
Improving the Laboratory Diagnosis of M-like Variants Related to Alpha1-Antitrypsin Deficiency

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
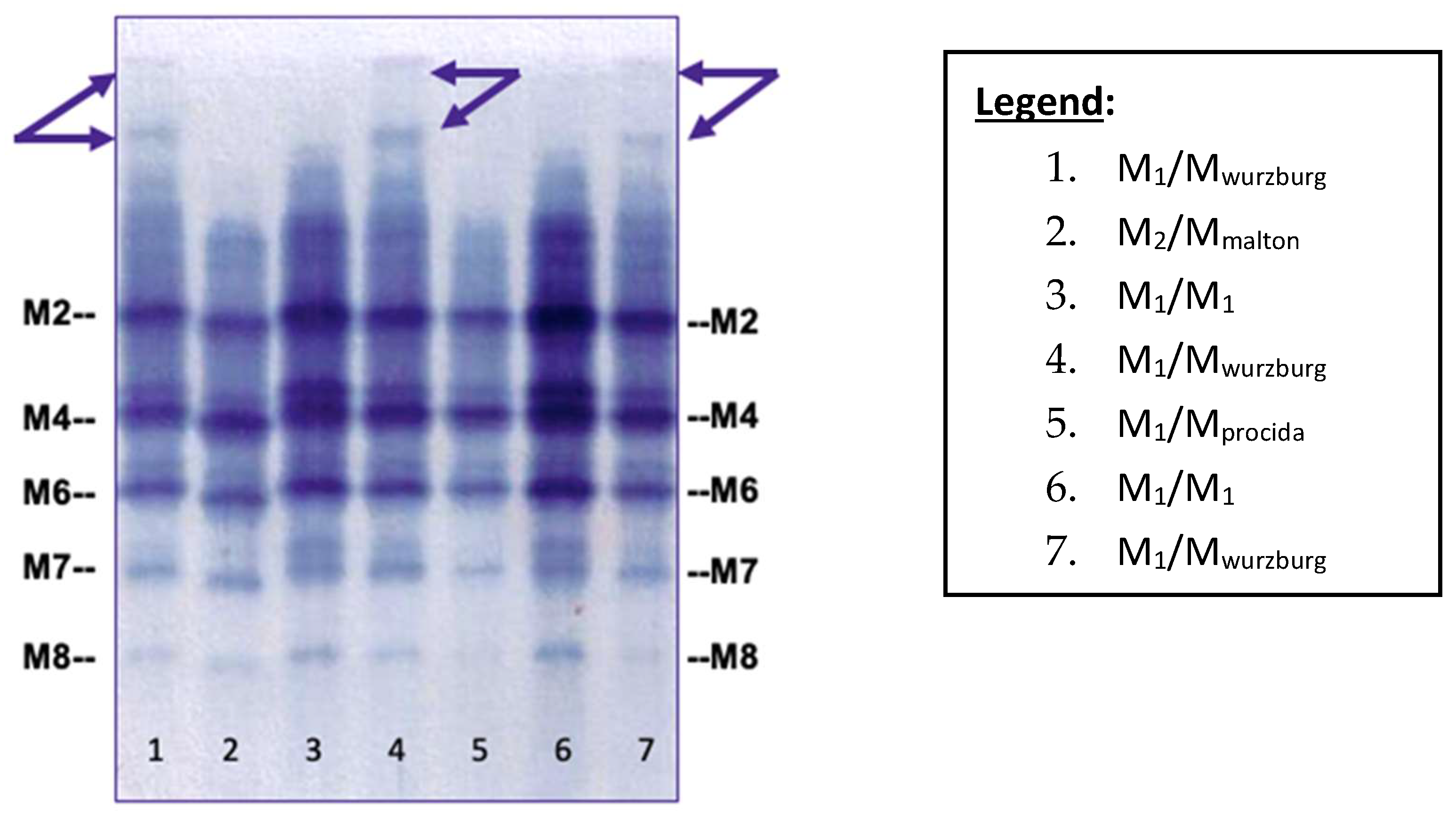
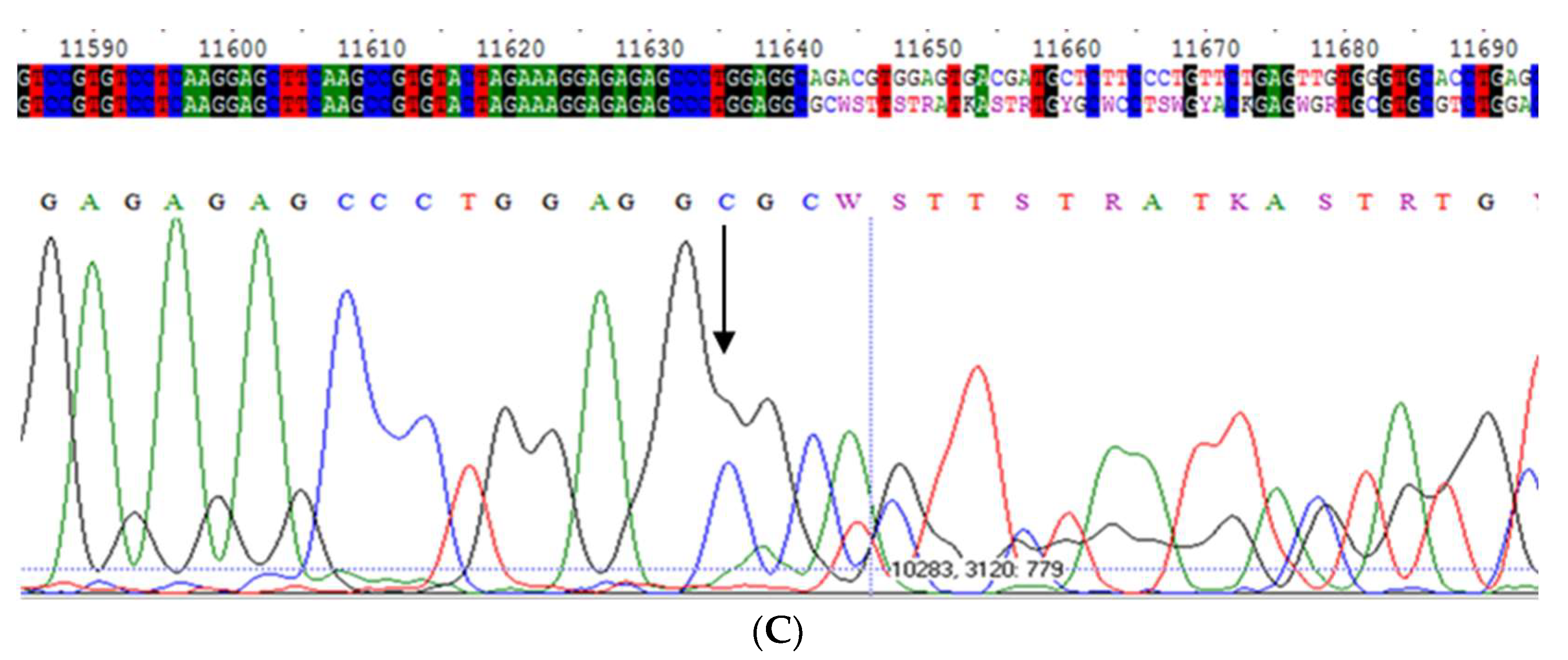
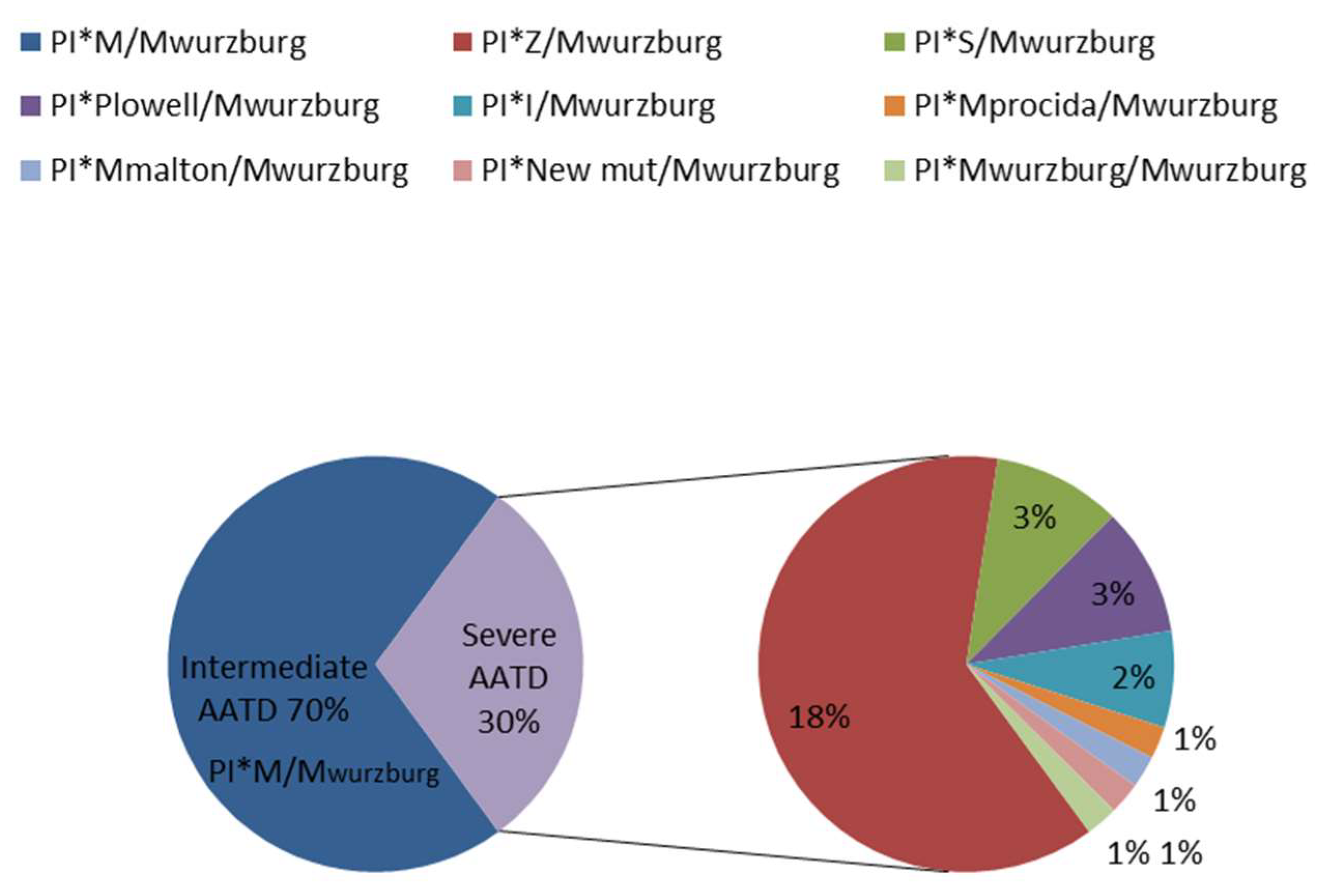
2.1. Mwurzburg
2.2. Mwhitstable
3. Discussion
4. Materials and Methods
4.1. Samples
4.2. Isoelectrophocusing (IEF)
4.3. Sequencing
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Greene, C.M.; Marciniak, S.J.; Teckman, J.; Ferrarotti, I.; Brantly, M.L.; Lomas, D.A.; Stoller, J.K.; McElvaney, N.G. α1-Antitrypsin deficiency. Nat. Rev. Dis. Primers 2016, 2, 16051, Erratum in Nat. Rev. Dis. Primers 2018, 4, 40. [Google Scholar] [CrossRef] [PubMed]
- Torres-Durán, M.; Lopez-Campos, J.L.; Barrecheguren, M.; Miravitlles, M.; Martinez-Delgado, B.; Castillo, S.; Escribano, A.; Baloira, A.; Navarro-Garcia, M.M.; Pellicer, D.; et al. Alpha-1 antitrypsin deficiency: Outstanding questions and future directions. Orphanet J. Rare Dis. 2018, 13, 114. [Google Scholar] [CrossRef] [PubMed]
- Seixas, S.; Marques, P.I. Known Mutations at the Cause of Alpha-1 Antitrypsin Deficiency an Updated Overview of SERPINA1 Variation Spectrum. Appl. Clin. Genet. 2021, 14, 173–194. [Google Scholar] [CrossRef] [PubMed]
- Ferrarotti, I.; Ottaviani, S.; De Silvestri, A.; Corsico, A.G. Update on α1-antitrypsin deficiency. Breathe 2018, 14, e17–e24. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Frias, F.; Miravitlles, M.; Vidal, R.; Camos, S.; Jardi, R. Rare alpha-1-antitrypsin variants: Are they really so rare? Ther. Adv. Respir. Dis. 2012, 6, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Balderacchi, A.M.; Barzon, V.; Ottaviani, S.; Corino, A.; Zorzetto, M.; Wencker, M.; Corsico, A.G.; Ferrarotti, I. Comparison of different algorithms in laboratory diagnosis of alpha1-antitrypsin deficiency. Clin. Chem. Lab. Med. 2021, 59, 1384–1391. [Google Scholar] [CrossRef] [PubMed]
- Cox, D.W.; Johnson, A.M.; Fagerhol, M.K. Report of Nomenclature Meeting for alpha 1-antitrypsin, INSERM, Rouen/Bois-Guillaume-1978. Hum. Genet. 1980, 53, 429–433. [Google Scholar] [CrossRef] [PubMed]
- Poller, W.; Merklein, F.; Schneider-Rasp, S.; Haack, A.; Fechner, H.; Wang, H.; Anagnostopoulos, I.; Weidinger, S. Molecular characterisation of the defective alpha 1-antitrypsin alleles PI Mwurzburg (Pro369Ser), Mheerlen (Pro369Leu), and Q0lisbon (Thr68Ile). Eur. J. Hum. Genet. 1999, 7, 321–331. [Google Scholar] [CrossRef] [PubMed]
- Ambrose, H.J.; Chambers, S.M.; Mieli-Vergani, G.; Ferrie, R.; Newton, C.R.; Robertson, N.H. Molecular characterization of a new alpha-1-antitrypsin M variant allele, Mwhitstable: Implications for DNA-based diagnosis. Diagn. Mol. Pathol. 1999, 8, 205–210. [Google Scholar] [CrossRef]
- Seixas, S.; Lopes, A.I.; Rocha, J.; Silva, L.; Salgueiro, C.; Salazar-de-Sousa, J.; Batista, A. Association between the defective Pro369Ser mutation and in vivo intrahepatic 1-antitrypsin accumulation. J. Med. Genet. 2001, 38, 472–474. [Google Scholar] [CrossRef] [Green Version]
- Zerimech, F.; Hennache, G.; Bellon, F.; Barouh, G.; Lafitte, J.J.; Porchet, N.; Balduyck, M. Evaluation of a new Sebia isoelectrofocusing kit for alpha 1-antitrypsin phenotyping with the Hydrasys System. Clin. Chem. Lab. Med. 2008, 46, 260–263. [Google Scholar] [CrossRef] [PubMed]
- Ottaviani, S.; Barzon, V.; Divizia, L.; Corsico, A.G.; Ferrarotti, I. Alpha-1 antitrypsin phenotype determination using sebia isoelectrofocusing on dried blood spot samples. In Proceedings of the ERS Annual Congress, London, UK, 3–7 September 2016. [Google Scholar]
- Ottaviani, S.; Barzon, V.; Buxens, A.; Gorrini, M.; Larruskain, A.; El Hamss, R.; Balderacchi, A.M.; Corsico, A.G.; Ferrarotti, I. Molecular diagnosis of alpha1-antitrypsin deficiency: A new method based on Luminex technology. J. Clin. Lab. Anal. 2020, 34, e23279. [Google Scholar] [CrossRef] [PubMed]
- Veith, M.; Klemmer, A.; Anton, I.; El Hamss, R.; Rapun, N.; Janciauskiene, S.; Kotke, V.; Herr, C.; Bals, R.; Vogelmeier, C.F.; et al. Diagnosing Alpha-1-Antitrypsin Deficiency Using A PCR/Luminescence-Based Technology. Int. J. Chronic Obstr. Pulm. Dis. 2019, 14, 2535–2542. [Google Scholar] [CrossRef] [PubMed]
- Fra, A.M.; Gooptu, B.; Ferrarotti, I.; Miranda, E.; Scabini, R.; Ronzoni, R.; Benini, F.; Corda, L.; Medicina, D.; Luisetti, M.; et al. Three new alpha1-antitrypsin deficiency variants help to define a C-terminal region regulating conformational change and polymerization. PLoS ONE. 2012, 7, e38405. [Google Scholar] [CrossRef] [PubMed]
- Laffranchi, M.; Berardelli, R.; Ronzoni, R.; Lomas, D.A.; Fra, A. Heteropolymerization of α-1-antitrypsin mutants in cell models mimicking heterozygosity. Hum. Mol. Genet. 2018, 27, 1785–1793. [Google Scholar] [CrossRef] [PubMed]
- Corda, L.; La Piana, G.E.; Bertella, E.; Moretti, G.; Bianchi, L.; Pinelli, V.; Savoldi, G.; Baiardi, P.; Facchetti, F.; Gatta, N.; et al. Population genetic screening for alpha1-antitrypsin deficiency in a high-prevalence area. Respiration 2011, 82, 418–425. [Google Scholar] [CrossRef] [PubMed]
- Denden, S.; Braham, W.; Amri, F.; Lakhdar, R.; Lefranc, G.; Knani, J.; Chibani, J.B.; Khelil, A.H. Biochemical and Molecular Diagnosis of Alpha 1 Antitrypsin Deficiency in A Tunisian Family. Ann. Biol. Clin. 2009, 67, 441–445. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information (NCBI). Available online: https://www.ncbi.nlm.nih.gov/snp/rs61761869#frequency_tab (accessed on 3 August 2022).
- Ruiz-Duque, B.; Bañuls, L.; Reinoso-Arija, R.; Carrasco-Hernandez, L.; Caballero-Eraso, C.; Dasí, F.; Lopez-Campos, J.L. Methodologies for the Determination of Blood Alpha1 Antitrypsin Levels: A Systematic Review. J. Clin. Med. 2021, 10, 5132. [Google Scholar] [CrossRef] [PubMed]
- Gorrini, M.; Ferrarotti, I.; Lupi, A.; Bosoni, T.; Mazzola, P.; Scabini, R.; Campo, I.; Zorzetto, M.; Novazi, F. and Luisetti, M. Validation of a rapid, simple method to measure alpha1-antitrypsin in human dried blood spots. Clin. Chem. 2006, 52, 899–901. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Genotype | AAT (mg/dL) | CRP (mg/dL) | Age at Diagnosis | Smoking Habits (Pack/Year) | Disease |
|---|---|---|---|---|---|---|
| 1 | SMwurzburg | 86 | Not reported | 63 | Current (31) | Healthy |
| 2 | MMwurzburg | 122.5 | 0.11 | 67 | Not reported | Emphysema |
| 3 | MMwurzburg | 123 | 0.11 | 67 | Current (47) | Emphysema |
| 4 | MMwurzburg | 123.2 | 0.34 | 25 | Not reported | Not reported |
| 5 | MMwurzburg | 124 | 0.32 | 64 | Former (30) | Asthma |
| 6 | MMwurzburg | 125 | 0.15 | 61 | Former (16) | Bronchiectasis |
| 7 | MMwurzburg | 125.2 | 0.28 | 27 | Not reported | Not reported |
| 8 | MMwurzburg | 126 | 0.05 | 23 | Not reported | Not reported |
| 9 | MMwurzburg | 126 | 0.14 | 68 | Never | Not reported |
| 10 | MMwurzburg | 131.6 | 0.06 | 77 | Not reported | Not reported |
| 11 | MMwurzburg | 146.3 | 0.62 | 74 | Not reported | Healthy |
| 12 | MMwurzburg | 177.2 | 0.2 | 55 | Never | Asthma, bronchiectasis, dyspnea—hepatitis |
| 13 | MMwurzburg | 180.5 | 0.11 | 45 | Not reported | Not reported |
| 14 | MM | 182 | 0.25 | 78 | Never | Bronchiectasis |
| 15 | MM | 184 | Not reported | 48 | Not reported | Healthy |
| 16 | MMwurzburg | 197.4 | 0.05 | 55 | Not reported | Emphysema |
| 17 | MMwurzburg | 206 | 3.27 | 78 | Not reported | Not reported |
| 18 | MMwurzburg | 315.1 | 0.09 | 64 | Former (30) | Bronchitis |
| Sample | Genotype | AAT (mg/dL) | CRP (mg/dL) | Age at Diagnosis | Smoking Habits (Pack/Year) | Disease |
|---|---|---|---|---|---|---|
| 1 | ZMwhitstable | 48 | 0.1 | 64 | Never | Not reported |
| 2 | M2Mwhitstable | 55.7 | 0.1 | 60 | Not reported | Not reported |
| 3 | M2Mwhitstable | 56 | 0.1 | 70 | Never | Emphysema |
| 4 | ZMwhitstable | 56.2 | 0.11 | 58 | Never | Hepatitis |
| 5 | ZMwhitstable | 59.9 | 0.03 | 25 | Never | Healthy |
| 6 | ZMwhitstable | 60.6 | 0.01 | 35 | Never | Asthma |
| 7 | ZMwhitstable | 62 | 1.4 | 85 | Former (18) | Not reported |
| 8 | M2Mwhitstable | 66 | 0.1 | 55 | Former (10) | Bronchitis, |
| 9 | M1Mwhitstable | 66.3 | 0.28 | 76 | Former (6.3) | Asthma, bronchitis, bronchiectasis |
| 10 | M1Mwhitstable | 68.4 | 0.6 | 73 | Former (18) | Bronchitis |
| 11 | M1Mwhitstable | 70.7 | 0.2 | 64 | Never | Dyspnea-hepatitis |
| 12 | M2Mwhitstable | 72.4 | 0.2 | 46 | Never | Emphysema, pneumothorax |
| 13 | M1Mwhitstable | 74 | Not reported | Not reported | Not reported | Healthy |
| 14 | M1Mwithstable | 77.1 | 0.1 | 38 | Not reported | Chronic bronchitis, allergic asthma |
| 15 | SMwhitstable | 78 | 0 | 15 | Never | Not reported |
| 16 | M1Mwhitstable | 78.3 | 0.06 | 55 | Former (38) | Asthma |
| 17 | M1Mwithstable | 82.1 | 0.01 | 47 | Not reported | Not reported |
| 18 | M1Mwhitstable | 84.5 | 0.23 | 53 | Never | Asthma |
| 19 | M2Mwhitstable | 86 | 0.1 | 10 months | Never | Bronchitis |
| 20 | M1Mwhitstable | 87.5 | 0.1 | 24 | Never | Bronchiectasis |
| 21 | M3Mwhitstable | 88 | 0.1 | 67 | Current (35) | Bronchiectasis, dyspnea, emphysema, bronchitis-hepatitis |
| 22 | MMwhitstable | 89.3 | 0.4 | 61 | Current (45) | Dyspnea |
| 23 | M3Mwhitstable | 89.8 | 0.13 | 71 | Former | Asthma |
| 24 | M1Mwhitstable | 89.9 | 0.004 | 64 | Current (31) | Emphysema-hepatitis |
| 25 | M1Mwhitstable | 90 | 0.13 | 47 | Never | Asthma, emphysema |
| 26 | M1Mwhitstable | 90.7 | 0.1 | 66 | Never | Bronchiectasis |
| 27 | M1Mwhitstable | 91 | Not reported | 62 | Former (40) | Emphysema |
| 28 | M2Mwhitstable | 91.7 | 0.01 | 54 | Former (Not reported) | Bronchitis |
| 29 | M1Mwhitstable | 92.9 | 0 | 37 | Not reported | Not reported |
| 30 | M1Mwhitstable | 93.3 | 0.1 | 73 | Not reported | Emphysema, pulmonary fibrosis |
| 31 | M3Mwhitstable | 93.7 | 0.03 | 3 | Never | Healthy |
| 32 | M2Mwhitstable | 94.6 | 0.12 | 50 | Current (12) | Emphysema |
| 33 | M2Mwhitstable | 96 | 0.2 | 55 | Current (40) | Emphysema |
| 34 | MMwhitstable | 99.6 | 0.13 | 27 | Current (7.4) | Healthy |
| 35 | M1Mwithstable | 103.9 | 0.83 | 72 | Not reported | COPD, emphysema |
| 36 | M3Mwhitstable | 104 | 1 | 64 | Former (Not reported) | Emphysema |
| 37 | M1Mwhitstable | 111.4 | 0.9 | 72 | Former (0.25) | Asthma, bronchiectasis, dyspnea |
| 38 | M2Mwhitstable | 125 | 0.05 | 56 | Current (45) | Bronchitis, emphysema, dyspnea |
| 39 | MMwhitstable | 128.1 | 0.12 | 31 | Current (14) | Healthy |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barzon, V.; Ottaviani, S.; Balderacchi, A.M.; Corino, A.; Piloni, D.; Accordino, G.; Coretti, M.; Mariani, F.; Corsico, A.G.; Ferrarotti, I. Improving the Laboratory Diagnosis of M-like Variants Related to Alpha1-Antitrypsin Deficiency. Int. J. Mol. Sci. 2022, 23, 9859. https://doi.org/10.3390/ijms23179859
Barzon V, Ottaviani S, Balderacchi AM, Corino A, Piloni D, Accordino G, Coretti M, Mariani F, Corsico AG, Ferrarotti I. Improving the Laboratory Diagnosis of M-like Variants Related to Alpha1-Antitrypsin Deficiency. International Journal of Molecular Sciences. 2022; 23(17):9859. https://doi.org/10.3390/ijms23179859
Chicago/Turabian StyleBarzon, Valentina, Stefania Ottaviani, Alice Maria Balderacchi, Alessandra Corino, Davide Piloni, Giulia Accordino, Manuela Coretti, Francesca Mariani, Angelo Guido Corsico, and Ilaria Ferrarotti. 2022. "Improving the Laboratory Diagnosis of M-like Variants Related to Alpha1-Antitrypsin Deficiency" International Journal of Molecular Sciences 23, no. 17: 9859. https://doi.org/10.3390/ijms23179859
APA StyleBarzon, V., Ottaviani, S., Balderacchi, A. M., Corino, A., Piloni, D., Accordino, G., Coretti, M., Mariani, F., Corsico, A. G., & Ferrarotti, I. (2022). Improving the Laboratory Diagnosis of M-like Variants Related to Alpha1-Antitrypsin Deficiency. International Journal of Molecular Sciences, 23(17), 9859. https://doi.org/10.3390/ijms23179859