
Lunasin as a Promising Plant-Derived Peptide for Cancer Therapy

 ,
,  and
and
Abstract
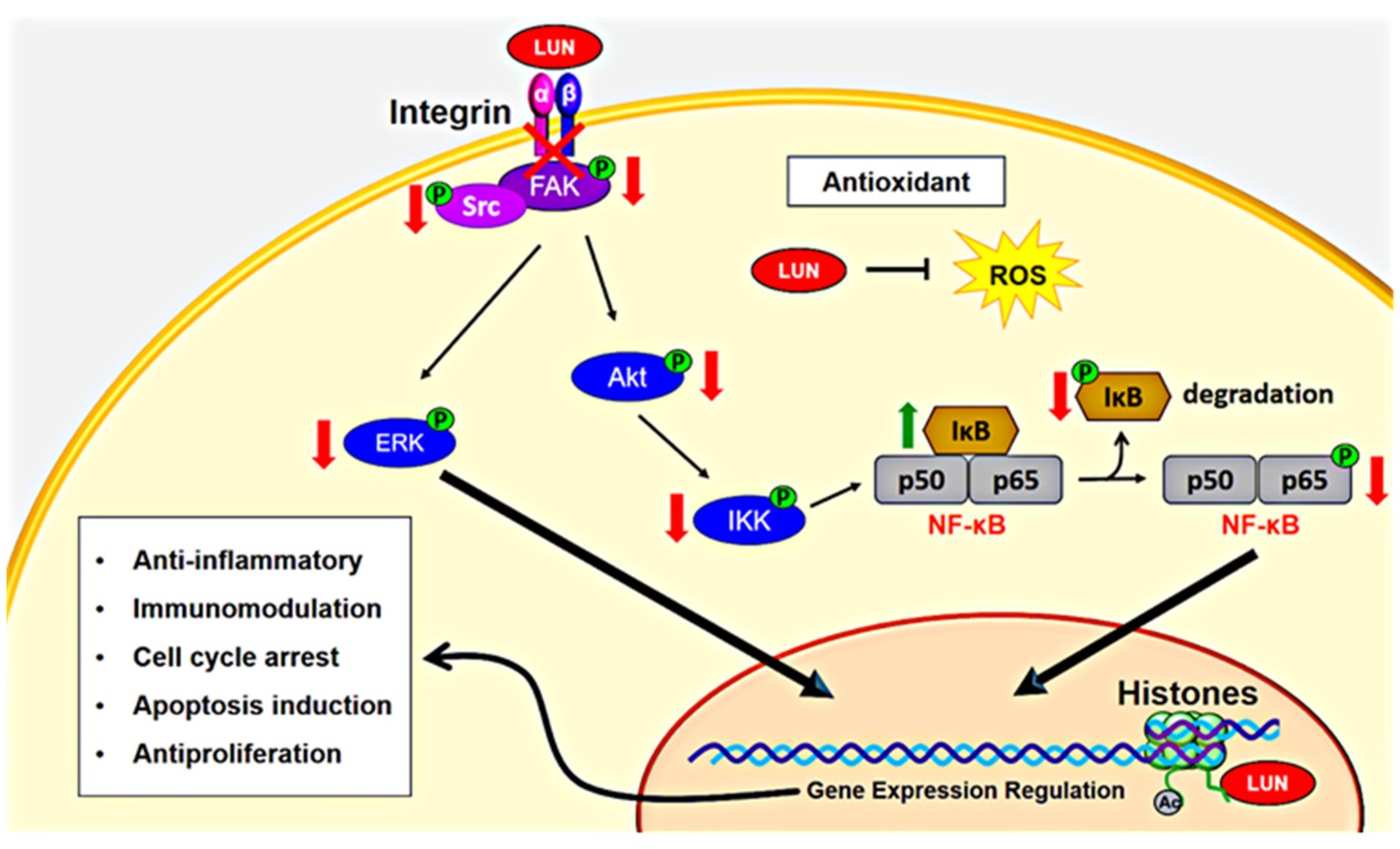
:1. Introduction
2. Lunasin Structure
2.1. Structure-Activity Relationship
2.2. Physicochemical Characterization
2.2.1. Primary Structure and Post-Translational Modifications
2.2.2. Secondary Structure
2.2.3. Tertiary and Quaternary Structures
2.2.4. Stability
3. Pharmacokinetics and Safety
4. Mechanisms of Action against Cancer
4.1. Molecular Partners
4.2. Cell Cycle and Apoptosis Regulation
4.3. Antioxidant, Anti-Inflammatory, and Immunomodulatory Activities
5. Lunasin Products and Potential Combination Therapeutics
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Stewart, B.W.; Kleihues, P. World Cancer Report; IARC Press: Lyon, France, 2003. [Google Scholar]
- Chakraborty, S.; Rahman, T. The difficulties in cancer treatment. Ecancermedicalscience 2012, 6, ed16. [Google Scholar] [PubMed]
- Ma, X.; Wu, C.; Wang, W.; Li, X. Peptides from plants: A new source for antitumor drug research. Asian J. Trad. Med. 2006, 1, 85–90. [Google Scholar]
- Galvez, A.F.; de Lumen, B.O. A soybean cDNA encoding a chromatin-binding peptide inhibits mitosis of mammalian cells. Nat. Biotechnol. 1999, 17, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Odani, S.; Koide, T.; Ono, T. Amino acid sequence of a soybean (Glycine max) seed polypeptide having a poly (L-aspartic acid) structure. J. Biol. Chem. 1987, 262, 10502–10505. [Google Scholar] [CrossRef]
- Chang, J.C. Cancer stem cells: Role in tumor growth, recurrence, metastasis, and treatment resistance. Medicine 2016, 95 (Suppl. 1), S20–S25. [Google Scholar] [CrossRef]
- D’Addio, S.M.; Bothe, J.R.; Neri, C.; Walsh, P.L.; Zhang, J.; Pierson, E.; Mao, Y.; Gindy, M.; Leone, A.; Templeton, A.C. New and evolving techniques for the characterization of peptide therapeutics. J. Pharm. Sci. 2016, 105, 2989–3006. [Google Scholar] [CrossRef]
- Mitra, M.S.; DeMarco, S.; Holub, B.; Thiruneelakantapillai, L.; Thackaberry, E.A. Development of peptide therapeutics: A nonclinical safety assessment perspective. Regul. Toxicol. Pharmacol. 2020, 117, 104766. [Google Scholar] [CrossRef]
- Wu, L. Regulatory considerations for peptide therapeutics. In Peptide Therapeutics: Strategy and Tactics for Chemistry, Manufacturing, and Controls; Srivastava, V., Ed.; The Royal Society of Chemistry: London, UK, 2019; pp. 1–30. [Google Scholar]
- Hernández-Ledesma, B.; Hsieh, C.-C.; de Lumen, B.O. Lunasin, a novel seed peptide for cancer prevention. Peptides 2009, 30, 426–430. [Google Scholar] [CrossRef]
- Jones, G.; Srivastava, A. Understanding Lunasin’s biology and potential as a cancer therapeutic by utilizing Drosophila genetics. Exp. Biol. Med. 2014, 239, 519–528. [Google Scholar] [CrossRef]
- Hernández-Ledesma, B.; Hsieh, C.-C.; de Lumen, B.O. Relationship between lunasin’s sequence and its inhibitory activity of histones H3 and H4 acetylation. Mol. Nutr. Food Res. 2011, 55, 989–998. [Google Scholar] [CrossRef]
- Galvez, A.F.; Chen, N.; Macasieb, J.; de Lumen, B.O. Chemopreventive property of a soybean peptide (lunasin) that binds to deacetylated histones and inhibits acetylation. Cancer Res. 2001, 61, 7473–7478. [Google Scholar]
- Hernández-Ledesma, B.; Hsieh, C.-C.; de Lumen, B.O. Antioxidant and anti-inflammatory properties of cancer preventive peptide lunasin in RAW 264.7 macrophages. Biochem. Biophys. Res. Commun. 2009, 390, 803–808. [Google Scholar] [CrossRef]
- Lam, Y.; Galvez, A.; de Lumen, B.O. Lunasin™ suppresses E1A-mediated transformation of mammalian cells but does not inhibit growth of immortalized and established cancer cell lines. Nutr. Cancer 2003, 47, 88–94. [Google Scholar] [CrossRef]
- Indiano-Romacho, P.; Fernández-Tomé, S.; Amigo, L.; Hernández-Ledesma, B. Multifunctionality of lunasin and peptides re-leased during its simulated gastrointestinal digestion. Food Res. Int. 2019, 125, 108513. [Google Scholar] [CrossRef]
- Chang, H.C.; Lewis, D.; Tung, C.Y.; Han, L.; Henriquez, S.M.; Voiles, L.; Lupov, I.P.; Pelloso, D.; Sinn, A.L.; Pollok, K.E.; et al. Soypeptide lunasin in cytokine immunotherapy for lymphoma. Cancer Immunol. Immunother. 2014, 63, 283–295. [Google Scholar] [CrossRef]
- Wang, F.; Li, Y.; Shen, Y.; Wang, A.; Wang, S.; Xie, T. The functions and applications of RGD in tumor therapy and tissue engineering. Int. J. Mol. Sci. 2013, 14, 13447–13462. [Google Scholar] [CrossRef]
- Nieberler, M.; Reuning, U.; Reichart, F.; Notni, J.; Wester, H.J.; Schwaiger, M.; Weinmüller, M.; Räder, A.; Steiger, K.; Kessler, H. Exploring the role of RGD-recognizing integrins in cancer. Cancers 2017, 9, 116. [Google Scholar] [CrossRef]
- Shidal, C.; Inaba, J.I.; Yaddanapudi, K.; Davis, K.R. The soy-derived peptide Lunasin inhibits invasive potential of melanoma initiating cells. Oncotarget 2017, 8, 25525. [Google Scholar] [CrossRef]
- Seber, L.E.; Barnett, B.W.; McConnell, E.J.; Hume, S.D.; Cai, J.; Boles, K.; Davis, K.R. Scalable purification and characterization of the anticancer lunasin peptide from soybean. PLoS ONE 2012, 7, e35409. [Google Scholar] [CrossRef]
- Serra, A.; Gallart-Palau, X.; See-Toh, R.S.; Hemu, X.; Tam, J.P.; Sze, S.K. Commercial processed soy-based food product contains glycated and glycoxidated lunasin proteoforms. Sci. Rep. 2016, 6, 1–12. [Google Scholar]
- Alves de Souza, S.M.; Fernandes, T.V.A.; Kalume, D.E.; Lima, L.M.T.; Pascutti, P.G.; de Souza, T.L.F. Physicochemical and structural properties of lunasin revealed by spectroscopic, chromatographic and molecular dynamics approaches. Biochim. Biophys. Acta Proteins Proteom. 2020, 1868, 140440. [Google Scholar] [CrossRef]
- Böttger, R.; Hoffmann, R.; Knappe, D. Differential stability of therapeutic peptides with different proteolytic cleavage sites in blood, plasma and serum. PLoS ONE 2017, 12, e0178943. [Google Scholar]
- Dia, V.P.; Torres, S.; de Lumen, B.O.; Erdman, J.W., Jr.; de Mejia, E.G. Presence of lunasin in plasma of men after soy protein consumption. J. Agric. Food Chem. 2009, 57, 1260–1266. [Google Scholar] [CrossRef]
- Park, J.H.; Jeong, H.J.; de Lumen, B.O. In vitro digestibility of the cancer-preventive soy peptides lunasin and BBI. J. Agric. Food Chem. 2007, 55, 10703–10706. [Google Scholar] [CrossRef]
- Hsieh, C.-C.; Hernández-Ledesma, B.; Jeong, H.J.; Park, J.H.; de Lumen, B.O. Complementary roles in cancer prevention: Protease inhibitor makes the cancer preventive peptide lunasin bioavailable. PLoS ONE 2010, 5, e8890. [Google Scholar] [CrossRef]
- Cruz-Huerta, E.; Fernández-Tomé, S.; Arques, M.C.; Amigo, L.; Recio, I.; Clemente, A.; Hernández-Ledesma, B. The protective role of the Bowman-Birk protease inhibitor in soybean lunasin digestion: The effect of released peptides on colon cancer growth. Food Funct. 2015, 6, 2626–2635. [Google Scholar] [CrossRef]
- Prasad, C.; Prasad, C.; Davis, K.E.; Imrhan, V.; Juma, S.; Vijayagopal, P. Advanced glycation end products and risks for chronic diseases: Intervening through lifestyle modification. Am. J. Lifestyle Med. 2019, 13, 384–404. [Google Scholar] [CrossRef]
- Friso, G.; van Wijk, K.J. Posttranslational protein modifications in plant metabolism. Plant Physiol. 2015, 169, 1469–1487. [Google Scholar]
- Gazme, B.; Boachie, R.T.; Tsopmo, A.; Udenigwe, C.C. Occurrence, properties and biological significance of pyroglutamyl peptides derived from different food sources. Food Sci. Hum. Wellness 2019, 8, 268–274. [Google Scholar] [CrossRef]
- Feige, M.J.; Braakman, I.; Hendershot, L.M. Disulfide bonds in protein folding and stability. In Oxidative Folding of Proteins: Basic Principles, Cellular Regulation and Engineering; Feige, M.J., Ed.; The Royal Society of Chemistry: London, UK, 2018; pp. 1–33. [Google Scholar]
- Lodish, H.; Berk, A.; Zipursky, S. Post-translational modifications and quality control in the rough ER. In Molecular Cell Biology, 4th ed.; Lodish, H., Berk, A., Zipursky, S., Matsudaira, P., Baltimore, D., Darnell, J., Eds.; W.H. Freeman: New York, NY, USA, 2000; pp. 707–712. [Google Scholar]
- Zhang, L.; Chou, C.P.; Moo-Young, M. Disulfide bond formation and its impact on the biological activity and stability of recombinant therapeutic proteins produced by Escherichia coli expression system. Biotechnol. Adv. 2011, 29, 923–929. [Google Scholar] [CrossRef] [PubMed]
- Aleksis, R.; Jaudzems, K.; Muceniece, R.; Liepinsh, E. Lunasin is a redox sensitive intrinsically disordered peptide with two transiently populated α-helical regions. Peptides 2016, 85, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Bisetty, K. A molecular dynamics study of lunasin. S. Afr. J. Chem. 2012, 65, 115–124. [Google Scholar]
- Dia, V.P.; Frankland-Searby, S.; Del Hierro, F.L.; Garcia, G.; De Mejia, E.G. Structural property of soybean lunasin and development of a method to quantify lunasin in plasma using an optimized immunoassay protocol. Food Chem. 2013, 138, 334–341. [Google Scholar] [CrossRef]
- Habchi, J.; Tompa, P.; Longhi, S.; Uversky, V.N. Introducing protein intrinsic disorder. Chem. Rev. 2014, 114, 6561–6588. [Google Scholar] [CrossRef]
- Tompa, P. The interplay between structure and function in intrinsically unstructured proteins. FEBS Lett. 2014, 579, 3346–3354. [Google Scholar] [CrossRef]
- Jeong, H.J.; Jeong, J.B.; Kim, D.S.; Park, J.H.; Lee, J.B.; Kweon, D.H.; Chung, G.Y.; Seo, E.W.; de Lumen, B.O. The cancer preventive peptide lunasin from wheat inhibits core histone acetylation. Cancer Lett. 2007, 255, 42–48. [Google Scholar] [CrossRef]
- European Medicines Agency. Guideline on the Clinical Investigation of the Pharmacokinetics of Therapeutic Proteins CHMP/EWP/89249/2004; European Medicines Agency: London, UK, 2007. [Google Scholar]
- Vugmeyster, Y.; Xu, X.; Theil, F.P.; Khawli, L.A.; Leach, M.W. Pharmacokinetics and toxicology of therapeutic proteins: Advances and challenges. World J. Biol. Chem. 2012, 3, 73. [Google Scholar] [CrossRef]
- Lu, Z.; Sheng, J.; Zhang, W. Pharmacokinetics of therapeutic proteins. In Protein Therapeutics; Vaughan, T., Osbourne, J., Jallal, B., Eds.; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2017; pp. 311–340. [Google Scholar]
- Meibohm, B. Pharmacokinetics and pharmacodynamics of therapeutic peptides and proteins. In Pharmaceutical Biotechnology; Crommelin, D., Sindelar, R., Meibohm, B., Eds.; Springer: Cham, Switzerland, 2019; pp. 105–137. [Google Scholar]
- de Mejia, E.G.; Wang, W.; Dia, V.P. Lunasin, with an arginine–glycine–aspartic acid motif, causes apoptosis to L1210 leukemia cells by activation of caspase-3. Mol. Nutr. Food Res. 2010, 54, 406–414. [Google Scholar] [CrossRef]
- Jeong, H.J.; Jeong, J.B.; Kim, D.S.; de Lumen, B.O. Inhibition of core histone acetylation by the cancer preventive peptide lunasin. J. Agric. Food Chem. 2007, 55, 632–637. [Google Scholar] [CrossRef]
- Price, S.J.; Pangloli, P.; Krishnan, H.B.; Dia, V.P. Kunitz trypsin inhibitor in addition to Bowman-Birk inhibitor influence stability of lunasin against pepsin-pancreatin hydrolysis. Food Res. Int. 2016, 90, 205–215. [Google Scholar] [CrossRef]
- Dan, N.; Samanta, K.; Almoazen, H. An update on pharmaceutical strategies for oral delivery of therapeutic peptides and proteins in adults and pediatrics. Children 2020, 7, 307. [Google Scholar] [CrossRef]
- Renukuntla, J.; Vadlapudi, A.D.; Patel, A.; Boddu, S.H.; Mitra, A.K. Approaches for enhancing oral bioavailability of peptides and proteins. Int. J. Pharm. 2013, 447, 75–93. [Google Scholar] [CrossRef]
- Fernández-Tomé, S.; Sanchón, J.; Recio, I.; Hernández-Ledesma, B. Transepithelial transport of lunasin and derived peptides: Inhibitory effects on the gastrointestinal cancer cells viability. J. Food Comp. Anal. 2018, 68, 101–110. [Google Scholar] [CrossRef]
- Dzirkale, Z.; Nakurte, I.; Jekabsons, K.; Muceniece, R.; Klusa, V. Intra-nasally administered oligopeptide lunasin acts as a possible anti-psychotic agent in mice models. Medicina 2019, 55, 393. [Google Scholar] [CrossRef]
- Hsieh, E.A.; Chai, C.M.; de Lumen, B.O.; Neese, R.A.; Hellerstein, M.K. Dynamics of keratinocytes in vivo using HO labeling: A sensitive marker of epidermal proliferation state. J. Investig. Dermatol. 2004, 123, 530–536. [Google Scholar] [CrossRef]
- Yang, X.; Zhu, J.; Tung, C.Y.; Gardiner, G.; Wang, Q.; Chang, H.C.; Zhou, B. Lunasin alleviates allergic airway inflammation while increases antigen-specific Tregs. PLoS ONE 2015, 10, e0115330. [Google Scholar] [CrossRef]
- Mentlein, R. Cell-surface peptidases. Int. Rev. Cytol. 2004, 235, 165. [Google Scholar]
- García-Nebot, M.J.; Recio, I.; Hernández-Ledesma, B. Antioxidant activity and protective effects of peptide lunasin against oxidative stress in intestinal Caco-2 cells. Food Chem. Toxicol. 2014, 65, 155–161. [Google Scholar] [CrossRef]
- Fernández-Tomé, S.; Ramos, S.; Cordero-Herrera, I.; Recio, I.; Goya, L.; Hernández-Ledesma, B. In vitro chemo-protective effect of bioactive peptide lunasin against oxidative stress in human HepG2 cells. Food Res. Int. 2014, 62, 793–800. [Google Scholar] [CrossRef]
- Cam, A.; de Mejia, E.G. RGD-peptide lunasin inhibits Akt-mediated NF-κB activation in human macrophages through interaction with the αVβ3 integrin. Mol. Nutr. Food Res. 2012, 56, 1569–1581. [Google Scholar] [CrossRef]
- Cam, A.; Sivaguru, M.; Gonzalez de Mejia, E. Endocytic mechanism of internalization of dietary peptide lunasin into macrophages in inflammatory condition associated with cardiovascular disease. PLoS ONE 2013, 8, e72115. [Google Scholar] [CrossRef]
- Conner, K.P.; Devanaboyina, S.C.; Thomas, V.A.; Rock, D.A. The biodistribution of therapeutic proteins: Mechanism, implications for pharmacokinetics, and methods of evaluation. Pharmacol. Ther. 2020, 212, 107574. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Jeong, H.J.; de Lumen, B.O. Contents and bioactivities of lunasin, bowman− birk inhibitor, and isoflavones in soybean seed. J. Agric. Food. Chem. 2005, 53, 7686–7690. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.-C.; Hernández-Ledesma, B.; de Lumen, B.O. Lunasin–aspirin combination against NIH/3T3 cells transformation induced by chemical carcinogens. Plant Foods Hum. Nutr. 2011, 66, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Setrerrahmane, S.; Zhang, Y.; Dai, G.; Lv, J.; Tan, S. Efficient production of native lunasin with correct N-terminal processing by using the pH-induced self-cleavable Ssp DnaB mini-intein system in Escherichia coli. Appl. Biochem. Biotechnol. 2014, 174, 612–622. [Google Scholar] [CrossRef]
- Tian, Q.; Zhang, P.; Gao, Z.; Li, H.; Bai, Z.; Tan, S. Hirudin as a novel fusion tag for efficient production of lunasin in Escherichia coli. Prep. Biochem. Biotechnol. 2017, 47, 619–626. [Google Scholar] [CrossRef]
- Gu, L.; Ye, P.; Li, H.; Wang, Y.; Xu, Y.; Tian, Q.; Lei, G.; Zhao, C.; Gao, Z.; Zhao, W.; et al. Lunasin attenuates oxidant-induced endothelial injury and inhibits atherosclerotic plaque progression in ApoE−/− mice by up-regulating heme oxygenase-1 via PI3K/Akt/Nrf2/ARE pathway. FASEB J. 2019, 33, 4836–4850. [Google Scholar] [CrossRef]
- Dia, V.P.; de Mejia, E.G. Lunasin induces apoptosis and modifies the expression of genes associated with extracellular matrix and cell adhesion in human metastatic colon cancer cells. Mol. Nutr. Food Res. 2011, 55, 623–634. [Google Scholar] [CrossRef]
- Jiang, Q.; Pan, Y.; Cheng, Y.; Li, H.; Liu, D.; Li, H. Lunasin suppresses the migration and invasion of breast cancer cells by inhibiting matrix metalloproteinase-2/-9 via the FAK/Akt/ERK and NF-κB signaling pathways. Oncol. Rep. 2016, 36, 253–262. [Google Scholar] [CrossRef]
- McConnell, E.J.; Devapatla, B.; Yaddanapudi, K.; Davis, K.R. The soybean-derived peptide lunasin inhibits non-small cell lung cancer cell proliferation by suppressing phosphorylation of the retinoblastoma protein. Oncotarget 2015, 6, 4649. [Google Scholar] [CrossRef]
- Hsieh, C.-C. Lunasin attenuates obesity-associated metastasis of 4T1 breast cancer cell through anti-inflammatory property. Int. J. Mol. Sci. 2016, 17, 2109. [Google Scholar] [CrossRef]
- de Mejia, E.G.; Dia, V.P. Lunasin and lunasin-like peptides inhibit inflammation through suppression of NF-κB pathway in the macrophage. Peptides 2009, 30, 2388–2398. [Google Scholar] [CrossRef]
- Hsieh, C.-C.; Chou, M.J.; Wang, C.H. Lunasin attenuates obesity-related inflammation in RAW264. 7 cells and 3T3-L1 adipocytes by inhibiting inflammatory cytokine production. PLoS ONE 2017, 12, e0171969. [Google Scholar] [CrossRef]
- Dai, W.; Liang, Z.; Liu, H.; Zhao, G.; Ju, C. Lunasin abrogates the expression of matrix metalloproteinases and reduction of type II collagen. Artif. Cells Nanomed. Biotechnol. 2019, 47, 3259–3264. [Google Scholar] [CrossRef]
- Jia, S.; Zhang, S.; Yuan, H.; Chen, N. Lunasin inhibits cell proliferation via apoptosis and reduces the production of proinflammatory cytokines in cultured rheumatoid arthritis synovial fibroblasts. BioMed Res. Int. 2015, 2015, 346839. [Google Scholar] [CrossRef]
- Nakurte, I.; Kirhnere, I.; Namniece, J.; Saleniece, K.; Krigere, L.; Mekss, P.; Vicupe, Z.; Bleidere, M.; Legzdina, L.; Muceniece, R. Detection of the lunasin peptide in oats (Avena sativa L). J. Cer. Sci. 2013, 57, 319–324. [Google Scholar] [CrossRef]
- Kavsan, V.M.; Iershov, A.V.; Balynska, O.V. Immortalized cells and one oncogene in malignant transformation: Old insights on new explanation. BMC Cell. Biol. 2011, 12, 23. [Google Scholar] [CrossRef]
- Stepanenko, A.A.; Dmitrenko, V.V. HEK293 in cell biology and cancer research: Phenotype, karyotype, tumorigenicity, and stress-induced genome-phenotype evolution. Gene 2015, 569, 182–190. [Google Scholar] [CrossRef]
- Shidal, C.; Al-Rayyan, N.; Yaddanapudi, K.; Davis, K.R. Lunasin is a novel therapeutic agent for targeting melanoma cancer stem cells. Oncotarget 2016, 7, 84128. [Google Scholar] [CrossRef]
- Dia, V.P.; de Mejia, E.G. Lunasin promotes apoptosis in human colon cancer cells by mitochondrial pathway activation and induction of nuclear clusterin expression. Cancer Lett. 2010, 295, 44–53. [Google Scholar] [CrossRef]
- Takeshima, H.; Ushijima, T. Accumulation of genetic and epigenetic alterations in normal cells and cancer risk. NPJ Precis. Oncol. 2019, 3, 7. [Google Scholar] [CrossRef]
- Dagogo-Jack, I.; Shaw, A.T. Tumour heterogeneity and resistance to cancer therapies. Nat. Rev. Clin. Oncol. 2018, 15, 81–94. [Google Scholar] [CrossRef]
- Palmer, A.C.; Sorger, P.K. Combination cancer therapy can confer benefit via patient-to-patient variability without drug additivity or synergy. Cell 2017, 171, 1678–1691. [Google Scholar] [CrossRef]
- Zhu, L.; Jiang, M.; Wang, H.; Sun, H.; Zhu, J.; Zhao, W.; Fang, Q.; Yu, J.; Chen, P.; Wu, S.; et al. A narrative review of tumor heterogeneity and challenges to tumor drug therapy. Ann. Transl. Med. 2021, 9, 1351. [Google Scholar] [CrossRef]
- Fernández-Tomé, S.; Xu, F.; Han, Y.; Hernández-Ledesma, B.; Xiao, H. Inhibitory effects of peptide lunasin in colorectal cancer HCT-116 cells and their tumorsphere-derived subpopulation. Int. J. Mol. Sci. 2020, 21, 537. [Google Scholar] [CrossRef]
- Montales, M.T.E.; Simmen, R.C.; Ferreira, E.S.; Neves, V.A.; Simmen, F.A. Metformin and soybean-derived bioactive molecules attenuate the expansion of stem cell-like epithelial subpopulation and confer apoptotic sensitivity in human colon cancer cells. Genes Nutr. 2015, 10, 49. [Google Scholar] [CrossRef]
- Pabona, J.M.P.; Dave, B.; Su, Y.; Montales, M.T.; de Lumen, B.O.; de Mejia, E.G.; Rahal, O.M.; Simmen, R.C. The soybean peptide lunasin promotes apoptosis of mammary epithelial cells via induction of tumor suppressor PTEN: Similarities and distinct actions from soy isoflavone genistein. Genes Nutr. 2013, 8, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.-C.; Hernández-Ledesma, B.; de Lumen, B.O. Lunasin, a novel seed peptide, sensitizes human breast cancer MDA-MB-231 cells to aspirin-arrested cell cycle and induced apoptosis. Chem. Biol. Interact. 2010, 186, 127–134. [Google Scholar] [CrossRef]
- Hsieh, C.C.; Hernández-Ledesma, B.; de Lumen, B.O. Cell proliferation inhibitory and apoptosis-inducing properties of anacardic acid and lunasin in human breast cancer MDA-MB-231 cells. Food Chem. 2011, 125, 630–636. [Google Scholar] [CrossRef]
- Devapatla, B.; Shidal, C.; Yaddanapudi, K.; Davis, K.R. Validation of syngeneic mouse models of melanoma and non-small cell lung cancer for investigating the anticancer effects of the soy-derived peptide Lunasin. F1000 Res. 2016, 5, 2432. [Google Scholar] [CrossRef]
- Lea, T. Caco-2 cell line. In The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Verhoeckx, K., Ed.; Springer: Cham, Switzerland, 2015; pp. 103–111. [Google Scholar]
- Donato, M.T.; Tolosa, L.; Gómez-Lechón, M.J. Culture and functional characterization of human hepatoma HepG2 cells. Methods Mol. Biol. 2015, 1250, 77–93. [Google Scholar] [PubMed]
- Auwerx, J. The human leukemia cell line, THP-1: A multifacetted model for the study of monocyte-macrophage differentiation. Experientia 1991, 4, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Dzirkale, Z.; Rumaks, J.; Svirskis, S.; Mazina, O.; Allikalt, A.; Rinken, A.; Jekabsons, K.; Muceniece, R.; Klusa, V. Lunasin-induced behavioural effects in mice: Focus on the dopaminergic system. Behav. Brain Res. 2013, 256, 5–9. [Google Scholar] [CrossRef]
- Galvez, A.F.; Huang, L.; Magbanua, M.M.; Dawson, K.; Rodriguez, R.L. Differential expression of thrombospondin (THBS1) in tumorigenic and nontumorigenic prostate epithelial cells in response to a chromatin-binding soy peptide. Nutr. Cancer 2011, 63, 623–636. [Google Scholar] [CrossRef]
- Inaba, J.; McConnell, E.J.; Davis, K.R. Lunasin sensitivity in non-small cell lung cancer cells is linked to suppression of integrin signaling and changes in histone acetylation. Int. J. Mol. Sci. 2014, 15, 23705–23724. [Google Scholar] [CrossRef]
- Jeong, H.J.; Park, J.H.; Lam, Y.; de Lumen, B.O. Characterization of lunasin isolated from soybean. J. Agric. Food Chem. 2003, 51, 7901–7906. [Google Scholar] [CrossRef]
- Jeong, J.B.; Jeong, H.J.; Park, J.H.; Lee, S.H.; Lee, J.R.; Lee, H.K.; Chung, G.Y.; Choi, J.D.; de Lumen, B.O. Cancer-preventive peptide lunasin from Solanum nigrum L. inhibits acetylation of core histones H3 and H4 and phosphorylation of retinoblastoma protein (Rb). J. Agric. Food Chem. 2007, 55, 10707–10713. [Google Scholar] [CrossRef]
- Chen, H.P.; Zhao, Y.T.; Zhao, T.C. Histone deacetylases and mechanisms of regulation of gene expression. Crit. Rev. Oncog. 2015, 20, 35–47. [Google Scholar] [CrossRef]
- Dia, V.P.; de Mejia, E.G. Lunasin potentiates the effect of oxaliplatin preventing outgrowth of colon cancer metastasis, binds to α5β1 integrin and suppresses FAK/ERK/NF-κB signaling. Cancer Lett. 2011, 313, 167–180. [Google Scholar] [CrossRef]
- D’Souza, S.E.; Ginsberg, M.H.; Plow, E.F. Arginyl-glycyl-aspartic acid (RGD): A cell adhesion motif. Trends Biochem. Sci. 1991, 16, 246–250. [Google Scholar] [CrossRef]
- Ruoslahti, E. RGD and other recognition sequences for integrins. Ann. Rev. Cell Dev. Biol. 1996, 12, 697–715. [Google Scholar] [CrossRef]
- Harburger, D.S.; Calderwood, D.A. Integrin signalling at a glance. J. Cell Sci. 2009, 122, 159–163. [Google Scholar] [CrossRef]
- Klein, S.; de Fougerolles, A.R.; Blaikie, P.; Khan, L.; Pepe, A.; Green, C.D.; Koteliansky, V.; Giancotti, F.G. α5β1 integrin activates an NF-κB-dependent program of gene expression important for angiogenesis and inflammation. Mol. Cell Biol. 2002, 22, 5912–5922. [Google Scholar] [CrossRef]
- Scatena, M.; Almeida, M.; Chaisson, M.L.; Fausto, N.; Nicosia, R.F.; Giachelli, C.M. NF-κB mediates αvβ3 integrin-induced endothelial cell survival. J. Cell Biol. 1998, 141, 1083–1093. [Google Scholar] [CrossRef]
- Li, M.; Wang, Y.; Li, M.; Wu, X.; Setrerrahmane, S.; Xu, H. Integrins as attractive targets for cancer therapeutics. Acta Pharm. Sin. B 2021, 11, 2726–2737. [Google Scholar] [CrossRef]
- Giacinti, C.; Giordano, A. RB and cell cycle progression. Oncogene 2006, 25, 5220–5227. [Google Scholar] [CrossRef]
- Moreno-Layseca, P.; Streuli, C.H. Signalling pathways linking integrins with cell cycle progression. Matrix Biol. 2014, 34, 144–153. [Google Scholar] [CrossRef]
- Dia, V.P.; Wang, W.; Oh, V.L.; de Lumen, B.O.; de Mejia, E.G. Isolation, purification and characterisation of lunasin from defatted soybean flour and in vitro evaluation of its anti-inflammatory activity. Food Chem. 2009, 114, 108–115. [Google Scholar] [CrossRef]
- Tuli, H.S.; Tuorkey, M.J.; Thakral, F.; Sak, K.; Kumar, M.; Sharma, A.K.; Sharma, U.; Jain, A.; Aggarwal, A.; Bishayee, A. Molecular mechanisms of action of genistein in cancer: Recent advances. Front. Pharmacol. 2019, 10, 1336. [Google Scholar] [CrossRef]
- Banerjee, S.; Li, Y.; Wang, Z.; Sarkar, F.H. Multi-targeted therapy of cancer by genistein. Cancer Lett. 2008, 269, 226–242. [Google Scholar] [CrossRef]
- Rasheed, S.; Rehman, K.; Shahid, M.; Suhail, S.; Akash, M.S.H. Therapeutic potentials of genistein: New insights and perspectives. J. Food Biochem. 2022, e14228. [Google Scholar] [CrossRef]
- Marini, H.; Bitto, A.; Altavilla, D.; Burnett, B.P.; Polito, F.; Di Stefano, V.; Minutoli, L.; Atteritano, M.; Levy, R.M.; D’Anna, R.; et al. Breast safety and efficacy of genistein aglycone for postmenopausal bone loss: A follow-up study. J. Clin. Endocrinol. Metab. 2008, 93, 4787–4796. [Google Scholar] [CrossRef]
- Lazarevic, B.; Boezelijn, G.; Diep, L.M.; Kvernrod, K.; Ogren, O.; Ramberg, H.; Moen, A.; Wessel, N.; Berg, R.E.; Egge-Jacobsen, W.; et al. Efficacy and safety of short-term genistein intervention in patients with localized prostate cancer prior to radical prostatectomy: A randomized, placebo-controlled, double-blind Phase 2 clinical trial. Nutr. Cancer. 2011, 63, 889–898. [Google Scholar] [CrossRef]
- Bloedon, L.T.; Jeffcoat, A.R.; Lopaczynski, W.; Schell, M.J.; Black, T.M.; Dix, K.J.; Thomas, B.F.; Albright, C.; Busby, M.J.; Crowell, J.A.; et al. Safety and pharmacokinetics of purified soy isoflavones: Single-dose administration to postmenopausal women. Am. J. Clin. Nutr. 2002, 76, 1126–1137. [Google Scholar] [CrossRef]
- Shankar, M.G.; Swetha, M.; Keerthana, C.K.; Rayginia, T.P.; Anto, R.J. Cancer Chemoprevention: A Strategic Approach Using Phytochemicals. Front. Pharmacol. 2021, 12, 809308. [Google Scholar]
- Marini, H.R. Mediterranean Diet and Soy Isoflavones for Integrated Management of the Menopausal Metabolic Syndrome. Nutrients 2022, 14, 1550. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Part of the Sequence | Activity | Ref. |
|---|---|---|
| SKWQHQQDSCRKQLQGVNLTPCEKHIME-KIQGRGDDDDDDDDD | Cytotoxicity | [13] |
| Anti-transformation | [14,16] | |
| Anti-inflammatory | [15] | |
| SKWQHQQDSCRKQLQGVNLTPCEKHIMEKIQGDDDDDDDDD | Inhibition of histone H4 acetylation | [14] |
| SKWQHQQDSCRKQLQGVNLTPCEKHIMEKIQG | Antioxidant | [15,17] |
| Immunomodulatory | [18] | |
| EKHIMEKIQG | Chromatin binding | [14] |
| RGD | Cell internalization | [14] |
| DDDDDDDDD | Inhibition of histone H3 acetylation | [5,13] |
| Cell Line | Lunasin Source | Concentration | Assay | Effects | Ref. |
|---|---|---|---|---|---|
| NIH/3T3 | Synthetic | 10 µM | Coulter counter | No effect | [16] |
| Synthetic | 100 nM | MTT | No effect | [61] | |
| Synthetic | 0.01–10 µM | MTT | No effect | [62] | |
| SRA 01/04 | Recombinant | 1–100 µM | MTT | No effect | [63] |
| Recombinant | 20, 40, 80 µM | Apoptosis detection (Annexin V-PE/7-AAD) | No effect | [64] | |
| CCD-33Co | Soybean (~90%) | 1–100 µM | MTS | No effect | [78] |
| Soybean (~90%) | 0–100 µM | MTS | No effect | [66] | |
| BEAS-2B | Soybean (>99%) | 1–100 µM | MTS | No effect | [68] |
| HBE135-E6E7 | Soybean (>99%) | 1–100 µM | MTS | No effect | [68] |
| MCF-10A | Synthetic | 0–320 µM | MTT | No effect | [67] |
| EA.hy926 | Recombinant | 0–120 µM | MTT | No effect | [65] |
| HEK-293 | Synthetic | 0.1–100 µM | MTT | ↓ Proliferation at ˃100 µM | [74] |
| Chondrocyte | Synthetic | 50–500 µM | MTT | ↓ Proliferation at ˃500 µM | [72] |
| 3T3-L1 | Synthetic | 0.1–25 µM | MTT | No effect | [69] |
| Synovial Fibroblast | Synthetic | 0–200 µM | Crystal violet staining and apoptosis detection (annexin V-FITC/PI) | ↓ Proliferation (IC50 153.3 µM). Apoptosis induction | [73] |
| RAW 264.7 | Synthetic | 0.2–200 µM | MTT | No effect | [15] |
| Soybean (85.3%) | 10–50 µM | MTS | No effect | [70] | |
| Synthetic | 0.2–200 µM | MTT | No effect | [71] | |
| Synthetic | 10–100 µM | MTT | No effect | [17] |
| Tissue | Cell Line | Lunasin Source | Concentration | Assay | Effects | Ref. |
|---|---|---|---|---|---|---|
| Colon | HT-29 (Human) | Soybean (~90%) | 1–100 µM | MTS and crystal violet staining | ↓ Proliferation (IC50 61.7 µM) and morphologic alterations | [78] |
| Soybean (>90%) | 1–100 µM | MTS | ↓ Proliferation (IC50 61.7 µM) | [66] | ||
| Synthetic | 10–200 µM | MTT | ↓ Proliferation | [51] | ||
| HCT-116 (Human) | Soybean (>90%) | 1–100 µM | MTS | ↓ Proliferation (IC50 26.3 µM) | [66] | |
| Recombinant | 1–100 µM | MTT | ↓ Proliferation (IC50 64.25 μM) | [63] | ||
| Synthetic | 2 µM | Trypan blue exclusion, apoptosis detection (annexin V) and colonosphere formation | ↓ Proliferation. ↑ Apoptotic cells. ↓ Colonosphere formation | [84] | ||
| Recombinant | 20, 40 and 80 µM | Apoptosis detection (annexin V-PE/7-AAD) | ↑ Apoptotic cells | [64] | ||
| Synthetic | 5–160 µM | MTT, tumorsphere formation and apoptosis detection (annexin V/PI) | ↓ Proliferation (IC50 107.5 µM). ↓ Tumorsphere formation (IC50 161 µM). ↑ Apoptotic cells | [83] | ||
| HCT-116OxR (Human) | Soybean (>90%) | 1–100 µM | MTS | ↓ Proliferation (IC50 31.6 µM) | [66] | |
| KM12L4 (Human) | Soybean (>90%) | 1–100 µM | MTS | ↓ Proliferation (IC50 13 µM) | [66] | |
| KM12L4OxR (Human) | Soybean (>90%) | 1–100 µM | MTS | ↓ Proliferation (IC50 34.7 µM) | [66] | |
| RKO (Human) | Soybean (>90%) | 1–100 µM | MTS | ↓ Proliferation (IC50 21.6 µM) | [66] | |
| RKOOxR (Human) | Soybean (>90%) | 1–100 µM | MTS | ↓ Proliferation (IC50 38.9 µM) | [66] | |
| Caco-2 (Human) | Synthetic | 0.5–25 µM | MTT | No effect | [56] | |
| Synthetic | 10–200 µM | MTT | Modest decrease in viability (24 h and 48 h). No cytotoxicity (72 h) | [51] | ||
| Stomach | AGS (Human) | Synthetic | 10–200 µM | MTT | Modest decrease in viability | [51] |
| Liver | HepG2 (Human) | Synthetic | 0.5–50 µM | Crystal violet staining | No effect | [57] |
| Recombinant | 1–100 µM | MTT | No effect | [63] | ||
| Recombinant | 20, 40 and 80 µM | Apoptosis detection (annexin V-PE/7-AAD) | Negligible early apoptosis induction | [64] | ||
| Breast | MCF-7 (Human) | Synthetic | 10 µM | Coulter counter | No effect | [16] |
| Synthetic | 2 µM | TUNEL and mammosphere formation | ↑ Apoptotic cells. No influence on mammosphere formation | [85] | ||
| Synthetic | 0–320 µM | MTT | ↓ Proliferation (IC50 431.9 µM) | [67] | ||
| MDA-MB-231 (Human) | Synthetic | 0.1–200 µM | MTT and apoptosis detection (annexin V/7-AAD) | ↓ Proliferation (IC50 181 µM). No apoptosis induction | [86] | |
| Synthetic | 1–200 µM | MTT and apoptosis detection (annexin V/7-AAD) | ↓ Proliferation (IC50 181 µM). No apoptosis induction | [87] | ||
| Synthetic | 10–200 µM | MTT | ↓ Proliferation (IC50 181 µM) | [13] | ||
| Recombinant | 1–100 µM | MTT | ↓ Proliferation (IC50 56.73 μM) | [63] | ||
| Synthetic | 0–320 µM | MTT | ↓ Proliferation (IC50 194.9 µM) | [67] | ||
| Recombinant | 20, 40 and 80 µM | Apoptosis detection (annexin V-PE/7-AAD) | ↑ Apoptotic cells | [64] | ||
| 4T1 (Mouse) | Synthetic | 1–50 µM | MTT | No effect | [69] | |
| Skin (Melanoma) | A375 (Human) | Soybean (>99%) | 100 µM | MTS and colony Formation | No effect on cell proliferation. ↓ Colony formation (37%) | [77] |
| SKMEL-28 (Human) | Soybean (>99%) | 100 µM | MTS and colony formation | No effect on cell proliferation. ↓ Colony formation (23%) | [77] | |
| B16-F10 (Mouse) | Soybean | 100 µM | Oncosphere formation and Transwell invasion | ↓ Oncosphere formation (29%). ↓ Invasion (60%) | [21] | |
| B16-F0 (Mouse) | Soybean (>99%) | 1–100 µM | MTS and colony formation | Modest decrease in viability. ↓ Colony formation | [88] | |
| Lung | NCI-H661 (Human) | Soybean (>99%) | 1–100 µM | MTS, colony formation and apoptosis detection (annexin V-Cy3™ and 6-CFDA) | ↓ Proliferation (IC50 63.9 µM) ↓ Number of colonies | [68] |
| NCI-H1299 (Human) | Soybean (>99%) | 1–100 µM | MTS and colony formation | No cytotoxicity. ↓ Number and size of colonies. | [68] | |
| NCI-H460 (Human) | Soybean (>99%) | 1–100 µM | MTS and colony formation | No cytotoxicity. ↓ Colony size. | [68] | |
| A549 (Human) | Soybean (>99%) | 1–100 µM | MTS and colony formation | No cytotoxicity. ↓ Colony number | [68] | |
| LLC (Mouse) | Soybean (>99%) | 1–100 µM | MTS and colony formation | Modest decrease in viability. ↓ Colony formation. | [88] | |
| Blood (Leukemia) | THP-1 (Human) | Soybean (>95%) | 10–100 µM | MTS | No effect | [59] |
| L1210 (Mouse) | Soybean (~98%) | 0–100 µM | CCK-8 | ↓ Proliferation (IC50 14 µM) | [46] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alves de Souza, S.M.; Hernández-Ledesma, B.; de Souza, T.L.F. Lunasin as a Promising Plant-Derived Peptide for Cancer Therapy. Int. J. Mol. Sci. 2022, 23, 9548. https://doi.org/10.3390/ijms23179548
Alves de Souza SM, Hernández-Ledesma B, de Souza TLF. Lunasin as a Promising Plant-Derived Peptide for Cancer Therapy. International Journal of Molecular Sciences. 2022; 23(17):9548. https://doi.org/10.3390/ijms23179548
Chicago/Turabian StyleAlves de Souza, Stephanny Miranda, Blanca Hernández-Ledesma, and Theo Luiz Ferraz de Souza. 2022. "Lunasin as a Promising Plant-Derived Peptide for Cancer Therapy" International Journal of Molecular Sciences 23, no. 17: 9548. https://doi.org/10.3390/ijms23179548
APA StyleAlves de Souza, S. M., Hernández-Ledesma, B., & de Souza, T. L. F. (2022). Lunasin as a Promising Plant-Derived Peptide for Cancer Therapy. International Journal of Molecular Sciences, 23(17), 9548. https://doi.org/10.3390/ijms23179548