
Unbalanced Expression of Glutathione Peroxidase 4 and Arachidonate 15-Lipoxygenase Affects Acrosome Reaction and In Vitro Fertilization
 and
and
Abstract
:1. Introduction
2. Results
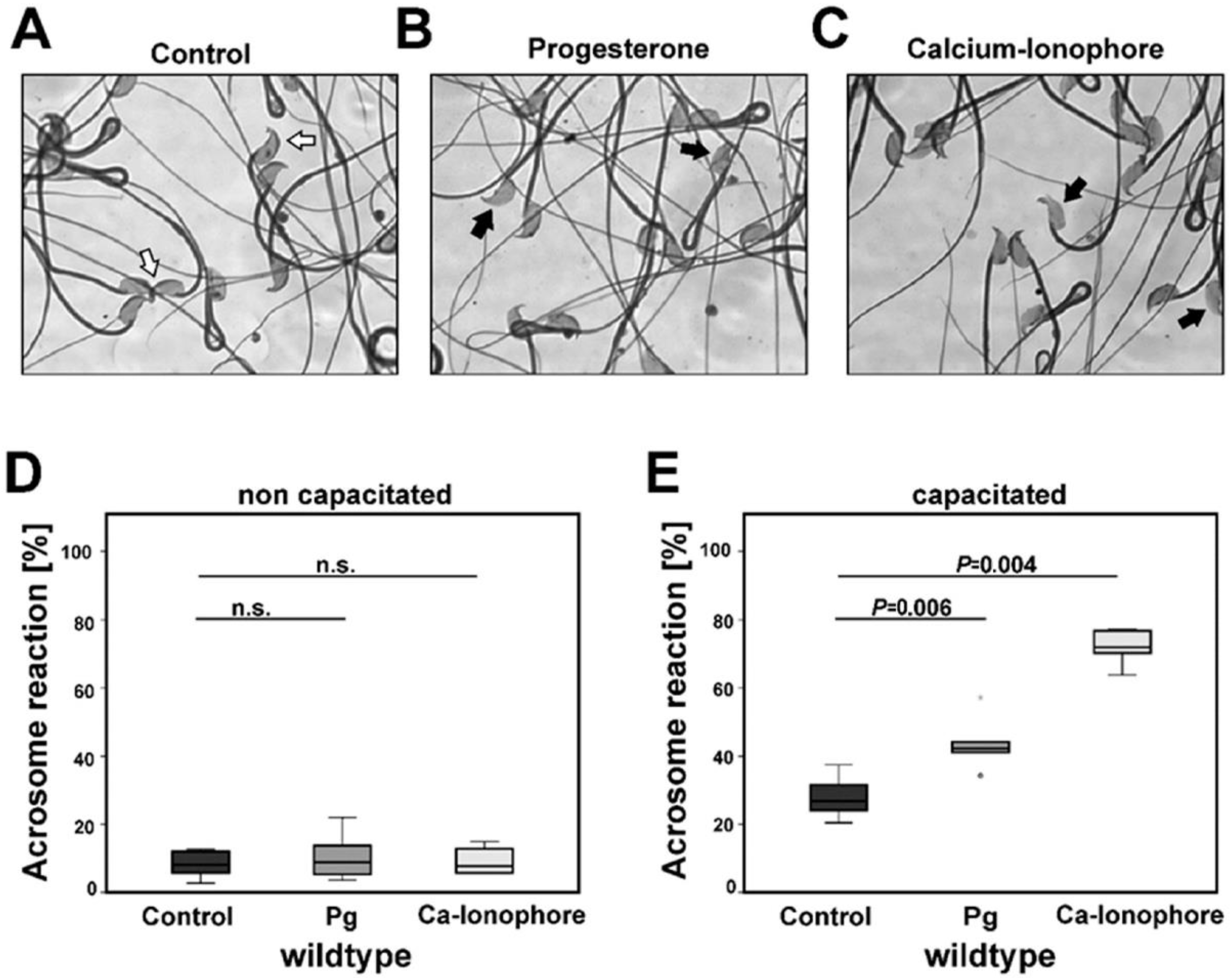
2.1. Capacitation Prepares Isolated Cauda Sperm for the Acrosome Reaction
2.2. Subcellular Distribution of Gpx4 and Alox15 in Non-Capacitated and Capacitated Sperm
2.3. Alox15 Products Are Present in the Cellular Lipids of Mouse Sperm
2.4. Heterozygous Expression of the Catalytically Inactive Sec46Ala Gpx4 Mutant Impairs Acrosomal Exocytosis
2.5. Homozygous Expression of Catalytically Active Gpx4 in Sperm Is Essential for In Vitro Fertilization
2.6. Heterozygous Expression of Catalytically Inactive Gpx4 in Sperm Increases Tyrosine Phosphorylation during In Vitro Capacitation
2.7. Expression of Catalytically Inactive Gpx4+/U46A Mutant and Additional Alox15 Deficiency Induces Spontaneous Acrosome Reaction
2.8. Systemic Inactivation of the Alox15 Gene Normalizes the Increased Degree of Total Protein Tyrosine Phosphorylation That Was Elevated by the Expression of the Catalytically Inactive Gpx4+/U46A Mutant
3. Discussion
4. Materials and Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| A | alanine |
| Ala | alanine |
| Alox15 | mouse arachidonate 15-lipoxygenase |
| AA | 5,8,11,14-eicosatetraensäure |
| DHA | 4,7,10,13,16,19-docosahexaenoic acid |
| DPA | 7,10,13,16,19-docosapentaenoic acid |
| EPA | 5,8,11,14,17-eicosapentaenoic acid |
| Gpx4 | glutathione peroxidase 4 |
| H(p)ETE | hydro(pero)xyeicosatetraenoic acids |
| PUFA | polyenoic fatty acid |
| Sec | selenocysteine |
| Ser | serine |
| U | selenocysteine |
References
- Bhakta, H.H.; Refai, F.H.; Avella, M.A. The molecular mechanisms mediating mammalian fertilization. Development 2019, 146, dev176966. [Google Scholar] [CrossRef]
- Dey, S.; Brothag, C.; Vijayaraghavan, S. Signaling Enzymes Required for Sperm Maturation and Fertilization in Mammals. Front. Cell Dev. Biol. 2019, 7, 341. [Google Scholar] [CrossRef]
- Hirohashi, N.; Yanagimachi, R. Sperm acrosome reaction: Its site and role in fertilization. Biol. Reprod. 2018, 99, 127–133. [Google Scholar] [CrossRef]
- Khawar, M.B.; Gao, H.; Li, W. Mechanism of Acrosome Biogenesis in Mammals. Front. Cell Dev. Biol. 2019, 7, 195. [Google Scholar] [CrossRef]
- Fesahat, F.; Henkel, R.; Agarwal, A. Globozoospermia syndrome: An update. Andrologia 2020, 52, e13459. [Google Scholar] [CrossRef]
- Roldan, E.R.S. Assessments of sperm quality integrating morphology, swimming patterns, bioenergetics and cell signalling. Theriogenology 2020, 150, 388–395. [Google Scholar] [CrossRef]
- Mao, H.-T.; Yang, W.-X. Modes of acrosin functioning during fertilization. Gene 2013, 526, 75–79. [Google Scholar] [CrossRef]
- Ferrer, M.; Rodriguez, H.; Zara, L.; Yu, Y.; Xu, W.; Oko, R. MMP2 and acrosin are major proteinases associated with the inner acrosomal membrane and may cooperate in sperm penetration of the zona pellucida during fertilization. Cell Tissue Res. 2012, 349, 881–895. [Google Scholar] [CrossRef]
- Modelski, M.J.; Menlah, G.; Wang, Y.; Dash, S.; Wu, K.; Galileo, D.S.; Martin-DeLeon, P.A. Hyaluronidase 2: A Novel Germ Cell Hyaluronidase with Epididymal Expression and Functional Roles in Mammalian Sperm1. Biol. Reprod. 2014, 91, 109. [Google Scholar] [CrossRef]
- Ikawa, M.; Inoue, N.; Benham, A.M.; Okabe, M. Fertilization: A sperm’s journey to and interaction with the oocyte. J. Clin. Investig. 2010, 120, 984–994. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.K.; Bansal, P.; Ganguly, A.; Bhandari, B.; Chakrabarti, K. Human zona pellucida glycoproteins: Functional relevance during fertilization. J. Reprod. Immunol. 2009, 83, 50–55. [Google Scholar] [CrossRef]
- Fléchon, J.-E. The acrosome of eutherian mammals. Cell Tissue Res. 2016, 363, 147–157. [Google Scholar] [CrossRef]
- Puga Molina, L.C.; Luque, G.M.; Balestrini, P.A.; Marín-Briggiler, C.I.; Romarowski, A.; Buffone, M.G. Molecular Basis of Human Sperm Capacitation. Front. Cell Dev. Biol. 2018, 6, 72. [Google Scholar] [CrossRef]
- Visconti, P.E.; Moore, G.D.; Bailey, J.L.; Leclerc, P.; Connors, S.A.; Pan, D.; Olds-Clarke, P.; Kopf, G.S. Capacitation of mouse spermatozoa. II. Protein tyrosine phosphorylation and capacitation are regulated by a cAMP-dependent pathway. Development 1995, 121, 1139–1150. [Google Scholar] [CrossRef]
- Osheroff, J.E.; Visconti, P.E.; Valenzuela, J.P.; Travis, A.J.; Alvarez, J.; Kopf, G.S. Regulation of human sperm capacitation by a cholesterol efflux-stimulated signal transduction pathway leading to protein kinase A-mediated up-regulation of protein tyrosine phosphorylation. Mol. Hum. Reprod. 1999, 5, 1017–1026. [Google Scholar] [CrossRef]
- Battistone, M.A.; Alvau, A.; Salicioni, A.M.; Visconti, P.E.; Da Ros, V.G.; Cuasnicu, P.S. Evidence for the involvement of proline-rich tyrosine kinase 2 in tyrosine phosphorylation downstream of protein kinase A activation during human sperm capacitation. Mol. Hum. Reprod. 2014, 20, 1054–1066. [Google Scholar] [CrossRef]
- Fujii, J.; Imai, H. Redox reactions in mammalian spermatogenesis and the potential targets of reactive oxygen species under oxidative stress. Spermatogenesis 2014, 4, e979108. [Google Scholar] [CrossRef]
- Tsunoda, S.; Kimura, N.; Fujii, J. Oxidative stress and redox regulation of gametogenesis, fertilization, and embryonic development. Reprod. Med. Biol. 2014, 13, 71–79. [Google Scholar] [CrossRef]
- Aitken, R.J. Impact of oxidative stress on male and female germ cells: Implications for fertility. Reproduction 2020, 159, R189–R201. [Google Scholar] [CrossRef]
- Fischer, K.A.; Van Leyen, K.; Lovercamp, K.W.; Manandhar, G.; Sutovsky, M.; Feng, D.; Safranski, T. 15-Lipoxygenase is a component of the mammalian sperm cytoplasmic droplet. Reproduction 2005, 130, 213–222. [Google Scholar] [CrossRef]
- Moore, K.; Lovercamp, K.; Feng, D.; Antelman, J.; Sutovsky, M.; Manandhar, G.; van Leyen, K.; Safranski, T.; Sutovsky, P. Altered epididymal sperm maturation and cytoplasmic droplet migration in subfertile male Alox15 mice. Cell Tissue Res. 2010, 340, 569–581. [Google Scholar] [CrossRef] [PubMed]
- Ford, W.C.L. Regulation of sperm function by reactive oxygen species. Hum. Reprod. Updat. 2004, 10, 387–399. [Google Scholar] [CrossRef] [PubMed]
- Qazi, I.H.; Angel, C.; Yang, H.; Zoidis, E.; Pan, B.; Wu, Z.; Ming, Z.; Zeng, C.-J.; Meng, Q.; Han, H.; et al. Role of Selenium and Selenoproteins in Male Reproductive Function: A Review of Past and Present Evidences. Antioxidants 2019, 8, 268. [Google Scholar] [CrossRef] [PubMed]
- Roveri, A.; Ursini, F.; Flohé, L.; Maiorino, M. PHGPx and spermatogenesis. BioFactors 2001, 14, 213–222. [Google Scholar] [CrossRef]
- Ursini, F.; Heim, S.; Kiess, M.; Maiorino, M.; Roveri, A.; Wissing, J.; Flohé, L. Dual Function of the Selenoprotein PHGPx During Sperm Maturation. Science 1999, 285, 1393–1396. [Google Scholar] [CrossRef]
- Ingold, I.; Aichler, M.; Yefremova, E.; Roveri, A.; Buday, K.; Doll, S.; Tasdemir, A.; Hoffard, N.; Wurst, W.; Walch, A.; et al. Expression of a Catalytically Inactive Mutant Form of Glutathione Peroxidase 4 (Gpx4) Confers a Dominant-negative Effect in Male Fertility. J. Biol. Chem. 2015, 290, 14668–14678. [Google Scholar] [CrossRef]
- Brütsch, S.H.; Rademacher, M.; Roth, S.R.; Müller, K.; Eder, S.; Viertel, D.; Franz, C.; Kuhn, H.; Borchert, A. Male Subfertility Induced by Heterozygous Expression of Catalytically Inactive Glutathione Peroxidase 4 Is Rescued in Vivo by Systemic Inactivation of the Alox15 Gene. J. Biol. Chem. 2016, 291, 23578–23588. [Google Scholar] [CrossRef]
- Kühn, H.; Borchert, A. Regulation of enzymatic lipid peroxidation: The interplay of peroxidizing and peroxide reducing enzymes. Free Radic. Biol. Med. 2002, 33, 154–172. [Google Scholar] [CrossRef]
- Schnurr, K.; Borchert, A.; Kuhn, H. Inverse regulation of lipid-peroxidizing and hydroperoxyl lipid-reducing enzymes by interleukins 4 and 13. FASEB J. 1999, 13, 143–154. [Google Scholar] [CrossRef]
- Ivanov, I.; Kuhn, H.; Heydeck, D. Structural and functional biology of arachidonic acid 15-lipoxygenase-1 (ALOX15). Gene 2015, 573, 1–32. [Google Scholar] [CrossRef]
- Singh, N.K.; Rao, G.N. Emerging role of 12/15-Lipoxygenase (ALOX15) in human pathologies. Prog. Lipid Res. 2019, 73, 28–45. [Google Scholar] [CrossRef] [PubMed]
- Brigelius-Flohé, R.; Maiorino, M. Glutathione peroxidases. Biochim. Biophys. Acta 2013, 1830, 3289–3303. [Google Scholar] [CrossRef]
- Schnurr, K.; Belkner, J.; Ursini, F.; Schewe, T.; Kühn, H. The Selenoenzyme Phospholipid Hydroperoxide Glutathione Peroxidase Controls the Activity of the 15-Lipoxygenase with Complex Substrates and Preserves the Specificity of the Oxygenation Products. J. Biol. Chem. 1996, 271, 4653–4658. [Google Scholar] [CrossRef] [PubMed]
- Grossman, S.; Shahin, I.; Sredni, B. Rat testis lipoxygenase-like enzyme characterization of products from linoleic acid. Biochim. Biophys. Acta 1979, 572, 293–297. [Google Scholar] [CrossRef]
- Schon, I.; Sofer, Y.; Cojacaru, M.; Grossman, S. Arachidonic acid metabolism by perfused ram testis. Int. J. Biochem. 1989, 21, 7–13. [Google Scholar] [CrossRef]
- Reddy, G.P.; Prasad, M.; Sailesh, S.; Kumar, Y.V.K.; Reddanna, P. The production of arachidonic acid metabolites in rat testis. Prostaglandins 1992, 44, 497–507. [Google Scholar] [CrossRef]
- Lax, Y.; Grossman, S.; Rubinstein, S.; Magid, N.; Breitbart, H. Role of lipoxygenase in the mechanism of acrosome reaction in mammalian spermatozoa. Biochim. Biophys. Acta 1990, 1043, 12–18. [Google Scholar] [CrossRef]
- Breitbart, H. The biochemistry of the acrosome reaction. Mol. Hum. Reprod. 1997, 3, 195–202. [Google Scholar] [CrossRef]
- Sun, D.; Funk, C.D. Disruption of 12/15-lipoxygenase expression in peritoneal macrophages. Enhanced utilization of the 5-lipoxygenase pathway and diminished oxidation of low density lipoprotein. J. Biol. Chem. 1996, 271, 24055–24062. [Google Scholar] [CrossRef]
- Chen, X.-S.; Shelter, J.R.; Johnson, E.N.; Funk, C. Role of leukotrienes revealed by targeted disruption of the 5-lipoxygenase gene. Nature 1994, 372, 179–182. [Google Scholar] [CrossRef]
- Johnson, E.N.; Brass, L.F.; Funk, C.D. Increased platelet sensitivity to ADP in mice lacking platelet-type 12-lipoxygenase. Proc. Natl. Acad. Sci. USA 1998, 95, 3100–3105. [Google Scholar] [CrossRef] [PubMed]
- Dreher, I.; Schmutzler, C.; Jakob, F.; Köhrle, J. Expression of Selenoproteins in Various Rat and Human Tissues and Cell Lines. J. Trace Elem. Med. Biol. 1997, 11, 83–91. [Google Scholar] [CrossRef]
- Roveri, A.; Casasco, A.; Maiorino, M.; Dalan, P.; Calligaro, A.; Ursini, F. Phospholipid hydroperoxide glutathione peroxidase of rat testis. Gonadotropin dependence and immunocytochemical identification. J. Biol. Chem. 1992, 267, 6142–6146. [Google Scholar] [CrossRef]
- Diaconu, M.S.; Tangat, Y.; Böhm, D.; Kühn, H.; Michelmann, H.W.; Schreiber, G.; Haidl, G.; Glander, H.-J.; Engel, W.; Nayernia, K. Failure of phospholipid hydroperoxide glutathione peroxidase expression in oligoasthenozoospermia and mutations in the PHGPx gene. Andrologia 2006, 38, 152–157. [Google Scholar] [CrossRef]
- Puglisi, R.; Maccari, I.; Pipolo, S.; Conrad, M.; Mangia, F.; Boitani, C. The nuclear form of glutathione peroxidase 4 is associated with sperm nuclear matrix and is required for proper paternal chromatin decondensation at fertilization. J. Cell. Physiol. 2012, 227, 1420–1427. [Google Scholar] [CrossRef]
- Imai, H.; Hirao, F.; Sakamoto, T.; Sekine, K.; Mizukura, Y.; Saito, M.; Kitamoto, T.; Hayasaka, M.; Hanaoka, K.; Nakagawa, Y. Early embryonic lethality caused by targeted disruption of the mouse PHGPx gene. Biochem. Biophys. Res. Commun. 2003, 305, 278–286. [Google Scholar] [CrossRef]
- Yant, L.J.; Ran, Q.; Rao, L.; Van Remmen, H.; Shibatani, T.; Belter, J.G.; Motta, L.; Richardson, A.; Prolla, T.A. The selenoprotein GPX4 is essential for mouse development and protects from radiation and oxidative damage insults. Free Radic. Biol. Med. 2003, 34, 496–502. [Google Scholar] [CrossRef]
- Brütsch, S.H.; Wang, C.C.; Li, L.; Stender, H.; Neziroglu, N.; Richter, C.; Kuhn, H.; Borchert, A. Expression of Inactive Glutathione Peroxidase 4 Leads to Embryonic Lethality, and Inactivation of the Alox15 Gene Does Not Rescue Such Knock-In Mice. Antioxid. Redox Signal. 2015, 22, 281–293. [Google Scholar] [CrossRef]
- Imai, H.; Hakkaku, N.; Iwamoto, R.; Suzuki, J.; Suzuki, T.; Tajima, Y.; Konishi, K.; Minami, S.; Ichinose, S.; Ishizaka, K.; et al. Depletion of Selenoprotein GPx4 in Spermatocytes Causes Male Infertility in Mice. J. Biol. Chem. 2009, 284, 32522–32532. [Google Scholar] [CrossRef]
- Freire-Moar, J.; Alavi-Nassab, A.; Ng, M.; Mulkins, M.; Sigal, E. Cloning and characterization of a murine macrophage lipoxygenase. Biochim. Biophys. Acta 1995, 1254, 112–116. [Google Scholar] [CrossRef]
- Kutzner, L.; Goloshchapova, K.; Heydeck, D.; Stehling, S.; Kuhn, H.; Schebb, N.H. Mammalian ALOX15 orthologs exhibit pronounced dual positional specificity with docosahexaenoic acid. Biochim. Biophys. Acta 2017, 1862, 666–675. [Google Scholar] [CrossRef] [PubMed]
- Baskaran, S.; Finelli, R.; Agarwal, A.; Henkel, R. Reactive oxygen species in male reproduction: A boon or a bane? Andrologia 2021, 53, e13577. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J.; Drevet, J.R. The Importance of Oxidative Stress in Determining the Functionality of Mammalian Spermatozoa: A Two-Edged Sword. Antioxidants 2020, 9, 111. [Google Scholar] [CrossRef]
- Tramer, F.; Micali, F.; Sandri, G.; Bertoni, A.; Lenzi, A.; Gandini, L.; Panfili, E. Enzymatic and immunochemical evaluation of phospholipid hydroperoxide glutathione peroxidase (PHGPx) in testes and epididymal spermatozoa of rats of different ages. Int. J. Androl. 2002, 25, 72–83. [Google Scholar] [CrossRef]
- Haraguchi, C.M.; Mabuchi, T.; Hirata, S.; Shoda, T.; Yamada, A.; Hoshi, K.; Yokota, S. Spatiotemporal Changes of Levels of a Moonlighting Protein, Phospholipid Hydroperoxide Glutathione Peroxidase, in Subcellular Compartments During Spermatogenesis in the Rat Testis. Biol. Reprod. 2003, 69, 885–895. [Google Scholar] [CrossRef]
- Kwon, W.-S.; Rahman, S.; Lee, J.-S.; Kim, J.; Yoon, S.-J.; Park, Y.-J.; You, Y.-A.; Hwang, S.; Pang, M.-G. A comprehensive proteomic approach to identifying capacitation related proteins in boar spermatozoa. BMC Genom. 2014, 15, 897. [Google Scholar] [CrossRef]
- Petit, F.M.; Serres, C.; Bourgeon, F.; Pineau, C.; Auer, J. Identification of sperm head proteins involved in zona pellucida binding. Hum. Reprod. 2013, 28, 852–865. [Google Scholar] [CrossRef]
- Petit, F.M.; Serres, C.; Auer, J. Moonlighting proteins in sperm–egg interactions. Biochem. Soc. Trans. 2014, 42, 1740–1743. [Google Scholar] [CrossRef]
- Flohé, L. Selenium in mammalian spermiogenesis. Biol. Chem. 2007, 388, 987–995. [Google Scholar] [CrossRef]
- Scheerer, P.; Borchert, A.; Krauß, N.; Wessner, H.; Gerth, C.; Höhne, A.; Kuhn, H. Structural Basis for Catalytic Activity and Enzyme Polymerization of Phospholipid Hydroperoxide Glutathione Peroxidase-4 (GPx4). Biochemistry 2007, 46, 9041–9049. [Google Scholar] [CrossRef]
- Borchert, A.; Kalms, J.; Roth, S.R.; Rademacher, M.; Schmidt, A.; Holzhutter, H.-G.; Kuhn, H.; Scheerer, P. Crystal structure and functional characterization of selenocysteine-containing glutathione peroxidase 4 suggests an alternative mechanism of peroxide reduction. Biochim. Biophys. Acta 2018, 1863, 1095–1107. [Google Scholar] [CrossRef] [PubMed]
- Klotz, L.-O.; Sánchez-Ramos, C.; Prieto-Arroyo, I.; Urbánek, P.; Steinbrenner, H.; Monsalve, M. Redox regulation of FoxO transcription factors. Redox Biol. 2015, 6, 51–72. [Google Scholar] [CrossRef]
- Buelna-Chontal, M.; Zazueta, C. Redox activation of Nrf2 & NF-κB: A double end sword? Cell. Signal. 2013, 25, 2548–2557. [Google Scholar] [CrossRef] [PubMed]
- Collodel, G.; Castellini, C.; Lee, J.C.-Y.; Signorini, C. Relevance of Fatty Acids to Sperm Maturation and Quality. Oxidative Med. Cell. Longev. 2020, 2020, 7038124. [Google Scholar] [CrossRef] [PubMed]
- Nowicka-Bauer, K.; Nixon, B. Molecular Changes Induced by Oxidative Stress that Impair Human Sperm Motility. Antioxidants 2020, 9, 134. [Google Scholar] [CrossRef]
- Flesch, F.M.; Gadella, B.M. Dynamics of the mammalian sperm plasma membrane in the process of fertilization. Biochim. Biophys. Acta 2000, 1469, 197–235. [Google Scholar] [CrossRef]
- Aitken, R.J. Reactive oxygen species as mediators of sperm capacitation and pathological damage. Mol. Reprod. Dev. 2017, 84, 1039–1052. [Google Scholar] [CrossRef]
- Donà, G.; Fiore, C.; Andrisani, A.; Ambrosini, G.; Brunati, A.; Ragazzi, E.; Armanini, D.; Bordin, L.; Clari, G. Evaluation of correct endogenous reactive oxygen species content for human sperm capacitation and involvement of the NADPH oxidase system. Hum. Reprod. 2011, 26, 3264–3273. [Google Scholar] [CrossRef]
- Naz, R.K.; Rajesh, P.B. Role of tyrosine phosphorylation in sperm capacitation / acrosome reaction. Reprod. Biol. Endocrinol. 2004, 2, 75. [Google Scholar] [CrossRef]
- Aitken, R.J.; Paterson, M.; Fisher, H.; Buckingham, D.; van Duin, M. Redox regulation of tyrosine phosphorylation in human spermatozoa and its role in the control of human sperm function. J. Cell Sci. 1995, 108 Pt 5, 2017–2025. [Google Scholar] [CrossRef]
- Nadel, J.A.; Conrad, D.J.; Ueki, I.F.; Schuster, A.; Sigal, E. Immunocytochemical localization of arachidonate 15-lipoxygenase in erythrocytes, leukocytes, and airway cells. J. Clin. Investig. 1991, 87, 1139–1145. [Google Scholar] [CrossRef] [PubMed]
- Conrad, D.J. The arachidonate 12/15 lipoxygenases. A review of tissue expression and biologic function. Clin. Rev. Allergy Immunol. 1999, 17, 71–89. [Google Scholar] [CrossRef] [PubMed]
- Conrad, D.J.; Kuhn, H.; Mulkins, M.; Highland, E.; Sigal, E. Specific inflammatory cytokines regulate the expression of human monocyte 15-lipoxygenase. Proc. Natl. Acad. Sci. USA 1992, 89, 217–221. [Google Scholar] [CrossRef] [PubMed]
- Roy, B.; Cathcart, M.K. Induction of 15-Lipoxygenase Expression by IL-13 Requires Tyrosine Phosphorylation of Jak2 and Tyk2 in Human Monocytes. J. Biol. Chem. 1998, 273, 32023–32029. [Google Scholar] [CrossRef]
- Ostareck-Lederer, A.; Ostareck, D.H.; Standart, N.; Thiele, B.J. Translation of 15-lipoxygenase mRNA is inhibited by a protein that binds to a repeated sequence in the 3′ untranslated region. EMBO J. 1994, 13, 1476–1481. [Google Scholar] [CrossRef]
- Ostareck, D.H.; Ostareck-Lederer, A.; Wilm, M.; Thiele, B.J.; Mann, M.; Hentze, M.W. mRNA Silencing in Erythroid Differentiation: hnRNP K and hnRNP E1 Regulate 15-Lipoxygenase Translation from the 3′ End. Cell 1997, 89, 597–606. [Google Scholar] [CrossRef]
- Rapoport, S.M.; Schewe, T. The maturational breakdown of mitochondria in reticulocytes. Biochim. Biophys. Acta 1986, 864, 471–495. [Google Scholar] [CrossRef]
- Schewe, T.; Rapoport, S.M.; Kühn, H. Enzymology and Physiology of Reticulocyte Lipoxygenase: Comparison with Other Lipoxygenases. Adv. Enzymol. Relat. Areas Mol. Biol. 1986, 58, 191–272. [Google Scholar] [CrossRef]
- Walters, J.L.H.; De Iuliis, G.; Dun, M.; Aitken, R.J.; McLaughlin, E.; Nixon, B.; Bromfield, E.G. Pharmacological inhibition of arachidonate 15-lipoxygenase protects human spermatozoa against oxidative stress. Biol. Reprod. 2018, 98, 784–794. [Google Scholar] [CrossRef]
- Rahban, R.; Nef, S. CatSper: The complex main gate of calcium entry in mammalian spermatozoa. Mol. Cell. Endocrinol. 2020, 518, 110951. [Google Scholar] [CrossRef]
- Breitbart, H.; Rubinstein, S.; Lax, Y. Regulatory mechanisms in acrosomal exocytosis. Rev. Reprod. 1997, 2, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Fujiwara, E.; Kakiuchi, Y.; Okabe, M.; Satouh, Y.; Baba, S.A.; Chiba, K.; Hirohashi, N. Most fertilizing mouse spermatozoa begin their acrosome reaction before contact with the zona pellucida during in vitro fertilization. Proc. Natl. Acad. Sci. USA 2011, 108, 4892–4896. [Google Scholar] [CrossRef] [PubMed]
- Hino, T.; Muro, Y.; Tamura-Nakano, M.; Okabe, M.; Tateno, H.; Yanagimachi, R. The Behavior and Acrosomal Status of Mouse Spermatozoa In Vitro, and Within the Oviduct During Fertilization after Natural Mating. Biol. Reprod. 2016, 95, 50. [Google Scholar] [CrossRef] [PubMed]
- Borchert, A.; Küttner, G.; Gießmann, E.; Wang, C.C.; Wessner, H.; Volkmer, R.; Höhne, W.; Kuhn, H. Defining the immunoreactive epitope for the monoclonal anti-human glutathione peroxidase-4 antibody anti-hGPx4 Mab63-1. Immunol. Lett. 2010, 133, 85–93. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Ostermann, A.I.; Willenberg, I.; Schebb, N.H. Comparison of sample preparation methods for the quantitative analysis of eicosanoids and other oxylipins in plasma by means of LC-MS/MS. Anal. Bioanal. Chem. 2015, 407, 1403–1414. [Google Scholar] [CrossRef]
- Willenberg, I.; Ostermann, A.I.; Schebb, N.H. Targeted metabolomics of the arachidonic acid cascade: Current state and challenges of LC–MS analysis of oxylipins. Anal. Bioanal. Chem. 2015, 407, 2675–2683. [Google Scholar] [CrossRef]
- Kolbe, T.; Landsberger, A.; Manz, S.; Na, E.; Urban, I.; Michel, G. Productivity of superovulated C57BL/6J oocyte donors at different ages. Lab Anim. 2015, 44, 346–349. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analyte | Internal Standard (IS) | Retention Time (min) | Molecular Mass (g/mol) | Mass Transition | LOD | Calibration Range (nM) | ||
|---|---|---|---|---|---|---|---|---|
| Q1 (m/z) | Q3 (m/z) | (nM) | Lower | Upper | ||||
| 12-HETE | 12(S)-HETE-d8 | 14.4 | 320.5 | 319.2 | 179.2 | 0.3 | 3 | 2000 |
| 14-HDHA | 15(S)-HETE-d8 | 14.2 | 344.5 | 343.2 | 205.2 | 0.3 | 1 | 2000 |
| 9-HOTrE | 13(S)-HODE-d4 | 12.4 | 294.4 | 293.2 | 171.2 | 0.3 | 1 | 2000 |
| 13-HOTrE | 13(S)-HODE-d4 | 12.6 | 294.4 | 293.2 | 195.1 | 1 | 3 | 2000 |
| 14-HPHA | 12(S)-HETE-d8 | 14.9 | 346.5 | 345.2 | 205.2 | 3 | 6 | 6000 |
| 13(S)-HODE-d4 | - | 13.6 | 300.5 | 299.5 | 198.1 | - | - | - |
| 12(S)-HETE-d8 | - | 14.4 | 328.2 | 327.2 | 184.2 | - | - | - |
| 15(S)-HETE-d8 | - | 14.0 | 328.2 | 327.2 | 226.2 | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soria-Tiedemann, M.; Michel, G.; Urban, I.; Aldrovandi, M.; O’Donnell, V.B.; Stehling, S.; Kuhn, H.; Borchert, A. Unbalanced Expression of Glutathione Peroxidase 4 and Arachidonate 15-Lipoxygenase Affects Acrosome Reaction and In Vitro Fertilization. Int. J. Mol. Sci. 2022, 23, 9907. https://doi.org/10.3390/ijms23179907
Soria-Tiedemann M, Michel G, Urban I, Aldrovandi M, O’Donnell VB, Stehling S, Kuhn H, Borchert A. Unbalanced Expression of Glutathione Peroxidase 4 and Arachidonate 15-Lipoxygenase Affects Acrosome Reaction and In Vitro Fertilization. International Journal of Molecular Sciences. 2022; 23(17):9907. https://doi.org/10.3390/ijms23179907
Chicago/Turabian StyleSoria-Tiedemann, Mariana, Geert Michel, Iris Urban, Maceler Aldrovandi, Valerie B. O’Donnell, Sabine Stehling, Hartmut Kuhn, and Astrid Borchert. 2022. "Unbalanced Expression of Glutathione Peroxidase 4 and Arachidonate 15-Lipoxygenase Affects Acrosome Reaction and In Vitro Fertilization" International Journal of Molecular Sciences 23, no. 17: 9907. https://doi.org/10.3390/ijms23179907
APA StyleSoria-Tiedemann, M., Michel, G., Urban, I., Aldrovandi, M., O’Donnell, V. B., Stehling, S., Kuhn, H., & Borchert, A. (2022). Unbalanced Expression of Glutathione Peroxidase 4 and Arachidonate 15-Lipoxygenase Affects Acrosome Reaction and In Vitro Fertilization. International Journal of Molecular Sciences, 23(17), 9907. https://doi.org/10.3390/ijms23179907