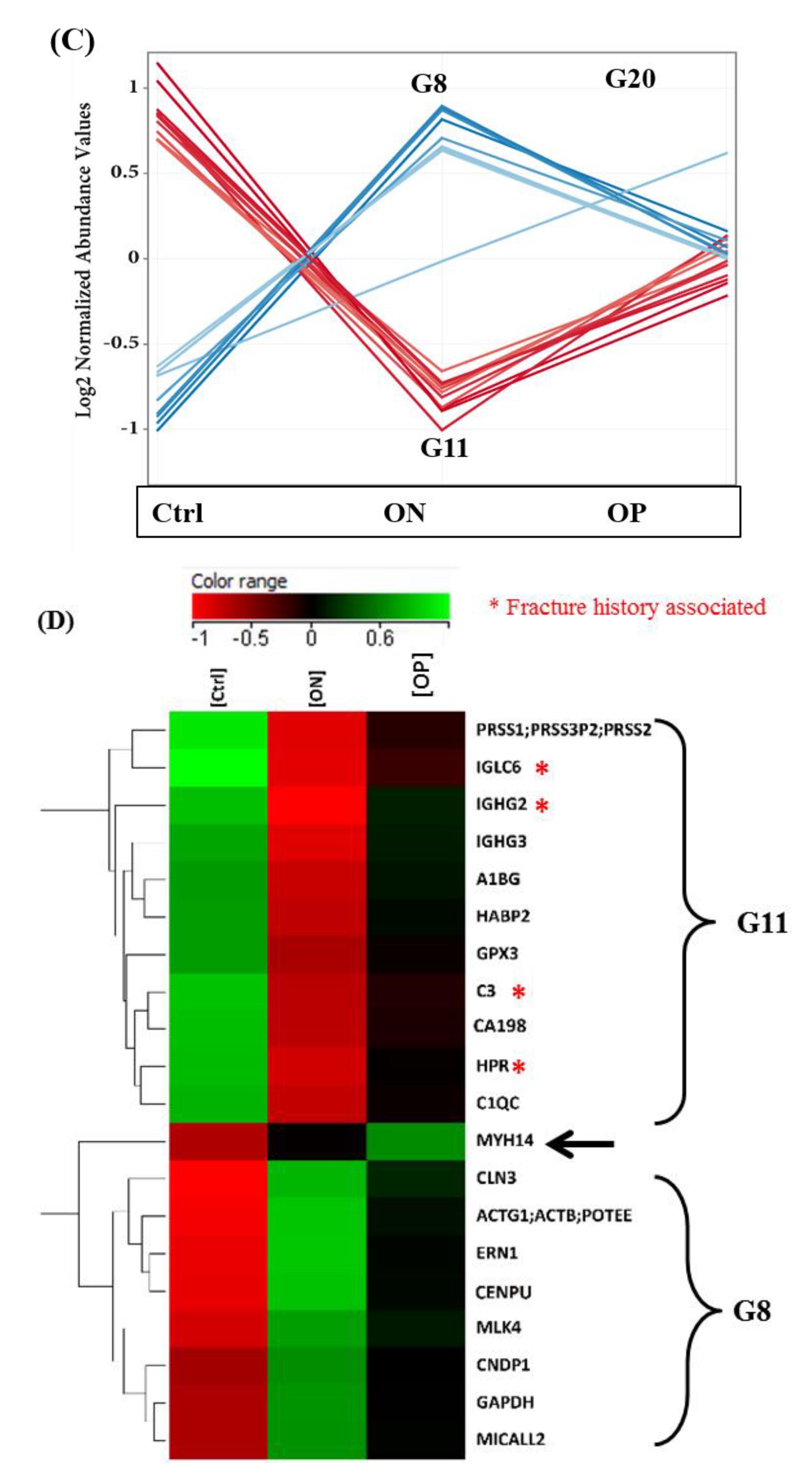
2.3. Proteomics Profiles of Control, ON, and OP Groups
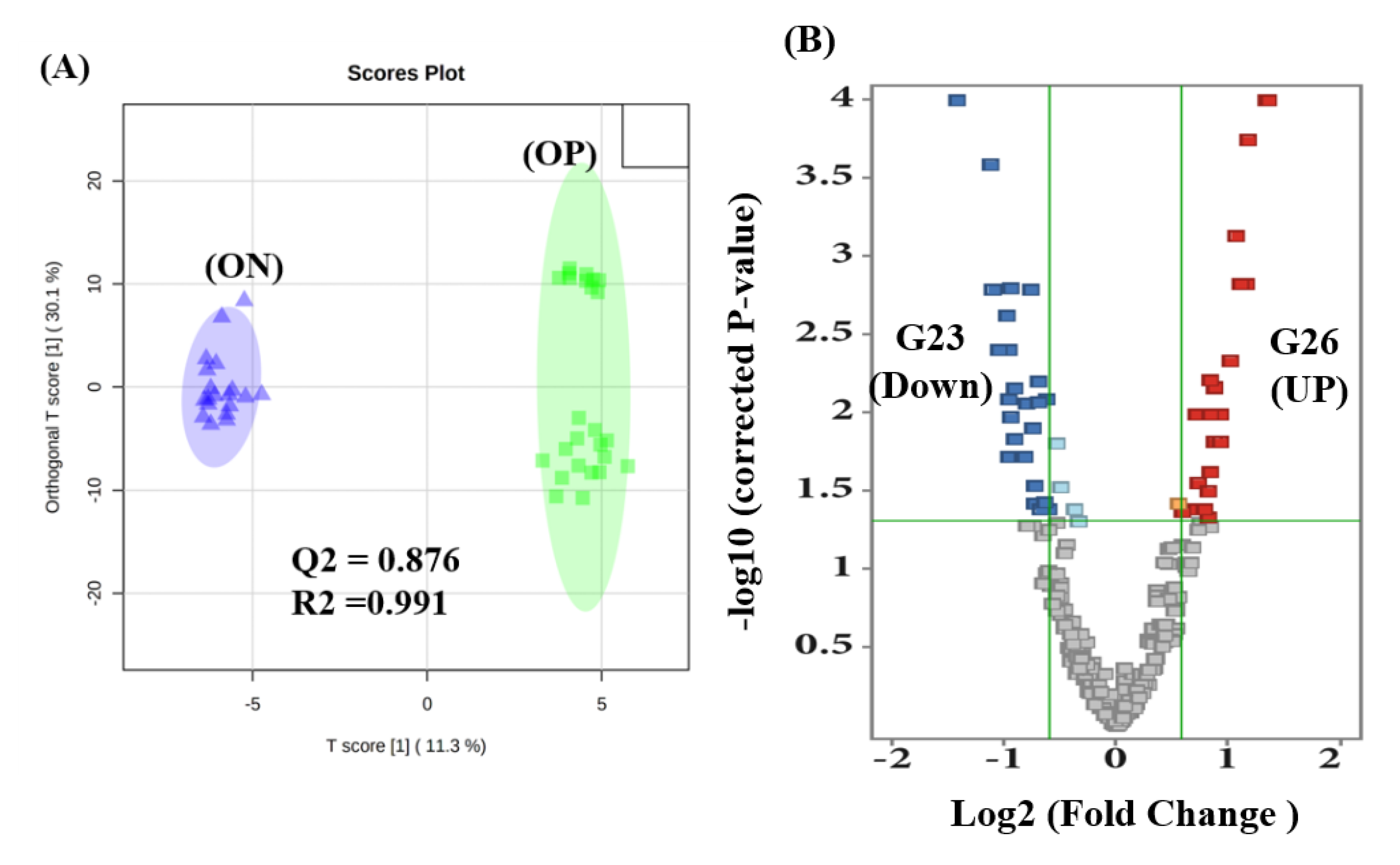
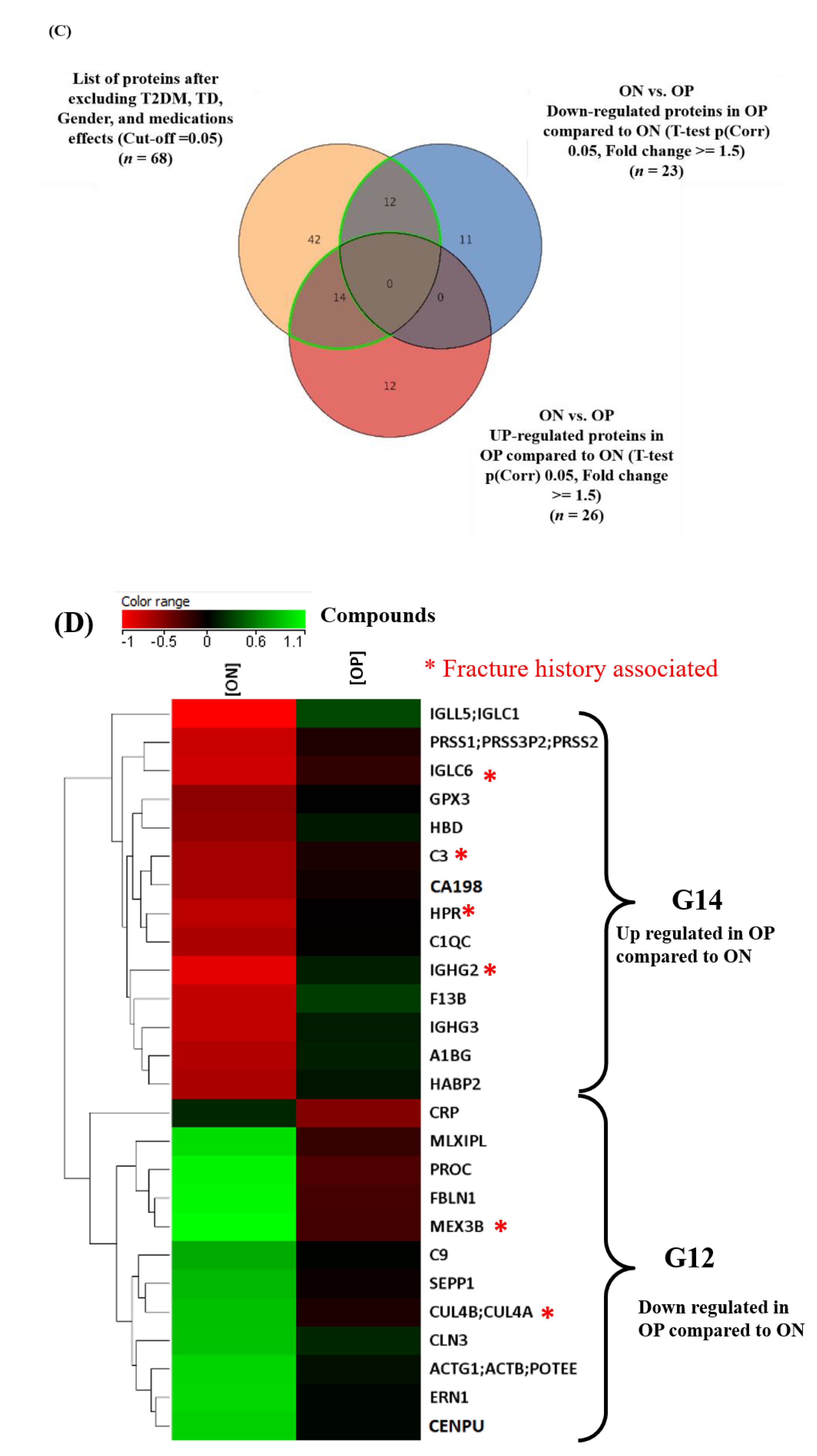
2.4. Proteomics Profile between ON and OP Groups
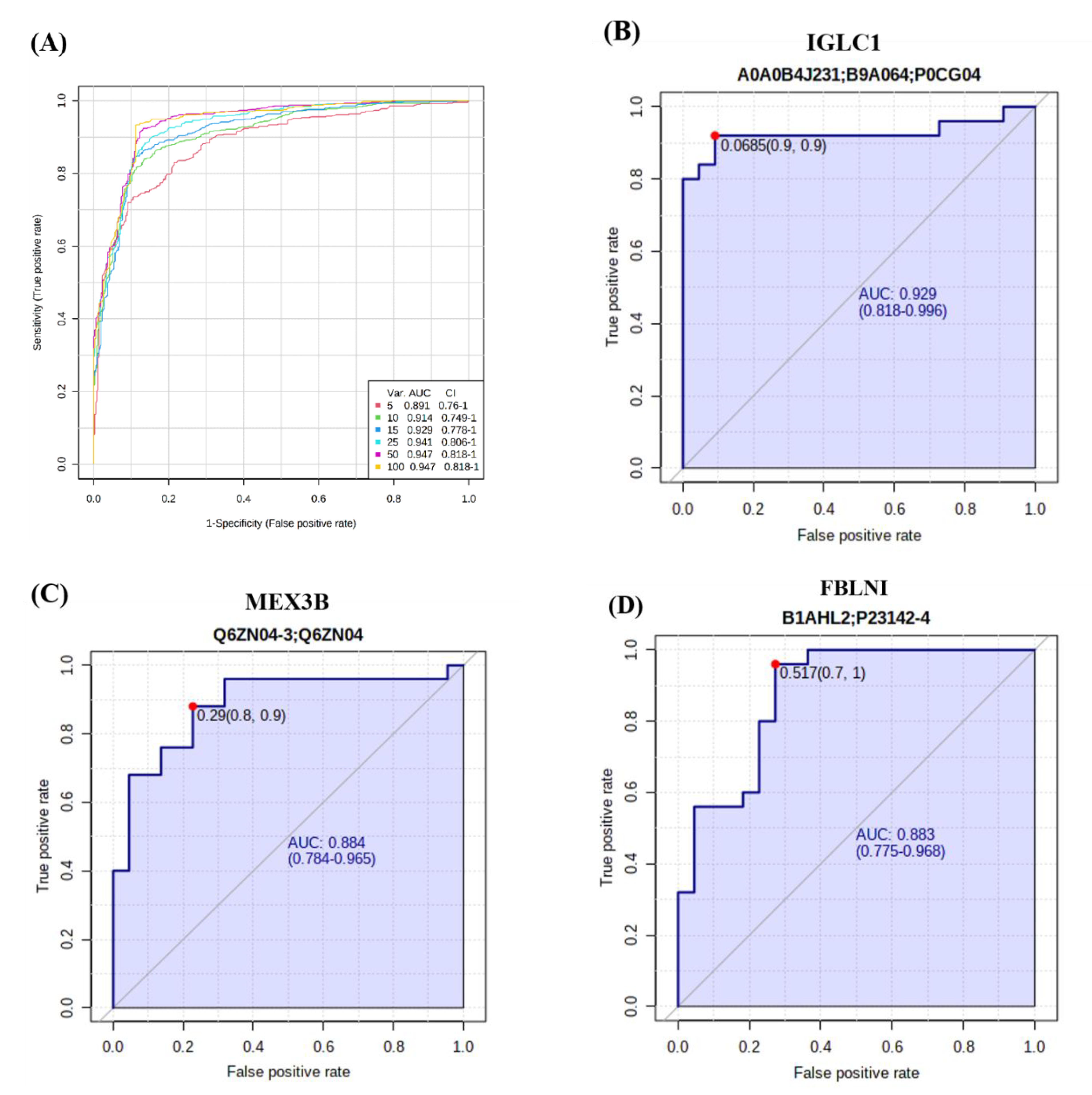
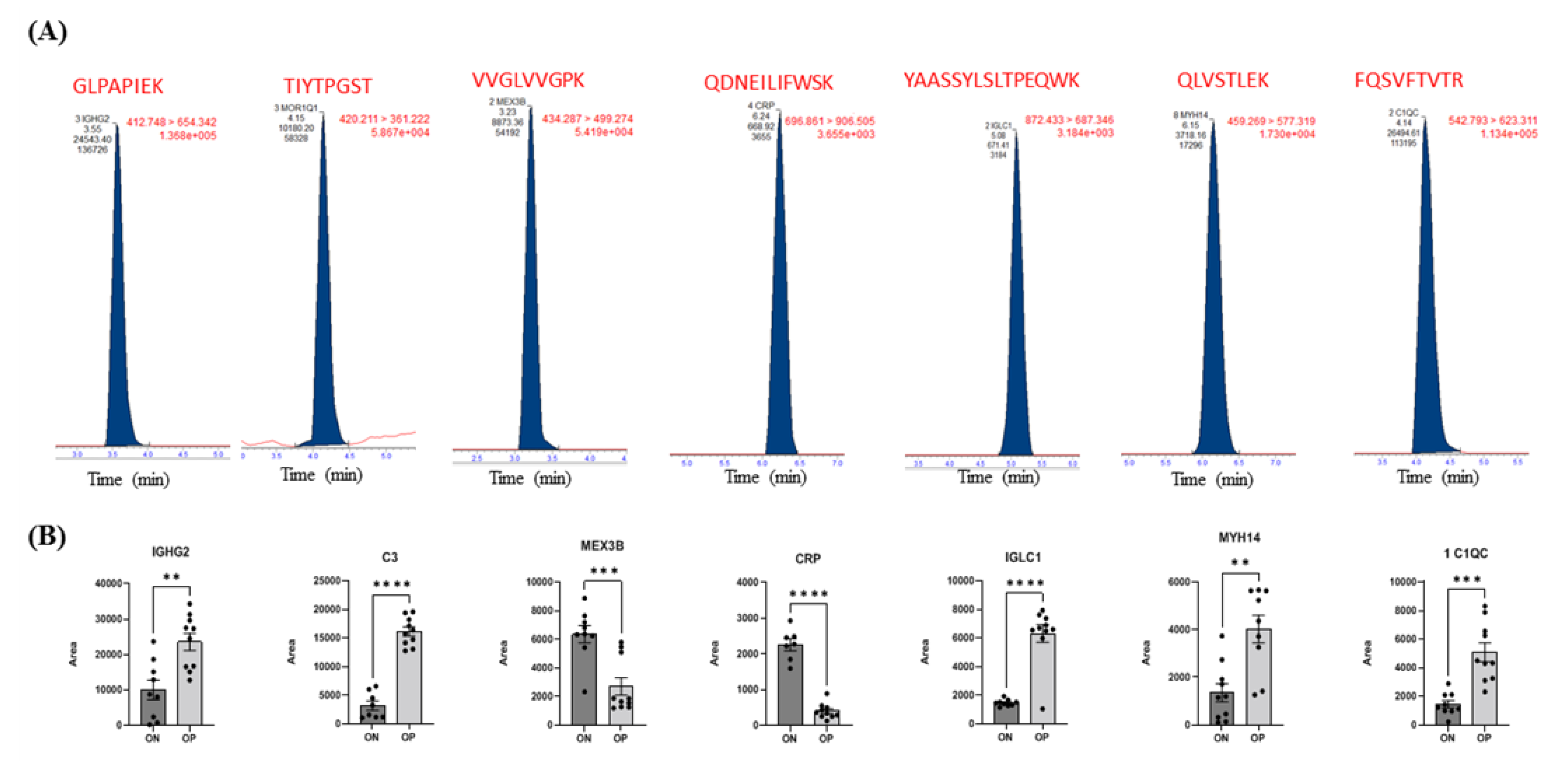
2.5. Evaluation of Biomarkers between ON and OP
2.6. Network Module Analysis and Biological Pathways Related to ON and OP
3. Discussion
3.1. Proteomics Profiles of Control, ON, and OP Groups
3.2. Fracture History-Associated Proteins in Control, ON, and OP Groups
3.3. Comparison of Proteomics Profiles between ON and OP Groups: Role of the Immune System
3.4. Common and Significant Proteins Biomarkers and Pathways in ON and OP
4. Materials and Methods
4.1. Study Population and Recruitment
4.2. Ethical Approval
4.3. Proteomics Analysis
4.3.1. Protein Digestion and Sample Preparation
4.3.2. Nano LC-MS/MS Analysis
4.4. LC-MS/MS—Multiple Reaction Monitoring (MRM)
4.5. Database Search
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, H.; Recker, R.; Lee, W.N.; Xiao, G.G. Proteomics in bone research. Expert Rev. Proteom. 2010, 7, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Swank, K.R.; Furness, J.E.; Baker, E.A.; Gehrke, C.K.; Biebelhausen, S.P.; Baker, K.C. Metabolomic Profiling in the Character-ization of Degenerative Bone and Joint Diseases. Metabolites 2020, 10, 223. [Google Scholar] [CrossRef] [PubMed]
- Tu, K.N.; Lie, J.D.; Wan, C.K.V.; Cameron, M.; Austel, A.G.; Nguyen, J.K.; Van, K.; Hyun, D. Osteoporosis: A Review of Treatment Options. Pharm. Ther. 2018, 43, 92–104. [Google Scholar]
- Sozen, T.; Ozisik, L.; Basaran, N.C. An overview and management of osteoporosis. Eur. J. Rheumatol. 2017, 4, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Malkawi, A.K.; Masood, A.; Shinwari, Z.; Jacob, M.; Benabdelkamel, H.; Matic, G.; Almuhanna, F.; Dasouki, M.; Alaiya, A.A.; Rahman, A.M.A. Proteomic Analysis of Morphologically Changed Tissues after Prolonged Dexamethasone Treatment. Int. J. Mol. Sci. 2019, 20, 3122. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Yang, R.; Wei, L.; Liu, J.; Yang, Y.; Shao, F.; Ma, W.; Li, T.; Wang, Y.; Guo, T. Prevalence of osteoporosis and related lifestyle and metabolic factors of postmenopausal women and elderly men: A cross-sectional study in Gansu province, Northwestern of China. Medicine 2017, 96, e8294. [Google Scholar] [CrossRef]
- Moh, P. National Plan for Osteoporosis Prevention and Management in KSA. 2019. Available online: https://www.moh.gov.sa/en/Ministry/MediaCenter/Publications/Documents/NPOPM-2018.pdf (accessed on 8 August 2020).
- Lems, W.F.; Raterman, H.G. Critical issues and current challenges in osteoporosis and fracture prevention. An overview of unmet needs. Ther. Adv. Musculoskelet. Dis. 2017, 9, 299–316. [Google Scholar] [CrossRef]
- Al-Saleh, Y.; Sulimani, R.; Sabico, S.; Raef, H.; Fouda, M.; Alshahrani, F.; Al Shaker, M.; Al Wahabi, B.; Sadat-Ali, M.; Al Rayes, H.; et al. 2015 Guidelines for Osteoporosis in Saudi Arabia: Recommendations from the Saudi Osteoporosis Society. Ann. Saudi Med. 2015, 35, 1–12. [Google Scholar] [CrossRef]
- NIH Consensus Development Panel on Osteoporosis Prevention, Diagnosis, and Therapy. Osteoporosis prevention, diagnosis, and therapy. JAMA 2001, 285, 785–795. [Google Scholar] [CrossRef]
- Willson, T.; Nelson, S.D.; Newbold, J.; Nelson, R.E.; LaFleur, J. The clinical epidemiology of male osteoporosis: A review of the recent literature. Clin. Epidemiol. 2015, 7, 65–76. [Google Scholar]
- Veiga Silva, A.C.; da Rosa, M.I.; Fernandes, B.; Lumertz, S.; Diniz, R.M.; dos Reis Damiani, M.E. Factors associated with osteopenia and osteoporosis in women undergoing bone mineral density test. Rev. Bras. De Reumatol. 2015, 55, 223–228. [Google Scholar] [CrossRef]
- Tella, S.H.; Gallagher, J.C. Prevention and treatment of postmenopausal osteoporosis. J. Steroid Biochem. Mol. biology 2014, 142, 155–170. [Google Scholar] [CrossRef] [Green Version]
- Aleidi, S.M.; Alnehmi, E.A.; Alshaker, M.; Masood, A.; Benabdelkamel, H.; Al-Ansari, M.M.; Abdel Rahman, A.M. A Distinctive Human Metabolomics Alteration Associated with Osteopenic and Osteoporotic Patients. Metabolites 2021, 11, 628. [Google Scholar] [CrossRef]
- Zhu, W.; Shen, H.; Zhang, J.G.; Zhang, L.; Zeng, Y.; Huang, H.L.; Zhao, Y.C.; He, H.; Zhou, Y.; Wu, K.H.; et al. Cytosolic proteome profiling of monocytes for male osteoporosis. Osteoporos. Int. 2017, 28, 1035–1046. [Google Scholar] [CrossRef]
- Deng, F.-Y.; Liu, Y.-Z.; Li, L.-M.; Jiang, C.; Wu, S.; Chen, Y.; Jiang, H.; Yang, F.; Xiong, J.-X.; Xiao, P.; et al. Proteomic analysis of circulating monocytes in Chinese premenopausal females with extremely discordant bone mineral density. Proteomics 2008, 8, 4259–4272. [Google Scholar] [CrossRef]
- Lv, M.; Cui, C.; Chen, P.; Li, Z. Identification of osteoporosis markers through bioinformatic functional analysis of serum proteome. Medicine 2020, 99, e22172. [Google Scholar] [CrossRef]
- Huang, D.; Wang, Y.; Lv, J.; Yan, Y.; Hu, Y.; Liu, C.; Zhang, F.; Wang, J.; Hao, D. Proteomic profiling analysis of postmeno-pausal osteoporosis and osteopenia identifies potential proteins associated with low bone mineral density. Peer J. 2020, 8, e9009. [Google Scholar] [CrossRef]
- Zhang, L.L.; Li, C.W.; Liu, K.; Liu, Z.; Liang, B.C.; Yang, Y.R.; Shi, X.L. Discovery and Identification of Serum Succinyl-Proteome for Postmenopausal Women with Osteoporosis and Osteopenia. Orthop. Surg. 2019, 11, 784–793. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, Y.-Z.; Zeng, Y.; Zhu, W.; Zhao, Y.-C.; Zhang, J.-G.; Zhu, J.-Q.; He, H.; Shen, H.; Tian, Q.; et al. Network-based proteomic analysis for postmenopausal osteoporosis in Caucasian females. Proteomics 2016, 16, 12–28. [Google Scholar] [CrossRef]
- Huo, C.; Li, Y.; Qiao, Z.; Shang, Z.; Cao, C.; Hong, Y.; Xiao, H. Comparative proteomics analysis of microvesicles in human serum for the evaluation of osteoporosis. Electrophoresis 2019, 40, 1839–1847. [Google Scholar] [CrossRef]
- Nielson, C.M.; Wiedrick, J.; Shen, J.; Jacobs, J.; Baker, E.S.; Baraff, A.; Piehowski, P.; Lee, C.G.; Baratt, A.; Petyuk, V.; et al. Identification of Hip BMD Loss and Fracture Risk Markers Through Population-Based Serum Proteomics. J. Bone Miner. Res. 2017, 32, 1559–1567. [Google Scholar] [CrossRef] [PubMed]
- Picke, A.K.; Campbell, G.; Napoli, N.; Hofbauer, L.C.; Rauner, M. Update on the impact of type 2 diabetes mellitus on bone metabolism and material properties. Endocr. Connect. 2019, 8, R55–R70. [Google Scholar] [CrossRef] [PubMed]
- Delitala, A.P.; Scuteri, A.; Doria, C. Thyroid Hormone Diseases and Osteoporosis. J. Clin. Med. 2020, 9, 1034. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Lovre, D.; Mauvais-Jarvis, F. The effect of selective estrogen receptor modulators on type 2 diabetes onset in women: Basic and clinical insights. Diabetes its Complicat. 2017, 31, 773–779. [Google Scholar] [CrossRef] [PubMed]
- Golomb, E.; Ma, X.; Jana, S.S.; Preston, Y.A.; Kawamoto, S.; Shoham, N.G.; Goldin, E.; Conti, M.A.; Sellers, J.R.; Adelstein, R. Identification and characterization of nonmuscle myosin II-C, a new member of the myosin II family. J. Biol. Chem. 2004, 279, 2800–2808. [Google Scholar] [CrossRef]
- Tasca, A.; Astleford, K.; Lederman, A.; Jensen, E.D.; Lee, B.S.; Gopalakrishnan, R.; Mansky, K.C. Regulation of Osteoclast Differentiation by Myosin X. Sci. Rep. 2017, 7, 7603. [Google Scholar] [CrossRef] [PubMed]
- McMichael, B.K.; Cheney, R.E.; Lee, B.S. Myosin X regulates sealing zone patterning in osteoclasts through linkage of podosomes and microtubules. J. Biol. Chem. 2010, 285, 9506–9515. [Google Scholar] [CrossRef]
- Mumtaz, M. An approach to the patient with osteoporosis. Malays. J. Med. Sci. 2001, 8, 11–19. [Google Scholar]
- Mori, G.; D’Amelio, P.; Faccio, R.; Brunetti, G. The Interplay between the bone and the immune system. Clin. Dev. Immunol. 2013, 2013, 720504. [Google Scholar] [CrossRef]
- Guder, C.; Gravius, S.; Burger, C.; Wirtz, D.C.; Schildberg, F.A. Osteoimmunology: A Current Update of the Interplay Between Bone and the Immune System. Front. Immunol. 2020, 11, 58. [Google Scholar] [CrossRef]
- Walsh, M.C.; Kim, N.; Kadono, Y.; Rho, J.; Lee, S.Y.; Lorenzo, J.; Choi, Y. Osteoimmunology: Interplay between the immune system and bone metabolism. Annu. Rev. Immunol. 2006, 24, 33–63. [Google Scholar] [CrossRef] [PubMed]
- Pineda, B.; Serna, E.; Laguna-Fernández, A.; Noguera, I.; Panach, L.; Hermenegildo, C.; Tarín, J.J.; Cano, A.; García-Pérez, M. Gene expression profile induced by ovariectomy in bone marrow of mice: A functional approach to identify new candidate genes associated to osteoporosis risk in women. Bone 2014, 65, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Eghbali-Fatourechi, G.; Khosla, S.; Sanyal, A.; Boyle, W.J.; Lacey, D.L.; Riggs, B.L. Role of RANK ligand in mediating increased bone resorption in early postmenopausal women. J. Clin. Investig. 2003, 111, 1221–1230. [Google Scholar] [CrossRef] [Green Version]
- Mödinger, Y.; Löffler, B.; Huber-Lang, M.; Ignatius, A. Complement involvement in bone homeostasis and bone disorders. Semin. Immunol. 2018, 37, 53–65. [Google Scholar] [CrossRef]
- Elsner, J.; Oppermann, M.; Czech, W.; Kapp, A. C3a activates the respiratory burst in human polymorphonuclear neu-trophilic leukocytes via pertussis toxin-sensitive G-proteins. Blood 1994, 83, 3324–3331. [Google Scholar] [CrossRef]
- Teo, B.H.; Bobryshev, Y.V.; Teh, B.K.; Wong, S.H.; Lu, J. Complement C1q production by osteoclasts and its regulation of osteoclast development. Biochem. J. 2012, 447, 229–237. [Google Scholar] [CrossRef]
- Andrades, J.A.; Nimni, M.E.; Becerra, J.; Eisenstein, R.; Davis, M.; Sorgente, N. Complement proteins are present in developing endochondral bone and may mediate cartilage cell death and vascularization. Exp. Cell Res. 1996, 227, 208–213. [Google Scholar] [CrossRef]
- Cooley, M.A.; Kern, C.B.; Fresco, V.M.; Wessels, A.; Thompson, R.P.; McQuinn, T.C.; Twal, W.O.; Mjaatvedt, C.H.; Drake, C.J.; Argraves, W.S. Fibulin-1 is required for morphogenesis of neural crest-derived structures. Dev. Biol. 2008, 319, 336–345. [Google Scholar] [CrossRef]
- Goel, D.; Vohora, D. Liver X receptors and skeleton: Current state-of-knowledge. Bone 2021, 144, 115807. [Google Scholar] [CrossRef]
- Menéndez-Gutiérrez, M.P.; Rőszer, T.; Fuentes, L.; Núñez, V.; Escolano, A.; Redondo, J.M.; De Clerck, N.; Metzger, D.; Valledor, A.F.; Ricote, M. Retinoid X receptors orchestrate osteoclast differentiation and postnatal bone remodeling. J. Clin. Investig. 2015, 125, 809–823. [Google Scholar] [CrossRef]
- Cho, S.W.; An, J.H.; Park, H.; Yang, J.Y.; Choi, H.J.; Kim, S.W.; Park, Y.J.; Kim, S.Y.; Yim, M.; Baek, W.Y.; et al. Positive regulation of osteogenesis by bile acid through FXR. J. Bone Miner. Res. 2013, 28, 2109–2121. [Google Scholar] [CrossRef] [PubMed]
- Benabdelkamel, H.; Alamri, H.; Okla, M.; Masood, A.; Jabar, M.A.; Alanazi, I.O.; Alfadda, A.A.; Nizami, I.; Dasouki, M.; Rahman, A.M.A. Serum-Based Proteomics Profiling in Adult Patients with Cystic Fibrosis. Int. J. Mol. Sci. 2020, 21, 7415. [Google Scholar] [CrossRef] [PubMed]
- Galal, M.A.; Jabar, M.A.; Zhra, M.; Rahman, A.M.A.; Aljada, A. Absolute quantification of senescence mediators in cells using multiple reaction monitoring liquid chromatography-Tandem mass spectrometry. Anal. Chim. Acta 2021, 1184, 339009. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ctrl | ON | OP | ||||
|---|---|---|---|---|---|---|
| Total n (%) | 22 (31.88) | 22 (31.88) | 25 (36.23) | |||
| Parameters | Mean | SEM | Mean | SEM | Mean | SEM |
| Age (years) | 54.82 | 1.03 | 64.64 § | 1.72 | 66.16 § | 1.78 |
| Gender (F/M) | (13/9) | - | (15/7) | - | (24/1) | - |
| Menopause * (Yes/No) | (13/0) | - | (14/1) | - | (24/0) | - |
| Weight (kg) | 85.13 | 3.63 | 74.21 | 3.88 | 69.23 § | 2.86 |
| Height (cm) | 162.22 | 0.02 | 157.11 | 0.021 | 150.68 § ‡ | 0.01 |
| BMI (kg/m2) | 32.21 | 1.1 | 30.38 | 1.84 | 30.70 | 1.4 |
| Lumbar t score | 0.29 | 0.24 | −1.25 § ‡ | 0.21 | −2.62 § | 0.12 |
| Femoral t score | 0.34 | 0.29 | −1.51 § ‡ | 0.14 | −1.93 § | 0.13 |
| FBG (mmol/L) | 10.2 | 1.16 | 6.08 § | 0.39 | 5.87 § | 0.41 |
| HDL (mmol/L) | 1.00 | 0.80 | 1.47 § | 0.12 | 1.42 § | 0.09 |
| TG (mmol/L) | 1.85 | 0.15 | 1.23 § | 0.11 | 1.127 § | 0.08 |
| Cholesterol (mmol/L) | 5.51 | 0.23 | 4.47 § | 0.19 | 4.27 § | 0.29 |
| Calcium (mmol/L) | 2.24 | 0.026 | 2.37 § | 0.025 | 2.33 § | 0.02 |
| Albumin (g/L) | 37.65 | 1.14 | 41.98 § | 2.0 | 42.75 § | 0.86 |
| Vitamin D 25 hydroxy (nmol/L) | 68.32 | 7.39 | 77.64 | 3.3 | 86.57 | 6.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Ansari, M.M.; Aleidi, S.M.; Masood, A.; Alnehmi, E.A.; Abdel Jabar, M.; Almogren, M.; Alshaker, M.; Benabdelkamel, H.; Abdel Rahman, A.M. Proteomics Profiling of Osteoporosis and Osteopenia Patients and Associated Network Analysis. Int. J. Mol. Sci. 2022, 23, 10200. https://doi.org/10.3390/ijms231710200
Al-Ansari MM, Aleidi SM, Masood A, Alnehmi EA, Abdel Jabar M, Almogren M, Alshaker M, Benabdelkamel H, Abdel Rahman AM. Proteomics Profiling of Osteoporosis and Osteopenia Patients and Associated Network Analysis. International Journal of Molecular Sciences. 2022; 23(17):10200. https://doi.org/10.3390/ijms231710200
Chicago/Turabian StyleAl-Ansari, Mysoon M., Shereen M. Aleidi, Afshan Masood, Eman A. Alnehmi, Mai Abdel Jabar, Maha Almogren, Mohammed Alshaker, Hicham Benabdelkamel, and Anas M. Abdel Rahman. 2022. "Proteomics Profiling of Osteoporosis and Osteopenia Patients and Associated Network Analysis" International Journal of Molecular Sciences 23, no. 17: 10200. https://doi.org/10.3390/ijms231710200
APA StyleAl-Ansari, M. M., Aleidi, S. M., Masood, A., Alnehmi, E. A., Abdel Jabar, M., Almogren, M., Alshaker, M., Benabdelkamel, H., & Abdel Rahman, A. M. (2022). Proteomics Profiling of Osteoporosis and Osteopenia Patients and Associated Network Analysis. International Journal of Molecular Sciences, 23(17), 10200. https://doi.org/10.3390/ijms231710200