
Interdomain Linker Effect on the Mechanical Stability of Ig Domains in Titin


Abstract
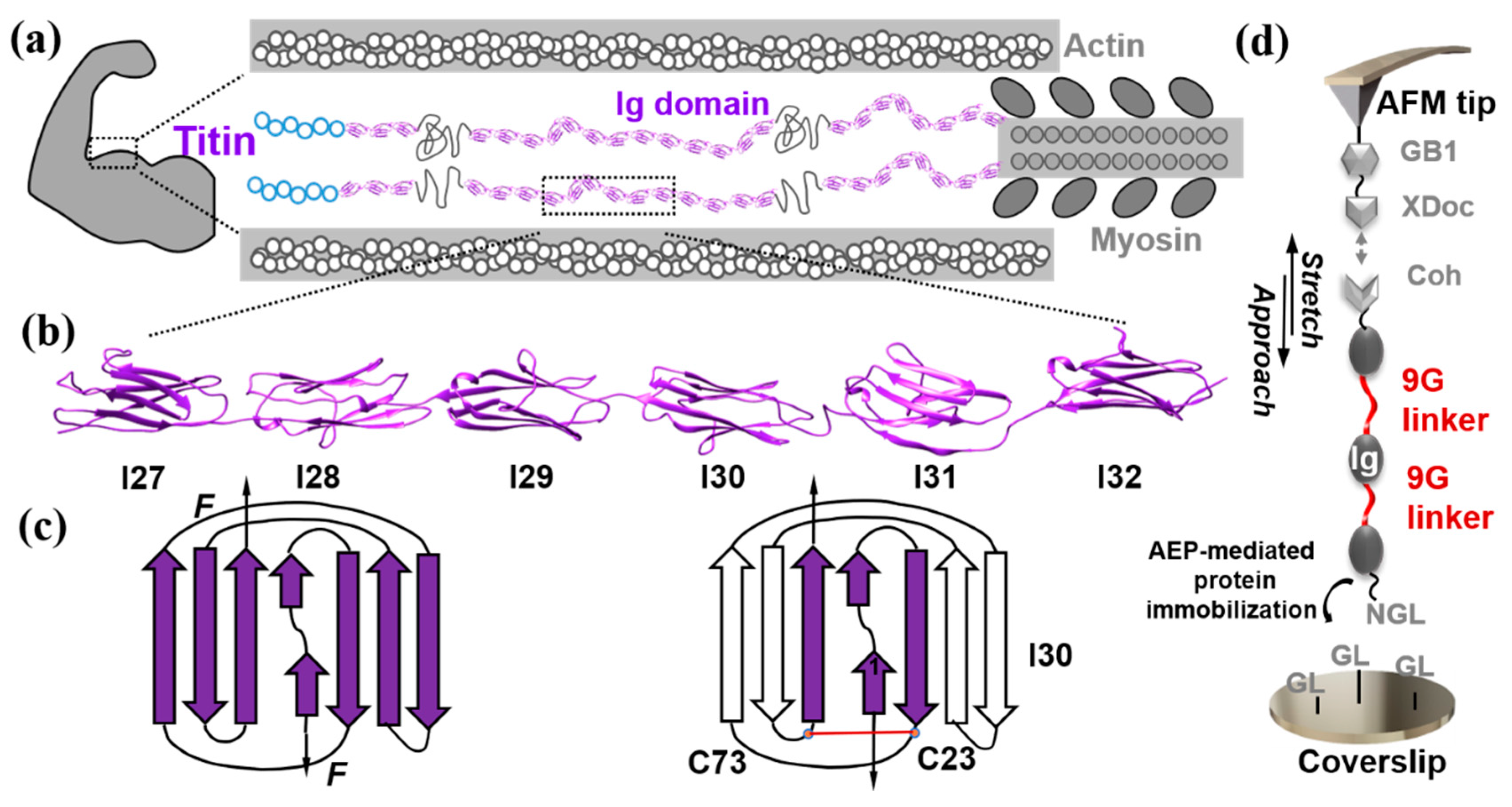
:1. Introduction
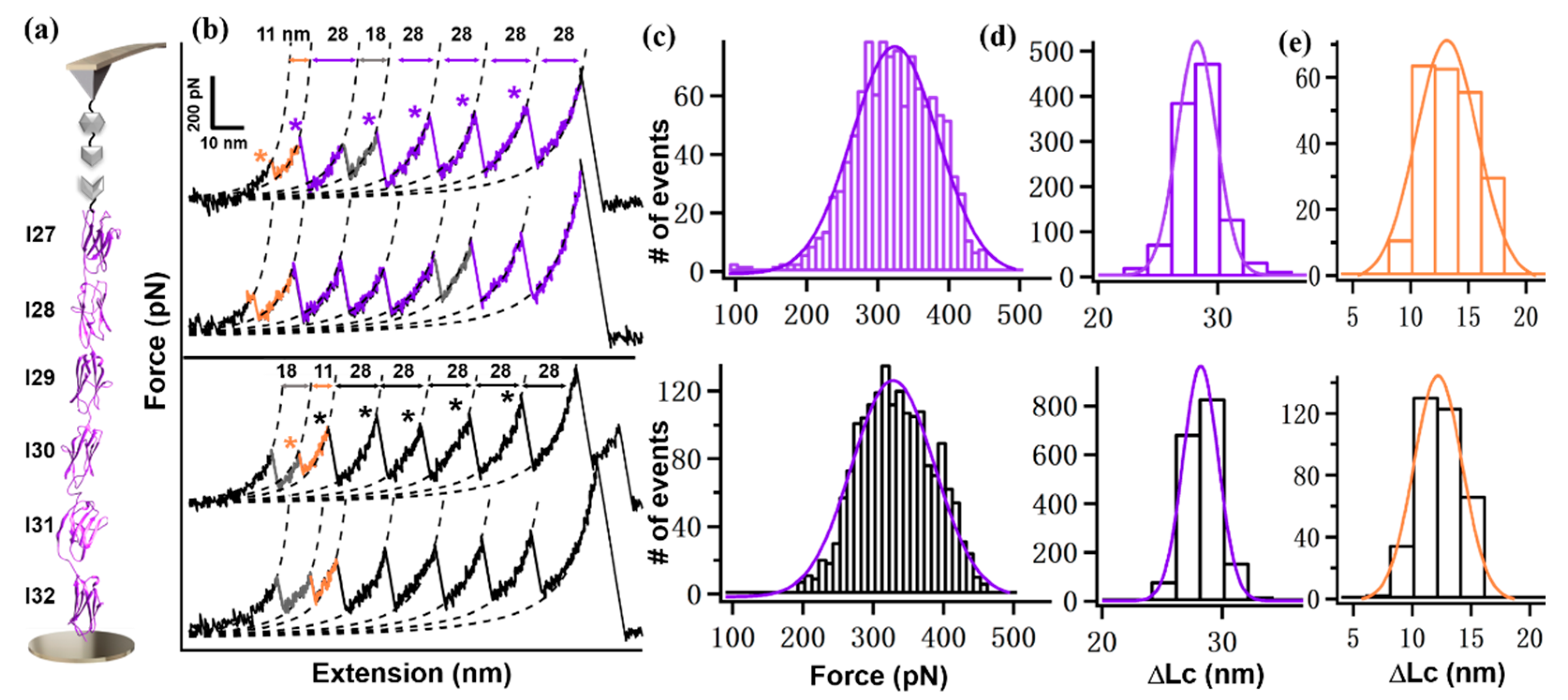
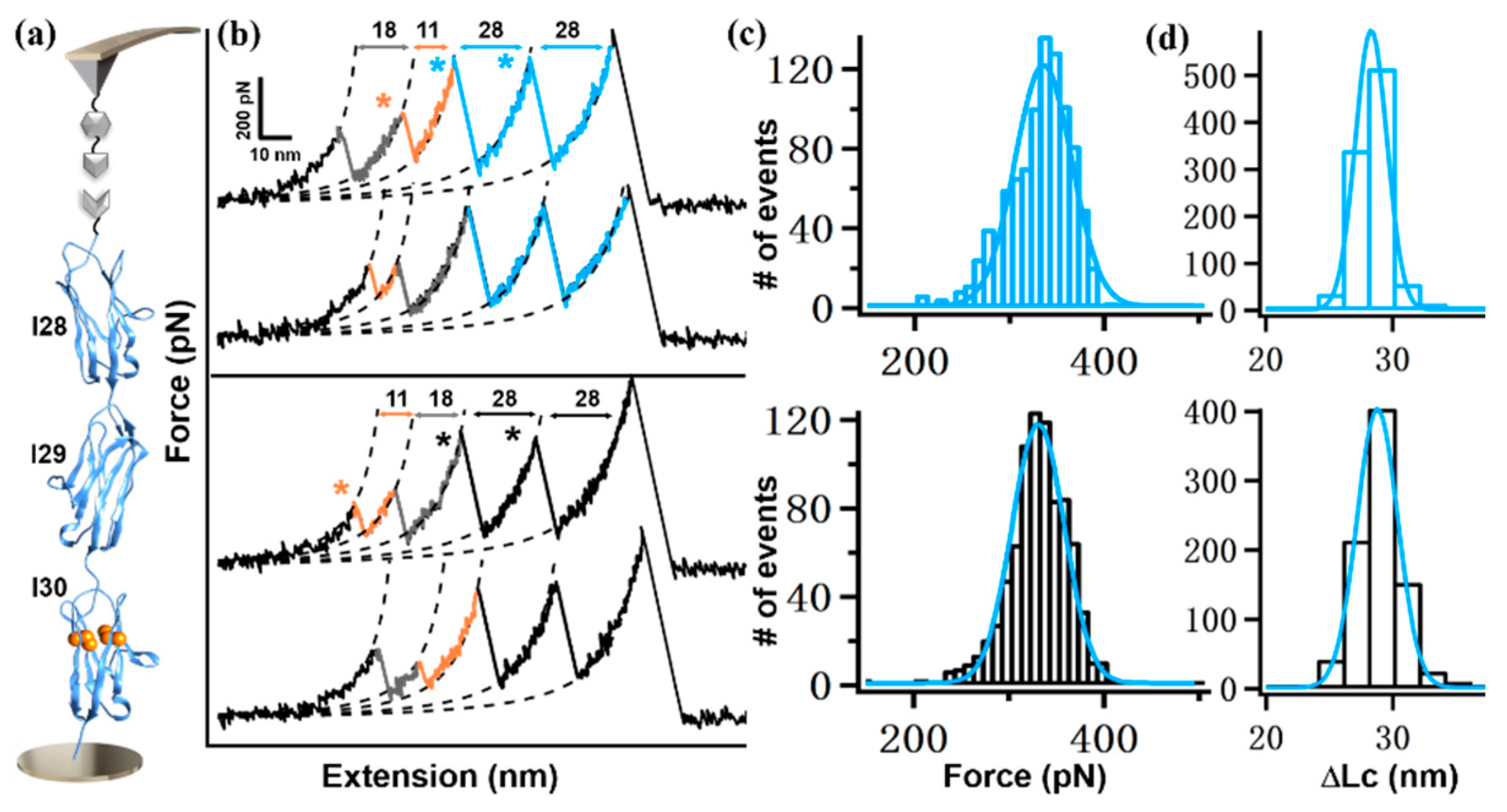
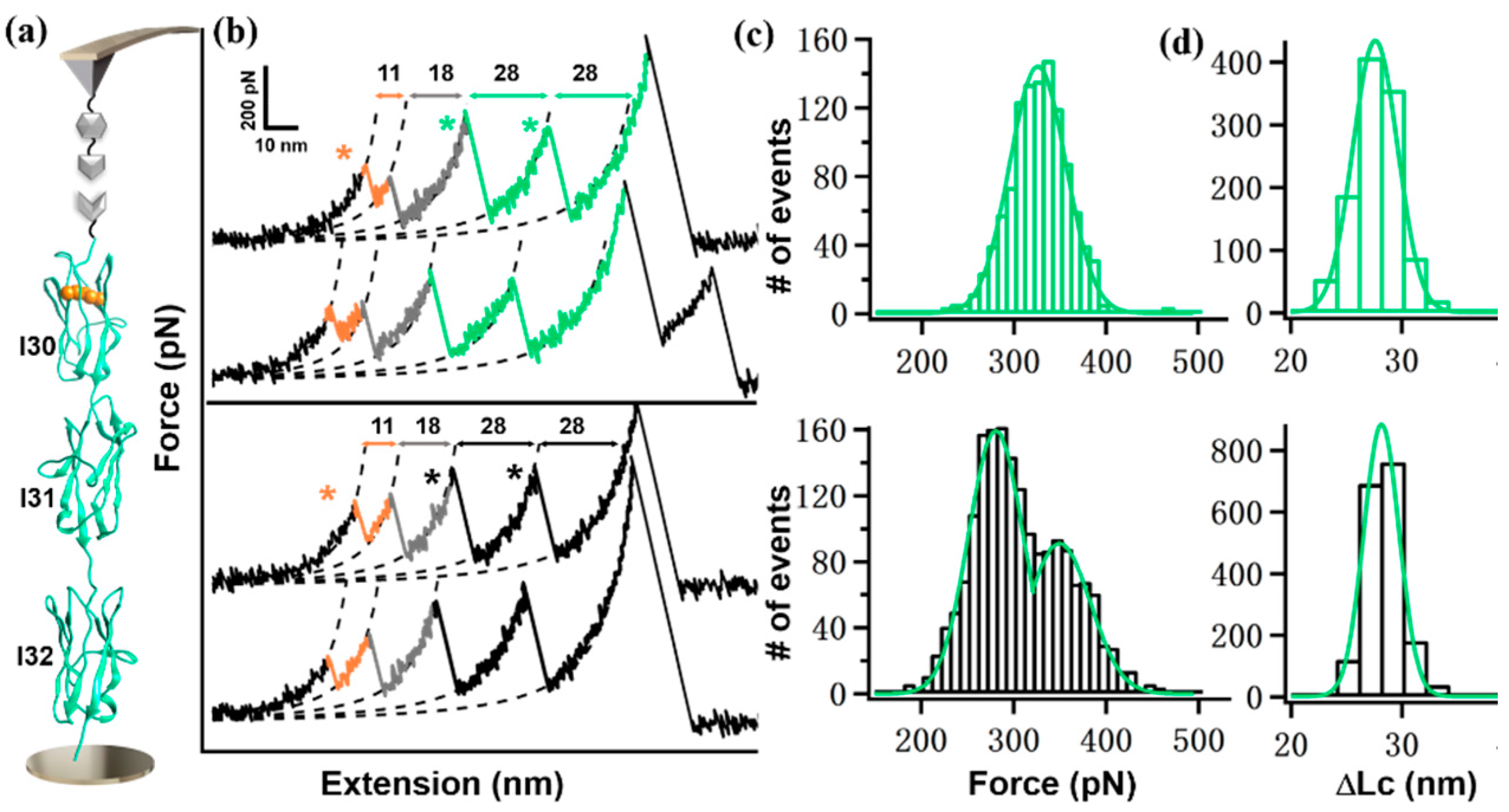
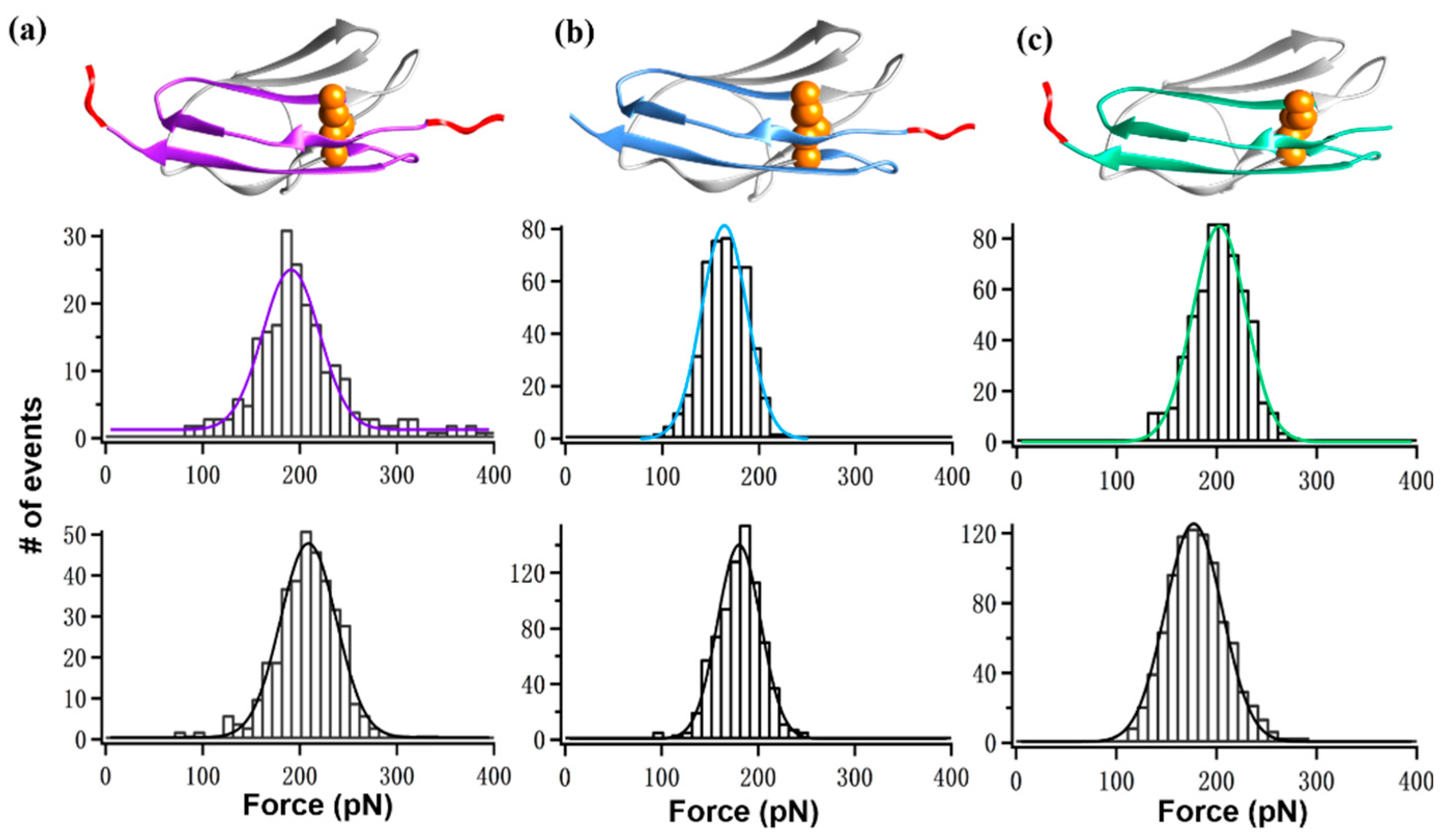
2. Results
3. Method and Material
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Labeit, S.; Kolmerer, B. Titins: Giant proteins in charge of muscle ultrastructure and elasticity. Science 1995, 270, 293–296. [Google Scholar] [CrossRef]
- Wang, R.; Li, J.; Li, X.; Guo, J.; Liu, J.; Li, H. Engineering protein polymers of ultrahigh molecular weight via supramolecular polymerization: Towards mimicking the giant muscle protein titin. Chem. Sci. 2019, 10, 9277–9284. [Google Scholar] [CrossRef] [PubMed]
- Li, H.B.; Linke, W.A.; Oberhauser, A.F.; Carrion-Vazquez, M.; Kerkviliet, J.G.; Lu, H.; Marszalek, P.E.; Fernandez, J.M. Reverse engineering of the giant muscle protein titin. Nature 2002, 418, 998–1002. [Google Scholar] [CrossRef] [PubMed]
- Suay-Corredera, C.; Pricolo, M.R.; Velazquez-Carreras, D.; Pathak, D.; Nandwani, N.; Pimenta-Lopes, C.; Sanchez-Ortiz, D.; Urrutia-Irazabal, I.; Vilches, S.; Dominguez, F.; et al. Nanomechanical phenotypes in cardiac myosin-binding protein c mutants that cause hypertrophic cardiomyopathy. ACS Nano 2021, 15, 10203–10216. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Marszalek, P.E. Exploiting a mechanical perturbation of a titin domain to identify how force field parameterization affects protein refolding pathways. J. Chem. Theory Comput. 2020, 16, 3240–3252. [Google Scholar] [CrossRef] [PubMed]
- Zacharchenko, T.; von Castelmur, E.; Rigden, D.J.; Mayans, O. Structural advances on titin: Towards an atomic understanding of multi-domain functions in myofilament mechanics and scaffolding. Biochem. Soc. Trans. 2015, 43, 850–855. [Google Scholar] [CrossRef]
- Fleming, J.R.; Rigden, D.J.; Mayans, O. The importance of chain context in assessing small nucleotide variants in titin: In silico case study of the i10-i11 tandem and its arrhythmic right ventricular cardiomyopathy linked position t2580. J. Biomol. Struct. Dyn. 2021, 39, 3480–3490. [Google Scholar] [CrossRef]
- Lanzicher, T.; Zhou, T.; Saripalli, C.; Keschrumrus, V.; Smith Iii, J.E.; Mayans, O.; Sbaizero, O.; Granzier, H. Single-molecule force spectroscopy on the n2a element of titin: Effects of phosphorylation and carp. Front. Physiol. 2020, 11, 173. [Google Scholar] [CrossRef]
- Zhang, W.; Hou, J.; Li, N.; Zhang, W.K. Application of atomic force microscopy (AFM)-based single-molecule force spectroscopy (SMFS) in polymer characterization. Acta Polym. Sin. 2021, 52, 1523–1546. [Google Scholar]
- Bao, Y.; Huang, X.; Xu, J.; Cui, S. Effect of intramolecular hydrogen bonds on the single-chain elasticity of poly(vinyl alcohol): Evidencing the synergistic enhancement effect at the single-molecule level. Macromolecules 2021, 54, 7314–7320. [Google Scholar] [CrossRef]
- Hao, X.; Zhang, J.; Yang, Y.; Wang, H.; Chi, Q. Single-molecule interactions between heme proteins and carboxylic groups in various chemical environments. Chemelectrochem 2020, 7, 4441–4445. [Google Scholar] [CrossRef]
- Le, S.; Yu, M.; Yan, J. Direct single-molecule quantification reveals unexpectedly high mechanical stability of vinculin-talin/alpha-catenin linkages. Sci. Adv. 2019, 5, eaav2720. [Google Scholar] [CrossRef]
- Koo, H.; Park, I.; Lee, Y.; Kim, H.J.; Jung, J.H.; Lee, J.H.; Kim, Y.; Kim, J.H.; Park, J.W. Visualization and quantification of microrna in a single cell using atomic force microscopy. J. Am. Chem. Soc. 2016, 138, 11664–11671. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Woodside, M.T. Mechanical strength of RNA knot in zika virus protects against cellular defenses. Nat. Chem. Biol. 2021, 17, 975–981. [Google Scholar] [CrossRef] [PubMed]
- Tapia-Rojo, R.; Alonso-Caballero, A.; Fernandez, J.M. Talin folding as the tuning fork of cellular mechanotransduction. Proc. Natl. Acad. Sci. USA 2020, 117, 21346–21353. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Caballero, A.; Echelman, D.J.; Tapia-Rojo, R.; Haldar, S.; Eckels, E.C.; Fernandez, J.M. Protein folding modulates the chemical reactivity of a gram-positive adhesin. Nat. Chem. 2021, 13, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Rico-Pasto, M.; Alemany, A.; Ritort, F. Force-dependent folding kinetics of single molecules with multiple intermediates and pathways. J. Phys. Chem. Lett. 2022, 13, 1025–1032. [Google Scholar] [CrossRef]
- Rissone, P.; Bizarro, C.V.; Ritort, F. Stem loop formation drives rna folding in mechanical unzipping experiments. Proc. Natl. Acad. Sci. USA 2022, 119, e2025575119. [Google Scholar] [CrossRef]
- Zhang, X.; Kou, X.; Zhang, W.; Zhang, W. Identification of the new type of g-quadruplex with multiple vacant sites in human telomeric DNA. CCS Chem. 2021, 3, 3192–3204. [Google Scholar] [CrossRef]
- Ma, L.; Xu, M.; Oberhauser, A.F. Naturally occurring osmolytes modulate the nanomechanical properties of polycystic kidney disease domains. J. Biol. Chem. 2010, 285, 38438–38443. [Google Scholar] [CrossRef]
- Bustamante, C.J.; Chemla, Y.R.; Liu, S.; Wang, M.D. Optical tweezers in single-molecule biophysics. Nat. Rev. Methods Primers 2021, 1, 25. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Cai, W.; Cao, N.; Qian, H.-J.; Lu, Z.-Y.; Cui, S. Understanding the extraordinary flexibility of polydimethylsiloxane through single-molecule mechanics. ACS Mater. Lett. 2022, 4, 329–335. [Google Scholar] [CrossRef]
- Liu, Z.; Moreira, R.A.; Dujmović, A.; Liu, H.; Yang, B.; Poma, A.B.; Nash, M.A. Mapping mechanostable pulling geometries of a therapeutic anticalin/ctla-4 protein complex. Nano Lett. 2021, 22, 179–181. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Gao, X.; Li, H. Single molecule force spectroscopy reveals the mechanical design governing the efficient translocation of the bacterial toxin protein rtx. J. Am. Chem. Soc. 2019, 141, 20498–20506. [Google Scholar] [CrossRef] [PubMed]
- Wolny, M.; Batchelor, M.; Bartlett, G.J.; Baker, E.G.; Kurzawa, M.; Knight, P.J.; Dougan, L.; Takagi, Y.; Woolfson, D.N.; Paci, E.; et al. Design and characterization of long and stable de novo single alpha-helix domains. Biophys. J. 2017, 112, 189a. [Google Scholar] [CrossRef]
- Arora, N.; Hazra, J.P.; Rakshit, S. Anisotropy in mechanical unfolding of protein upon partner-assisted pulling and handle-assisted pulling. Commun. Biol. 2021, 4, 925. [Google Scholar] [CrossRef]
- Brockwell, D.J.; Paci, E.; Zinober, R.C.; Beddard, G.S.; Olmsted, P.D.; Smith, D.A.; Perham, R.N.; Radford, S.E. Pulling geometry defines the mechanical resistance of a beta-sheet protein. Nat. Struct. Biol. 2003, 10, 731–737. [Google Scholar] [CrossRef]
- Takahashi, H.; Rico, F.; Chipot, C.; Scheuring, S. Alpha-helix unwinding as force buffer in spectrins. ACS Nano 2018, 12, 2719–2727. [Google Scholar] [CrossRef]
- Ding, Y.; Apostolidou, D.; Marszalek, P. Mechanical stability of a small, highly-luminescent engineered protein nanoluc. Int. J. Mol. Sci. 2021, 22, 55. [Google Scholar] [CrossRef]
- Zhang, X.; Rico, F.; Xu, A.; Moy, V. Atomic force microscopy of protein-protein interactions. In Handbook of Single-Molecule Biophysics; Hinterdorfer, P., Oijen, A., Eds.; Springer: New York, NY, USA, 2009; pp. 555–570. [Google Scholar]
- Tian, F.; Tong, B.; Sun, L.; Shi, S.; Zheng, B.; Wang, Z.; Dong, X.; Zheng, P. N501y mutation of spike protein in SARS-CoV-2 strengthens its binding to receptor ace2. Elife 2021, 10, e69091. [Google Scholar] [CrossRef]
- Hinterdorfer, P.; Baumgartner, W.; Gruber, H.J.; Schilcher, K.; Schindler, H. Detection and localization of individual antibody-antigen recognition events by atomic force microscopy. Proc. Natl. Acad. Sci. USA 1996, 93, 3477–3481. [Google Scholar] [CrossRef] [Green Version]
- Oh, Y.J.; Kohler, M.; Lee, Y.; Mishra, S.; Park, J.W.; Hinterdorfer, P. Label-free probing of binding affinity using topography and recognition imaging. Biophys. J. 2020, 118, 174a–175a. [Google Scholar] [CrossRef]
- Cao, W.; Dong, C.; Kim, S.; Hou, D.; Tai, W.; Du, L.; Im, W.; Zhang, X.F. Biomechanical characterization of SARS-CoV-2 spike rbd and human ace2 protein-protein interaction. Biophys. J. 2021, 120, 1011–1019. [Google Scholar] [CrossRef] [PubMed]
- Bauer, M.S.; Gruber, S.; Hausch, A.; Gomes, P.; Milles, L.F.; Nicolaus, T.; Schendel, L.C.; Navajas, P.L.; Procko, E.; Lietha, D.; et al. A tethered ligand assay to probe sars-cov-2:Ace2 interactions. Proc. Natl. Acad. Sci. USA 2022, 119, e2114397119. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Chen, J.; Li, E.; Hu, C.; Luo, S.Z.; He, C. Ultrahigh adhesion force between silica-binding peptide sb7 and glass substrate studied by single-molecule force spectroscopy and molecular dynamic simulation. Front. Chem. 2020, 8, 600918. [Google Scholar] [CrossRef]
- Wojcikiewicz, E.P.; Abdulreda, M.H.; Zhang, X.H.; Moy, V.T. Force spectroscopy of lfa-1 and its ligands, icam-1 and icam-2. Biomacromolecules 2006, 7, 3188–3195. [Google Scholar] [CrossRef]
- Wang, Y.J.; Rico-Lastres, P.; Lezamiz, A.; Mora, M.; Solsona, C.; Stirnemann, G.; Garcia-Manyes, S. DNA binding induces a nanomechanical switch in the rrm1 domain of tdp-43. J. Phys. Chem. Lett. 2018, 9, 3800–3807. [Google Scholar] [CrossRef]
- Milles, L.F.; Schulten, K.; Gaub, H.E.; Bernardi, R.C. Molecular mechanism of extreme mechanostability in a pathogen adhesin. Science 2018, 359, 1527–1533. [Google Scholar] [CrossRef]
- Prescilla-Ledezma, A.; Linares, F.; Ortega-Muñoz, M.; Retana Moreira, L.; Jódar-Reyes, A.B.; Hernandez-Mateo, F.; Santoyo-Gonzalez, F.; Osuna, A. Molecular recognition of surface trans-sialidases in extracellular vesicles of the parasite trypanosoma cruzi using atomic force microscopy (afm). Int. J. Mol. Sci. 2022, 23, 7193. [Google Scholar] [CrossRef]
- Lipke, P.N.; Rauceo, J.M.; Viljoen, A. Cell-cell mating interactions: Overview and potential of single-cell force spectroscopy. Int. J. Mol. Sci. 2022, 23, 1110. [Google Scholar] [CrossRef]
- Yang, J.; Petitjean, S.J.L.; Koehler, M.; Zhang, Q.; Dumitru, A.C.; Chen, W.; Derclaye, S.; Vincent, S.P.; Soumillion, P.; Alsteens, D. Molecular interaction and inhibition of SARS-CoV-2 binding to the ace2 receptor. Nat. Commun. 2020, 11, 4541. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tian, F.; Shi, S.; Deng, Y.; Zheng, P. Enzymatic protein–protein conjugation through internal site verified at the single-molecule level. J. Phys. Chem. Lett. 2021, 12, 10914–10919. [Google Scholar] [CrossRef] [PubMed]
- Yuan, G.; Curtolo, F.; Deng, Y.; Wu, T.; Tian, F.; Ma, Q.; Liu, Y.; Zuo, J.; Arantes, G.M.; Zheng, P. Highly dynamic polynuclear metal cluster revealed in a single metallothionein molecule. Research 2021, 2021, 9756945. [Google Scholar] [CrossRef] [PubMed]
- Pill, M.F.; East, A.L.L.; Marx, D.; Beyer, M.K.; Clausen-Schaumann, H. Mechanical activation drastically accelerates amide bond hydrolysis, matching enzyme activity. Angew. Chem. Int. Ed. 2019, 58, 9787–9790. [Google Scholar] [CrossRef]
- Zhao, P.; Xu, C.-Q.; Sun, C.; Xia, J.; Sun, L.; Li, J.; Xu, H. Exploring the difference of bonding strength between silver(i) and chalcogenides in block copolymer systems. Polym. Chem. 2020, 11, 7087–7093. [Google Scholar] [CrossRef]
- Lei, H.; Ma, Q.; Li, W.; Wen, J.; Ma, H.; Qin, M.; Wang, W.; Cao, Y. An ester bond underlies the mechanical strength of a pathogen surface protein. Nat. Commun. 2021, 12, 5082. [Google Scholar] [CrossRef]
- López-García, P.; de Araujo, A.D.; Bergues-Pupo, A.E.; Tunn, I.; Fairlie, D.P.; Blank, K.G. Fortified coiled coils: Enhancing mechanical stability with lactam or metal staples. Angew. Chem. Int. Ed. 2021, 60, 232–236. [Google Scholar] [CrossRef]
- Xiang, W.; Li, Z.; Xu, C.-Q.; Li, J.; Zhang, W.; Xu, H. Quantifying the bonding strength of gold-chalcogen bonds in block copolymer systems. Chem. Asian J. 2019, 14, 1481–1486. [Google Scholar] [CrossRef] [PubMed]
- Nie, J.; Deng, Y.; Tian, F.; Shi, S.; Zheng, P. Detection of weak non-covalent cation-π interactions in ngal by single-molecule force spectroscopy. Nano Res. 2022, 15, 4251–4257. [Google Scholar] [CrossRef]
- Rico, F.; Russek, A.; Gonzalez, L.; Grubmuller, H.; Scheuring, S. Heterogeneous and rate-dependent streptavidin-biotin unbinding revealed by high-speed force spectroscopy and atomistic simulations. Proc. Natl. Acad. Sci. USA 2019, 116, 6594–6601. [Google Scholar] [CrossRef]
- Nie, J.Y.; Tian, F.; Zheng, B.; Wang, Z.Y.; Zheng, P. Exploration of metal-ligand coordination bonds in proteins by single-molecule force spectroscopy. Chem. Lett. 2021, 50, 1667–1675. [Google Scholar] [CrossRef]
- Nunes-Alves, A.; Arantes, G.M. Mechanical unfolding of macromolecules coupled to bond dissociation. J. Chem. Theory Comput. 2018, 14, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Pang, X.; Zhao, J.; Zhang, Q.; Shan, Y. Evaluating the single-molecule interactions between targeted peptides and the receptors on living cell membrane. Nanoscale 2021, 13, 17318–17324. [Google Scholar] [CrossRef] [PubMed]
- Razgoniaev, A.O.; Glasstetter, L.M.; Kouznetsova, T.B.; Hall, K.C.; Horst, M.; Craig, S.L.; Franz, K.J. Single-molecule activation and quantification of mechanically triggered palladium–carbene bond dissociation. J. Am. Chem. Soc. 2021, 143, 1784–1789. [Google Scholar] [CrossRef]
- Song, G.; Tian, F.; Liu, H.; Li, G.; Zheng, P. Pioglitazone inhibits metal cluster transfer of mitoneet by stabilizing the labile Fe–N bond revealed at single-bond level. J. Phys. Chem. Lett. 2021, 12, 3860–3867. [Google Scholar] [CrossRef]
- Pang, S.M.; Le, S.; Yan, J. Mechanical responses of the mechanosensitive unstructured domains in cardiac titin. Biol. Cell 2018, 110, 65–76. [Google Scholar] [CrossRef]
- Yuan, G.; Le, S.; Yao, M.; Qian, H.; Zhou, X.; Yan, J.; Chen, H. Elasticity of the transition state leading to an unexpected mechanical stabilization of titin immunoglobulin domains. Angew. Chem. Int. Ed. 2017, 56, 5490–5493. [Google Scholar] [CrossRef]
- Williams, P.M.; Fowler, S.B.; Best, R.B.; Luis Toca-Herrera, J.; Scott, K.A.; Steward, A.; Clarke, J. Hidden complexity in the mechanical properties of titin. Nature 2003, 422, 446. [Google Scholar] [CrossRef]
- Carrion-Vazquez, M.; Oberhauser, A.F.; Fowler, S.B.; Marszalek, P.E.; Broedel, S.E.; Clarke, J.; Fernandez, J.M. Mechanical and chemical unfolding of a single protein: A comparison. Proc. Natl. Acad. Sci. USA 1999, 96, 3694–3699. [Google Scholar] [CrossRef]
- Giganti, D.; Yan, K.; Badilla, C.L.; Fernandez, J.M.; Alegre-Cebollada, J. Disulfide isomerization reactions in titin immunoglobulin domains enable a mode of protein elasticity. Nat. Commun. 2018, 9, 185. [Google Scholar] [CrossRef]
- Popa, I.; Kosuri, P.; Alegre-Cebollada, J.; Garcia-Manyes, S.; Fernandez, J.M. Force dependency of biochemical reactions measured by single-molecule force-clamp spectroscopy. Nat. Protoc. 2013, 8, 1261–1276. [Google Scholar] [CrossRef] [PubMed]
- Bang, M.-L.; Centner, T.; Fornoff, F.; Geach, A.J.; Gotthardt, M.; McNabb, M.; Witt, C.C.; Labeit, D.; Gregorio, C.C.; Granzier, H.; et al. The complete gene sequence of titin, expression of an unusual 700-KDa titin isoform, and its interaction with obscurin identify a novel z-line to i-band linking system. Circ. Res. 2001, 89, 1065–1072. [Google Scholar] [CrossRef] [PubMed]
- Stahl, S.W.; Nash, M.A.; Fried, D.B.; Slutzki, M.; Barak, Y.; Bayer, E.A.; Gaub, H.E. Single-molecule dissection of the high-affinity cohesin-dockerin complex. Proc. Natl. Acad. Sci. USA 2012, 109, 20431–20436. [Google Scholar] [CrossRef]
- Deng, Y.; Wu, T.; Wang, M.; Shi, S.; Yuan, G.; Li, X.; Chong, H.; Wu, B.; Zheng, P. Enzymatic biosynthesis and immobilization of polyprotein verified at the single-molecule level. Nat. Commun. 2019, 10, 2775. [Google Scholar] [CrossRef]
- LeBlanc, M.A.; Fink, M.R.; Perkins, T.T.; Sousa, M.C. Type III secretion system effector proteins are mechanically labile. Proc. Natl. Acad. Sci. USA 2021, 118, e2019566118. [Google Scholar] [CrossRef]
- Hu, X.; Zhao, X.; He, B.; Zhao, Z.; Zheng, Z.; Zhang, P.; Shi, X.; Kwok, R.T.K.; Lam, J.W.Y.; Qin, A.; et al. A simple approach to bioconjugation at diverse levels: Metal-free click reactions of activated alkynes with native groups of biotargets without prefunctionalization. Research 2018, 2018, 3152870. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.; Wang, Z.; Deng, Y.; Tian, F.; Wu, Q.; Zheng, P. Combination of click chemistry and enzymatic ligation for stable and efficient protein immobilization for single-molecule force spectroscopy. CCS Chem. 2022, 4, 598–604. [Google Scholar] [CrossRef]
- Cao, Y.; Lam, C.; Wang, M.; Li, H. Nonmechanical protein can have significant mechanical stability. Angew. Chem. Int. Ed. 2006, 45, 642–645. [Google Scholar] [CrossRef] [PubMed]
- Marko, J.F.; Siggia, E.D. Stretching DNA. Macromolecules 1995, 28, 8759–8770. [Google Scholar] [CrossRef]
- Garg, S.; Singaraju, G.S.; Yengkhom, S.; Rakshit, S. Tailored polyproteins using sequential staple and cut. Bioconjugate Chem. 2018, 29, 1714–1719. [Google Scholar] [CrossRef]
- Tian, F.; Li, G.; Zheng, B.; Liu, Y.; Shi, S.; Deng, Y.; Zheng, P. Verification of sortase for protein conjugation by single-molecule force spectroscopy and molecular dynamics simulations. Chem. Commun. 2020, 56, 3943–3946. [Google Scholar]
- Lv, S.; Dudek, D.M.; Cao, Y.; Balamurali, M.M.; Gosline, J.; Li, H. Designed biomaterials to mimic the mechanical properties of muscles. Nature 2010, 465, 69–73. [Google Scholar] [PubMed]
- Li, H.; Cao, Y. Protein mechanics: From single molecules to functional biomaterials. Acc. Chem. Res. 2010, 43, 1331–1341. [Google Scholar] [PubMed]
- Khoury, L.R.; Slawinski, M.; Collison, D.R.; Popa, I. Cation-induced shape programming and morphing in protein-based hydrogels. Sci. Adv. 2020, 6, eaba6112. [Google Scholar] [CrossRef]
- Hoffmann, T.; Tych, K.M.; Crosskey, T.; Schiffrin, B.; Brockwell, D.J.; Dougan, L. Rapid and robust polyprotein production facilitates single-molecule mechanical characterization of beta-barrel assembly machinery polypeptide transport associated domains. ACS Nano 2015, 9, 8811–8821. [Google Scholar]
- Yang, R.; Wong, Y.H.; Nguyen, G.K.T.; Tam, J.P.; Lescar, J.; Wu, B. Engineering a catalytically efficient recombinant protein ligase. J. Am. Chem. Soc. 2017, 139, 5351–5358. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Polyprotein | 9G Linker | No Linker |
|---|---|---|
| I(27–32) | 308 ± 64 pN (n = 1148) | 324 ± 54 pN (n = 1808) |
| I(28–30) | 325 ± 35 pN (n = 965) | 330 ± 36 pN (n = 860) |
| I(30–32) | 320 ± 33 pN (n = 1120) | 276 pN/345 pN (n = 1804) |
| Polyprotein | 9G Linker | No Linker |
|---|---|---|
| I(27–32) | 186 ± 55 pN (n = 225) | 203 ± 34 pN (n = 360) |
| I(28–30) | 159 ± 23 pN (n = 477) | 175 ± 24 pN (n = 404) |
| I(30–32) | 197 ± 26 pN (n = 570) | 172 ± 30 pN (n = 910) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tong, B.; Tian, F.; Zheng, P. Interdomain Linker Effect on the Mechanical Stability of Ig Domains in Titin. Int. J. Mol. Sci. 2022, 23, 9836. https://doi.org/10.3390/ijms23179836
Tong B, Tian F, Zheng P. Interdomain Linker Effect on the Mechanical Stability of Ig Domains in Titin. International Journal of Molecular Sciences. 2022; 23(17):9836. https://doi.org/10.3390/ijms23179836
Chicago/Turabian StyleTong, Bei, Fang Tian, and Peng Zheng. 2022. "Interdomain Linker Effect on the Mechanical Stability of Ig Domains in Titin" International Journal of Molecular Sciences 23, no. 17: 9836. https://doi.org/10.3390/ijms23179836
APA StyleTong, B., Tian, F., & Zheng, P. (2022). Interdomain Linker Effect on the Mechanical Stability of Ig Domains in Titin. International Journal of Molecular Sciences, 23(17), 9836. https://doi.org/10.3390/ijms23179836