
Aerobic Exercise Alleviates Abnormal Autophagy in Brain Cells of APP/PS1 Mice by Upregulating AdipoR1 Levels

Abstract
:1. Introduction
2. Results
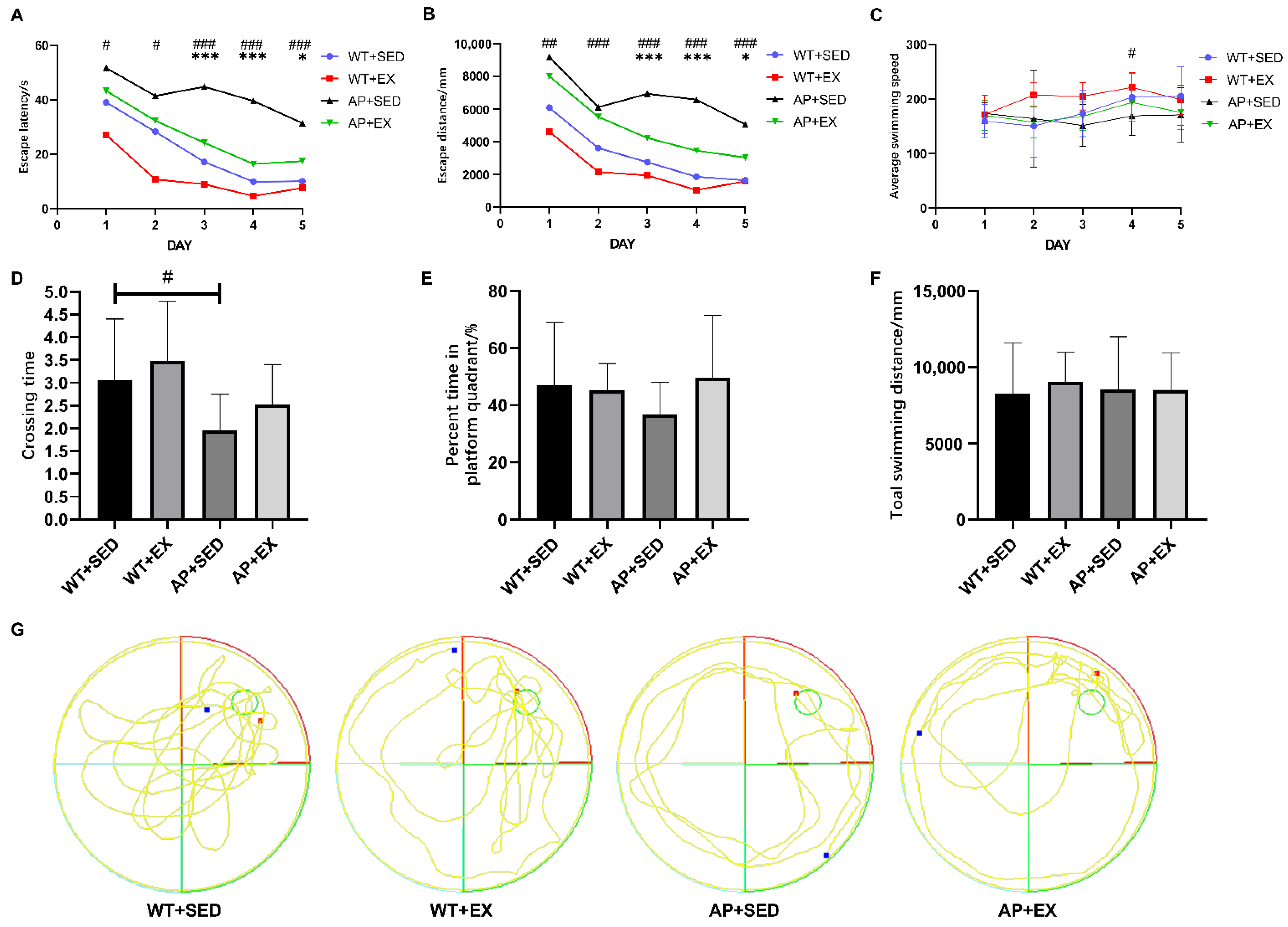
2.1. Exercise Alleviates Cognitive Impairment in AD Mice
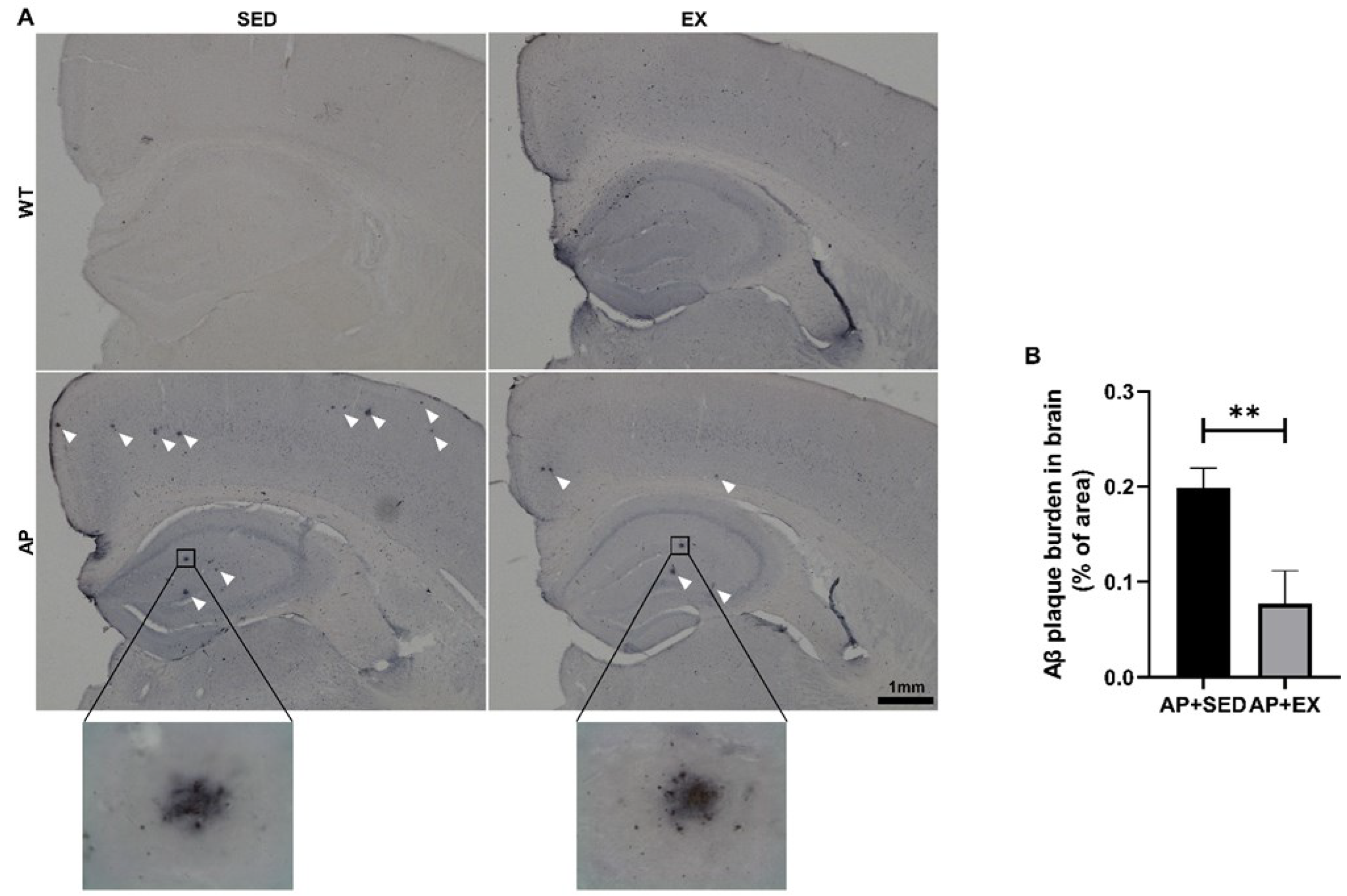
2.2. Exercise Reduces Aβ Accumulation in the Brains of AD Mice
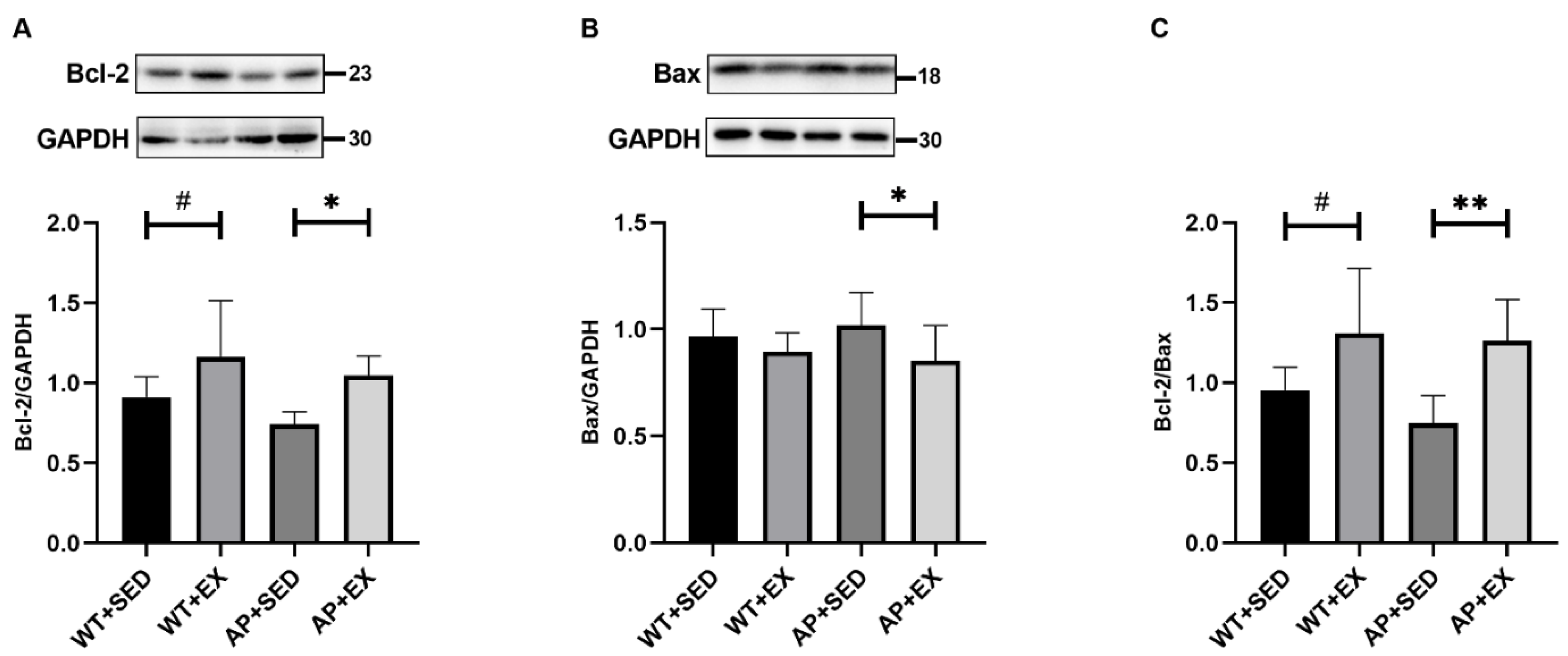
2.3. Exercise Inhibits Brain Cell Apoptosis in AD Mice
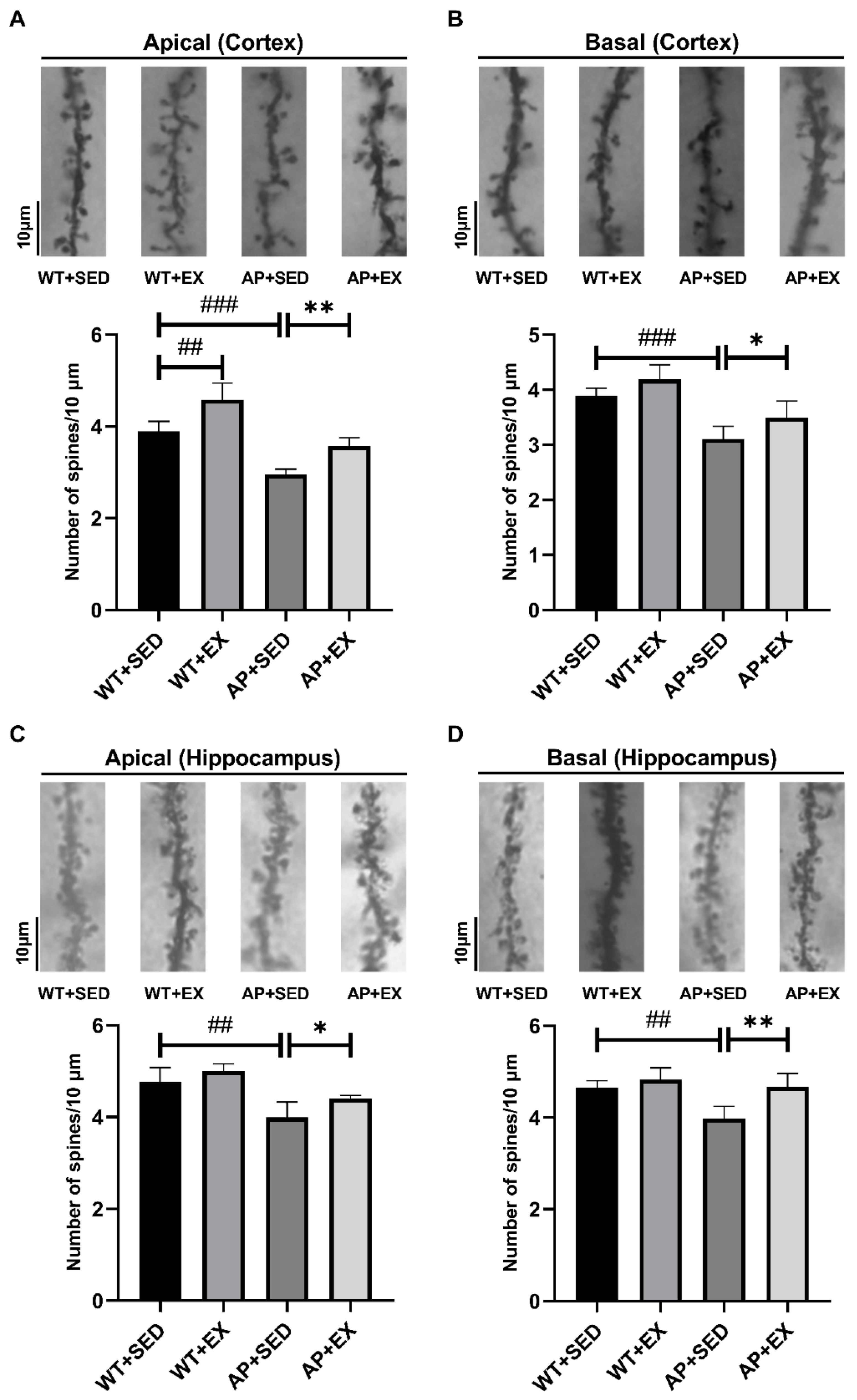
2.4. Exercise Increases the Density of Dendritic Spines in AD Mice
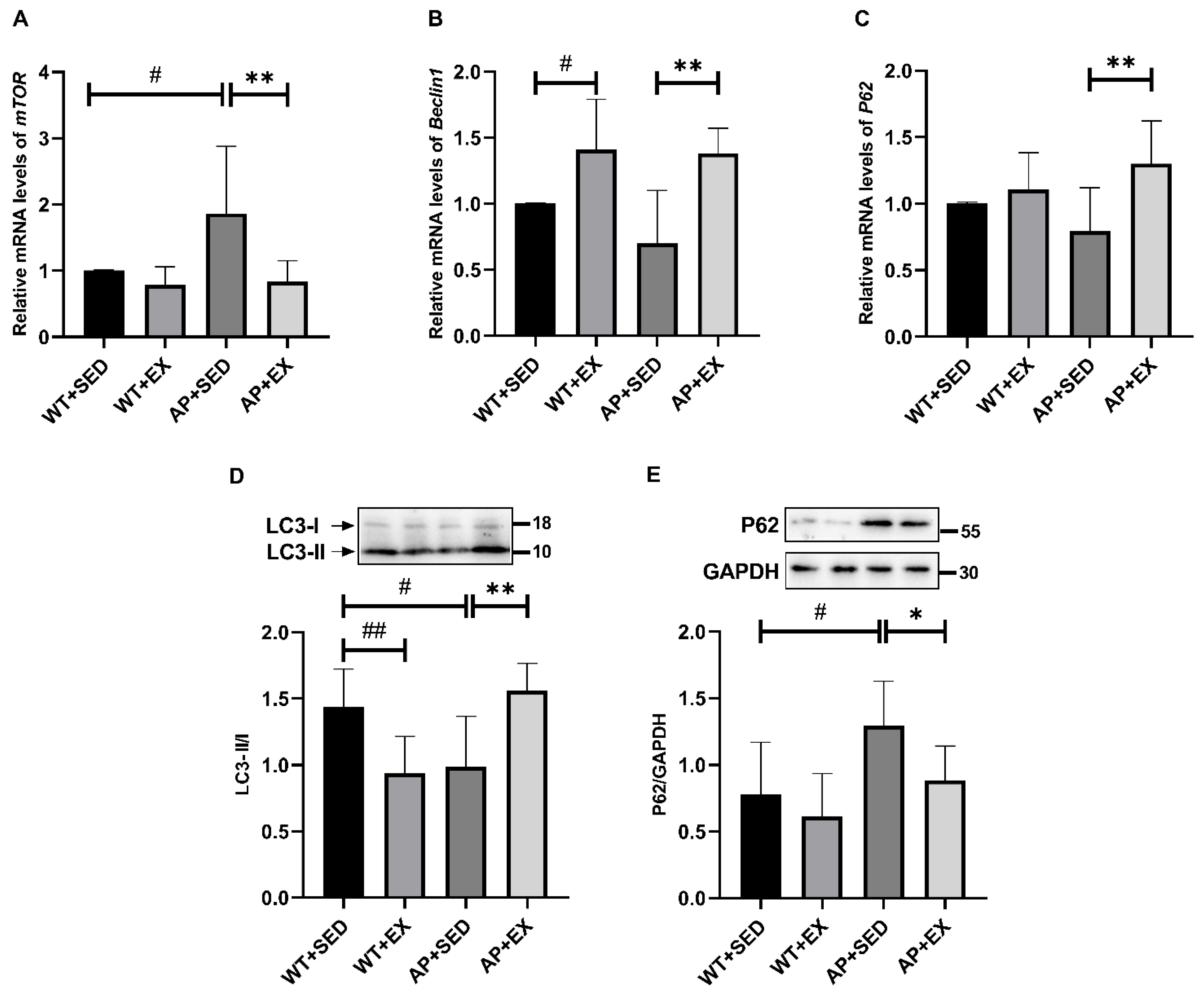
2.5. Exercise Alleviates Abnormal Autophagy in the Brain Cells of AD Mice
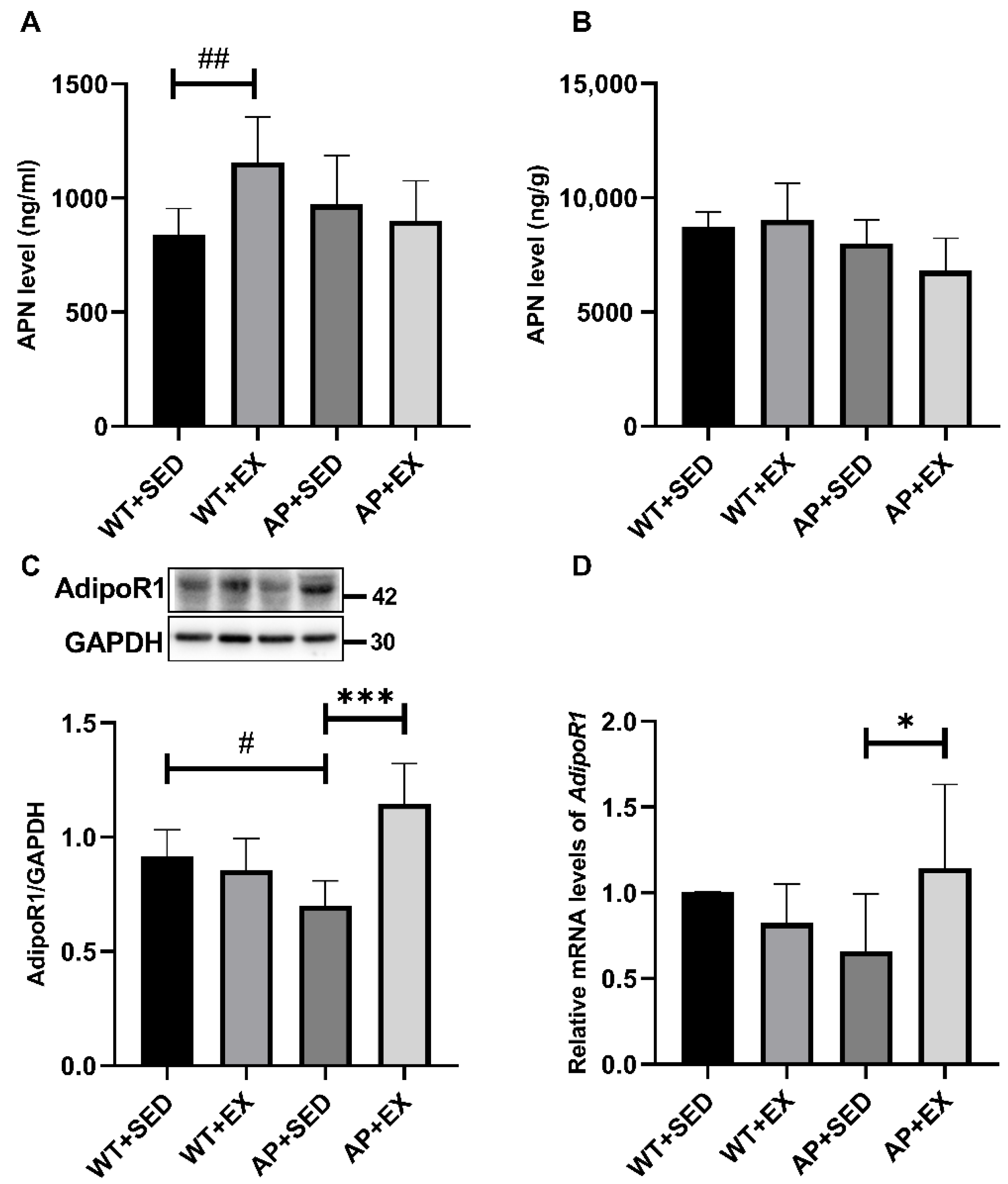
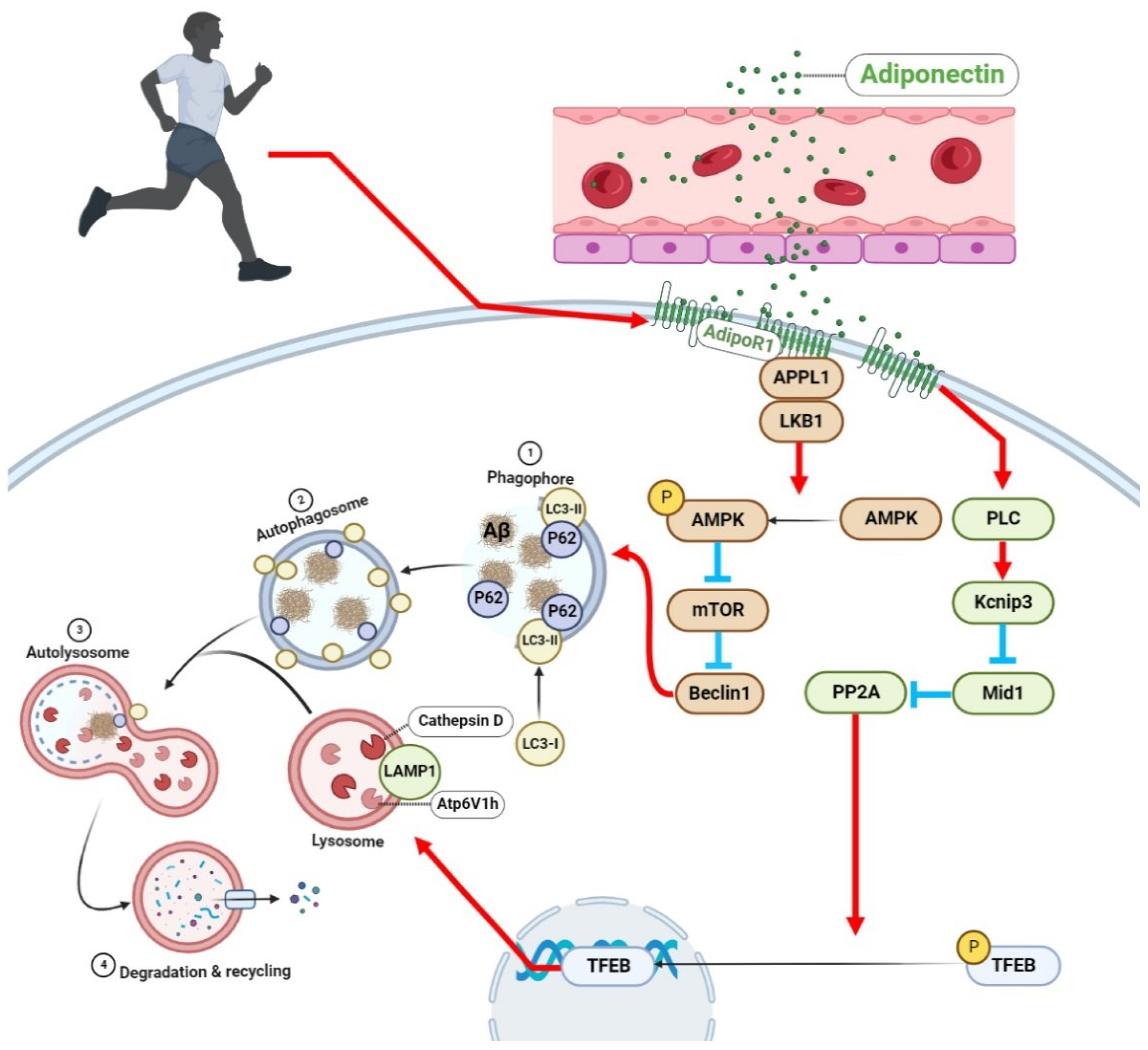
2.6. The Effect of Exercise on APN and Its Receptors in the Brains of AD Mice
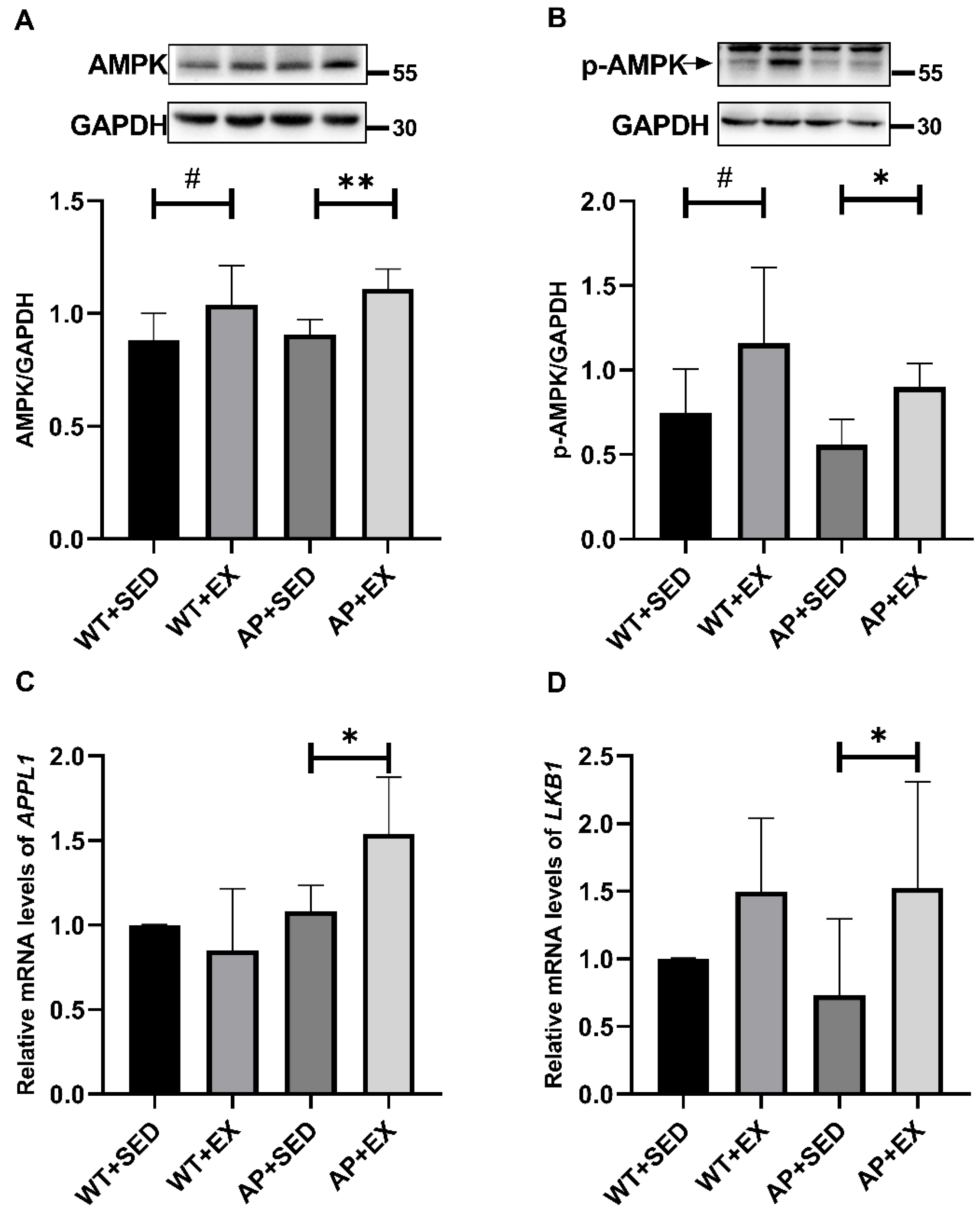
2.7. Exercise Promotes the Expression and Phosphorylation of AMPK in the Brain Cells of AD Mice
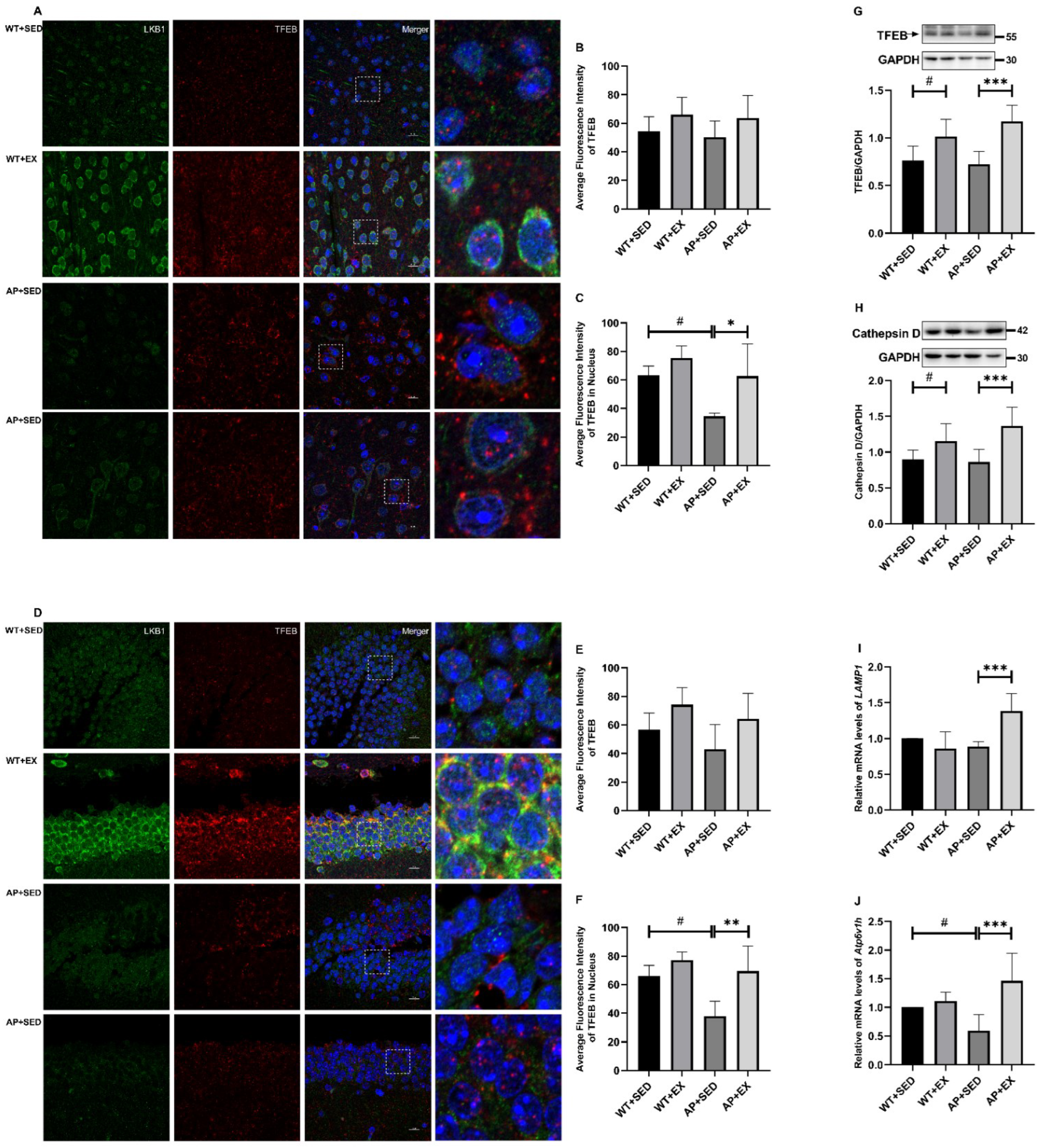
2.8. Exercise Activates TFEB and Lysosome Function in the Brain Cells of AD Mice
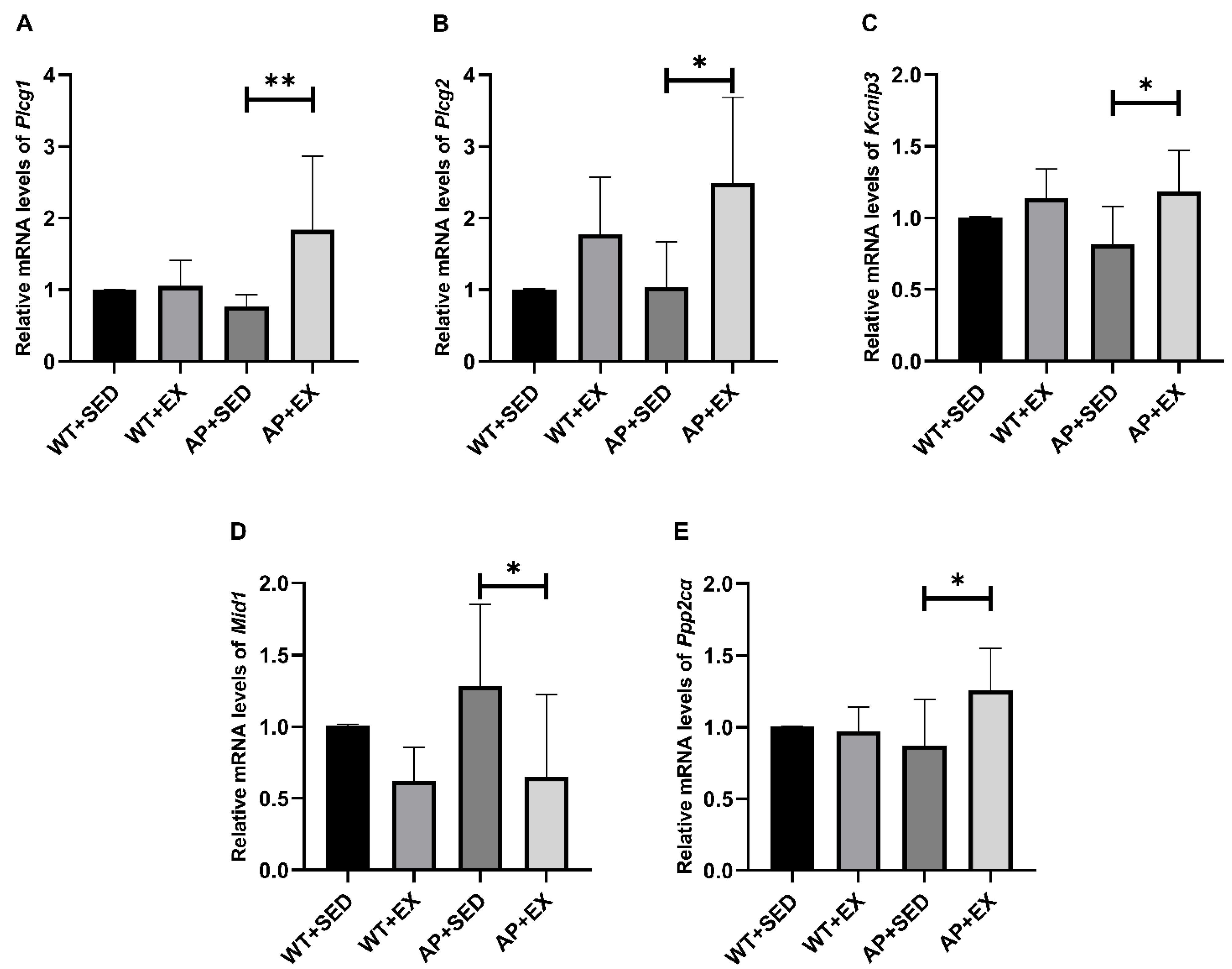
2.9. Exercise Activated PLC/Kcnip3/Mid1/PP2A Signaling Pathway in the Brain Cells of AD Mice
3. Discussion
4. Materials and Methods
4.1. Experimental Animals and Grouping
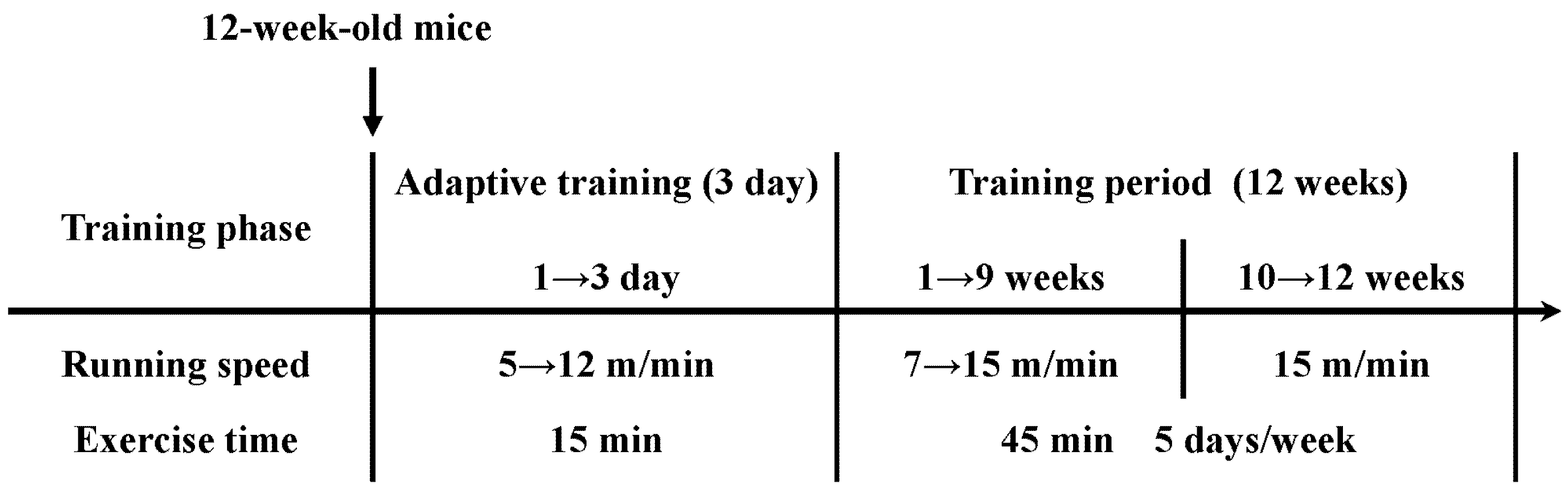
4.2. Exercise Protocols
4.3. Morris Water Maze Test
4.4. Immunohistochemical Detection
4.5. Enzyme-Linked Immunosorbent Assay
4.6. Real-Time PCR
4.7. Western Blotting
4.8. Golgi Staining
4.9. Immunofluorescence Staining
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hampel, H.; Hardy, J.; Blennow, K.; Chen, C.; Perry, G.; Kim, S.H.; Villemagne, V.L.; Aisen, P.; Vendruscolo, M.; Iwatsubo, T.; et al. The Amyloid-β Pathway in Alzheimer’s Disease. Mol. Psychiatry 2021, 26, 5481–5503. [Google Scholar] [CrossRef] [PubMed]
- Cai, Q.; Ganesan, D. Regulation of neuronal autophagy and the implications in neurodegenerative diseases. Neurobiol. Dis. 2022, 162, 105582. [Google Scholar] [CrossRef]
- Kuang, H.; Tan, C.Y.; Tian, H.Z.; Liu, L.H.; Yang, M.W.; Hong, F.F.; Yang, S.L. Exploring the bi-directional relationship between autophagy and Alzheimer’s disease. CNS Neurosci. Ther. 2020, 26, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Zhao, N.; Zhang, X.L.; Xia, J.; Yan, Q.W.; Xu, B. Effects of 12-week aerobic treadmill exercise on autophagy activity in the hippocampus of APP/PS1 mice. China Sport Sci. 2019, 39, 43–53. (In Chinese) [Google Scholar]
- Shen, K.; Liu, X.; Chen, D.; Chang, J.; Zhang, Y.; Kou, X. Voluntary wheel-running exercise attenuates brain aging of rats through activating miR-130a-mediated autophagy. Brain Res. Bull. 2021, 172, 203–211. [Google Scholar] [CrossRef]
- Wang, X.; Zhu, Y.T.; Zhu, Y.; Sun, Y.L.; Huang, J.; Li, Z.; Wang, Y.; Wu, J.C.; Qin, Z.H.; Lin, F. Long-term running exercise alleviates cognitive dysfunction in APP/PSEN1 transgenic mice via enhancing brain lysosomal function. Acta Pharmacol. Sin. 2022, 43, 580–861. [Google Scholar] [CrossRef]
- Kim, J.Y.; Barua, S.; Jeong, Y.J.; Lee, J.E. Adiponectin: The Potential Regulator and Therapeutic Target of Obesity and Alzheimer’s Disease. Int. J. Mol. Sci. 2020, 21, 6419. [Google Scholar] [CrossRef]
- Krause, M.P.; Milne, K.J.; Hawke, T.J. Adiponectin-Consideration for its Role in Skeletal Muscle Health. Int. J. Mol. Sci. 2019, 20, 1528. [Google Scholar] [CrossRef]
- Markofski, M.M.; Carrillo, A.E.; Timmerman, K.L.; Jennings, K.; Coen, P.M.; Pence, B.D.; Flynn, M.G. Exercise training modifies ghrelin and adiponectin concentrations and is related to inflammation in older adults. J. Gerontol. Ser. A Biomed. Sci. Med. Sci. 2014, 69, 675–681. [Google Scholar] [CrossRef]
- Huang, H.; Iida, K.T.; Sone, H.; Ajisaka, R. The regulation of adiponectin receptors expression by acute exercise in mice. Exp. Clin. Endocrinol. Diabetes 2007, 115, 417–422. [Google Scholar] [CrossRef] [PubMed]
- You, J.; Sun, L.; Wang, J.; Sun, F.; Wang, W.; Wang, D.; Fan, X.; Liu, D.; Xu, Z.; Qiu, C.; et al. Role of Adiponectin-Notch pathway in cognitive dysfunction associated with depression and in the therapeutic effect of physical exercise. Aging Cell 2021, 20, e13387. [Google Scholar] [CrossRef]
- Yau, S.Y.; Li, A.; Hoo, R.L.; Ching, Y.P.; Christie, B.R.; Lee, T.M.C.; Xu, A.; So, K.F. Physical exercise-induced hippocampal neurogenesis and antidepressant effects are mediated by the adipocyte hormone adiponectin. Proc. Natl. Acad. Sci. USA 2014, 111, 15810–15815. [Google Scholar] [CrossRef]
- Wang, P.; Liang, Y.; Chen, K.; Yau, S.Y.; Sun, X.; Cheng, K.K.Y.; Xu, A.; So, K.F.; Li, A. Potential Involvement of Adiponectin Signaling in Regulating Physical Exercise-Elicited Hippocampal Neurogenesis and Dendritic Morphology in Stressed Mice. Front. Cell Neurosci. 2020, 14, 189. [Google Scholar] [CrossRef]
- Yau, S.Y.; Lee, T.H.; Li, A.; Xu, A.; So, K.F. Adiponectin Mediates Running-Restored Hippocampal Neurogenesis in Streptozotocin-Induced Type 1 Diabetes in Mice. Front. Neurosci. 2018, 12, 679. [Google Scholar] [CrossRef]
- Yan, X.D.; Qu, X.S.; Yin, J.; Qiao, J.; Zhang, J.; Qi, J.S.; Wu, M.N. Adiponectin Ameliorates Cognitive Behaviors and in vivo Synaptic Plasticity Impairments in 3xTg-AD Mice. J. Alzheimer’s Dis. 2022, 85, 343–357. [Google Scholar] [CrossRef]
- He, K.; Nie, L.; Ali, T.; Wang, S.; Chen, X.; Liu, Z.; Li, W.; Zhang, K.; Xu, J.; Liu, J.; et al. Adiponectin alleviated Alzheimer-like pathologies via autophagy-lysosomal activation. Aging Cell 2021, 20, e13514. [Google Scholar] [CrossRef]
- Ali, T.; Rehman, S.U.; Khan, A.; Badshah, H.; Abid, N.B.; Kim, M.W.; Jo, M.H.; Chung, S.S.; Lee, H.G.; Rutten, B.P.F.; et al. Adiponectin-mimetic novel nonapeptide rescues aberrant neuronal metabolic-associated memory deficits in Alzheimer’s disease. Mol. Neurodegener. 2021, 16, 23. [Google Scholar] [CrossRef]
- Jian, Y.; Liu, W.F.; Yuan, S.L. Effects of adiponectin on exercise-induced autophagy in Alzheimer disease. Chin. J. Pathophysiol. 2022, 38, 154–161. (In Chinese) [Google Scholar]
- Rashtiani, S.; Goudarzi, I.; Jafari, A.; Rohampour, K. Adenosine monophosphate activated protein kinase (AMPK) is essential for the memory improving effect of adiponectin. Neurosci. Lett. 2021, 749, 135721. [Google Scholar] [CrossRef]
- Lopez-Ortiz, S.; Valenzuela, P.L.; Seisdedos, M.M.; Morales, J.S.; Vega, T.; Castillo-García, A.; Nisticò, R.; Mercuri, N.B.; Lista, S.; Lucia, A.; et al. Exercise interventions in Alzheimer’s disease: A systematic review and meta-analysis of randomized controlled trials. Ageing Res. Rev. 2021, 72, 101479. [Google Scholar] [CrossRef]
- Vorhees, C.V.; Williams, M.T. Value of water mazes for assessing spatial and egocentric learning and memory in rodent basic research and regulatory studies. Neurotoxicol. Teratol. 2014, 45, 75–90. [Google Scholar] [CrossRef] [PubMed]
- Walker, C.K.; Herskowitz, J.H. Dendritic Spines: Mediators of Cognitive Resilience in Aging and Alzheimer’s Disease. Neuroscientist 2021, 27, 487–505. [Google Scholar] [CrossRef]
- Zhang, Z.; Yang, X.; Song, Y.Q.; Tu, J. Autophagy in Alzheimer’s disease pathogenesis: Therapeutic potential and future perspectives. Ageing Res. Rev. 2021, 72, 101464. [Google Scholar] [CrossRef]
- Hwang, J.Y.; Yan, J.; Zukin, R.S. Autophagy and synaptic plasticity: Epigenetic regulation. Curr. Opin. Neurobiol. 2019, 59, 207–212. [Google Scholar] [CrossRef]
- Perluigi, M.; Di Domenico, F.; Barone, E.; Butterfield, D.A. mTOR in Alzheimer disease and its earlier stages: Links to oxidative damage in the progression of this dementing disorder. Free Radic. Biol. Med. 2021, 169, 382–396. [Google Scholar] [CrossRef]
- Pickford, F.; Masliah, E.; Britschgi, M.; Lucin, K.; Narasimhan, R.; Jaeger, P.A.; Small, S.; Spencer, B.; Rockenstein, E.; Levine, B.; et al. The autophagy-related protein beclin 1 shows reduced expression in early Alzheimer disease and regulates amyloid β accumulation in mice. J. Clin. Investig. 2008, 118, 2190–2199. [Google Scholar] [CrossRef]
- Klionsky, D.J.; Abdel-Aziz, A.K.; Abdelfatah, S.; Abdellatif, M.; Abdoli, A.; Abel, S.; Abeliovich, H.; Abildgaard, M.H.; Abudu, Y.P.; Acevedo-Arozena, A.; et al. Guidelines for the use and interpretation of assays for monitoring autophagy (4th edition). Autophagy 2021, 17, 1–382. [Google Scholar]
- Jiang, P.; Mizushima, N. LC3- and p62-based biochemical methods for the analysis of autophagy progression in mammalian cells. Methods 2015, 75, 13–18. [Google Scholar] [CrossRef]
- Fan, X.; Huang, T.; Tong, Y.; Fan, Z.; Yang, Z.; Yang, D.; Mao, X.; Yang, M. p62 works as a hub modulation in the ageing process. Ageing Res. Rev. 2022, 73, 101538. [Google Scholar] [CrossRef]
- Zhou, L.; Deepa, S.S.; Etzler, J.C.; Ryu, J.; Mao, X.; Fang, Q.; Liu, D.D.; Torres, J.M.; Jia, W.; Lechleiter, J.D.; et al. Adiponectin activates AMP-activated protein kinase in muscle cells via APPL1/LKB1-dependent and phospholipase C/Ca2+/Ca2+/calmodulin-dependent protein kinase kinase-dependent pathways. J. Biol. Chem. 2009, 284, 22426–22435. [Google Scholar] [CrossRef]
- Iwabu, M.; Yamauchi, T.; Okada-Iwabu, M.; Sato, K.; Nakagawa, T.; Funata, M.; Yamaguchi, M.; Namiki, S.; Nakayama, R.; Tabata, M.; et al. Adiponectin and AdipoR1 regulate PGC-1α and mitochondria by Ca2+ and AMPK/SIRT1. Nature 2010, 464, 1313–1319. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Zhang, Y.; Doycheva, D.M.; Ding, Y.; Zhang, Y.; Tang, J.; Guo, H.; Zhang, J.H. Adiponectin attenuates neuronal apoptosis induced by hypoxia-ischemia via the activation of AdipoR1/APPL1/LKB1/AMPK pathway in neonatal rats. Neuropharmacology 2018, 133, 415–428. [Google Scholar] [CrossRef] [PubMed]
- Iyaswamy, A.; Wang, X.; Krishnamoorthi, S.; Kaliamoorthy, V.; Sreenivasmurthy, S.G.; Durairajan, S.S.K.; Song, J.X.; Tong, B.C.K.; Zhu, Z.; Su, C.F.; et al. Theranostic F-SLOH mitigates Alzheimer’s disease pathology involving TFEB and ameliorates cognitive functions in Alzheimer’s disease models. Redox Biol. 2022, 51, 102280. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wang, K.; Long, A.; Jia, L.; Zhang, Y.; Deng, H.; Li, Y.; Han, J.; Wang, Y. Fasting-induced hormonal regulation of lysosomal function. Cell Res. 2017, 27, 748–763. [Google Scholar] [CrossRef]
- Wang, Y.T.; Chen, J.; Li, X.; Umetani, M.; Chen, Y.; Li, P.L.; Zhang, Y. Contribution of transcription factor EB to adipoRon-induced inhibition of arterial smooth muscle cell proliferation and migration. Am. J. Physiol. Cell Physiol. 2019, 317, C1034–C1047. [Google Scholar] [CrossRef]
- Sujkowski, A.; Hong, L.; Wessells, R.J.; Todi, S.V. The protective role of exercise against age-related neurodegeneration. Ageing Res. Rev. 2022, 74, 101543. [Google Scholar] [CrossRef]
- Yu, H.; Zhang, C.; Xia, J.; Xu, B. Treadmill Exercise Ameliorates Adult Hippocampal Neurogenesis Possibly by Adjusting the APP Proteolytic Pathway in APP/PS1 Transgenic Mice. Int. J. Mol. Sci. 2021, 22, 9570. [Google Scholar] [CrossRef]
- Zhang, L.; Tang, W.; Chao, F.L.; Zhou, C.N.; Jiang, L.; Zhang, Y.; Liang, X.; Tang, J.; Qi, Y.Q.; Yang, H.; et al. Four-month treadmill exercise prevents the decline in spatial learning and memory abilities and the loss of spinophilin-immunoreactive puncta in the hippocampus of APP/PS1 transgenic mice. Neurobiol. Dis. 2020, 136, 104723. [Google Scholar] [CrossRef]
- Krishnan, S.; Shrestha, Y.; Jayatunga, D.P.W.; Rea, S.; Martins, R.; Bharadwaj, P. Activate or Inhibit? Implications of Autophagy Modulation as a Therapeutic Strategy for Alzheimer’s Disease. Int. J. Mol. Sci. 2020, 21, 6739. [Google Scholar] [CrossRef]
- Lee, J.H.; Yang, D.S.; Goulbourne, C.N.; Im, E.; Stavrides, P.; Pensalfini, A.; Chan, H.; Bouchet-Marquis, C.; Cynthia, B.; Berg, M.J.; et al. Faulty autolysosome acidification in Alzheimer’s disease mouse models induces autophagic build-up of Aβ in neurons, yielding senile plaques. Nat. Neurosci. 2022, 25, 688–701. [Google Scholar] [CrossRef]
- Kandimalla, R.; Manczak, M.; Pradeepkiran, J.A.; Morton, H.; Reddy, P.H. A partial reduction of Drp1 improves cognitive behavior and enhances mitophagy, autophagy and dendritic spines in a transgenic Tau mouse model of Alzheimer disease. Hum. Mol. Genet. 2022, 31, 1788–1805. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Wang, X.; Lu, X.Y. Adiponectin Exerts Neurotrophic Effects on Dendritic Arborization, Spinogenesis, and Neurogenesis of the Dentate Gyrus of Male Mice. Endocrinology 2016, 157, 2853–2869. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Liu, B.; Yao, P.; Shao, Y.; Cheng, Y.; Zhao, J.; Wu, J.; Zhao, Z.W.; Huang, W.; Christopher, T.A.; et al. Adiponectin inhibits cardiac arrest/cardiopulmonary resuscitationinduced apoptosis in brain by increasing autophagy involved in AdipoR1AMPK signaling. Mol. Med. Rep. 2020, 22, 870–878. [Google Scholar] [CrossRef]
- Liu, W.; Xia, Y.; Kuang, H.; Wang, Z.; Liu, S.; Tang, C.; Yin, D. Proteomic Profile of Carbonylated Proteins Screen the Regulation of Calmodulin-Dependent Protein Kinases-AMPK-Beclin1 in Aerobic Exercise-Induced Autophagy in Middle-Aged Rat Hippocampus. Gerontology 2019, 65, 620–633. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Wang, Z.; Xia, Y.; Kuang, H.; Liu, S.; Li, L.; Tang, C.; Yin, D. The balance of apoptosis and autophagy via regulation of the AMPK signal pathway in aging rat striatum during regular aerobic exercise. Exp. Gerontol. 2019, 124, 110647. [Google Scholar] [CrossRef]
- Azimi, M.; Gharakhanlou, R.; Naghdi, N.; Khodadadi, D.; Heysieattalab, S. Moderate treadmill exercise ameliorates amyloid-β-induced learning and memory impairment, possibly via increasing AMPK activity and up-regulation of the PGC-1α/FNDC5/BDNF pathway. Peptides 2018, 102, 78–88. [Google Scholar] [CrossRef]
- Li, Z.; Ma, J.; Kuang, Z.; Jiang, Y. β-Asarone Attenuates Aβ-Induced Neuronal Damage in PC12 Cells Overexpressing APPswe by Restoring Autophagic Flux. Front. Pharmacol. 2021, 12, 701635. [Google Scholar] [CrossRef]
- Gao, L.J.; Dai, Y.; Li, X.Q.; Meng, S.; Zhong, Z.Q.; Xu, S.J. Chlorogenic acid enhances autophagy by upregulating lysosomal function to protect against SH-SY5Y cell injury induced by H2O2. Exp. Ther. Med. 2021, 21, 426. [Google Scholar] [CrossRef]
- Liu, Y.; Xue, X.; Zhang, H.; Che, X.; Luo, J.; Wang, P.; Xu, J.; Xing, Z.; Yuan, L.; Liu, Y.; et al. Neuronal-targeted TFEB rescues dysfunction of the autophagy-lysosomal pathway and alleviates ischemic injury in permanent cerebral ischemia. Autophagy 2019, 15, 493–509. [Google Scholar] [CrossRef]
- Huang, J.; Wang, X.; Zhu, Y.; Li, Z.; Zhu, Y.T.; Wu, J.C.; Qin, Z.H.; Xiang, M.; Lin, F. Exercise activates lysosomal function in the brain through AMPK-SIRT1-TFEB pathway. CNS Neurosci. Ther. 2019, 25, 796–807. [Google Scholar] [CrossRef]
- Steyn, F.J.; Boehme, F.; Vargas, E.; Wang, K.; Parkington, H.C.; Rao, J.R.; Chen, C. Adiponectin regulate growth hormone secretion via adiponectin receptor mediated Ca2+ signalling in rat somatotrophs in vitro. J. Neuroendocrinol. 2009, 21, 698–704. [Google Scholar] [CrossRef] [PubMed]
- Schweiger, S.; Matthes, F.; Posey, K.; Kickstein, E.; Weber, S.; Hettich, M.M.; Pfurtscheller, S.; Ehninger, D.; Schneider, R.; Krauß, S. Resveratrol induces dephosphorylation of Tau by interfering with the MID1-PP2A complex. Sci. Rep. 2017, 7, 13753. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Wang, Z.; Zhang, H.; Mahaman, Y.A.R.; Huang, F.; Meng, D.; Zhou, Y.; Wang, S.; Jiang, N.; Xiong, J.; et al. Chk1 Inhibition Ameliorates Alzheimer’s Disease Pathogenesis and Cognitive Dysfunction Through CIP2A/PP2A Signaling. Neurotherapeutics 2022, 19, 570–591. [Google Scholar] [CrossRef] [PubMed]
- Martina, J.A.; Puertollano, R. Protein phosphatase 2A stimulates activation of TFEB and TFE3 transcription factors in response to oxidative stress. J. Biol. Chem. 2018, 293, 12525–12534. [Google Scholar] [CrossRef]
- Baker, E.J.; Gleeson, T.T. The effects of intensity on the energetics of brief locomotor activity. J. Exp. Biol. 1999, 202, 3081–3087. [Google Scholar] [CrossRef] [PubMed]
- Ang, M.J.; Lee, S.; Wada, M.; Weerasinghe-Mudiyanselage, P.D.E.; Kim, S.H.; Shin, T.; Jeon, T.I.; Im, S.S.; Moon, C. SREBP-1c Deficiency Affects Hippocampal Micromorphometry and Hippocampus-Dependent Memory Ability in Mice. Int. J. Mol. Sci. 2021, 22, 6103. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| AdipoR1 | AATGGGGCTCCTTCTGGTAAC | GCAGACCTTATACACGAACTCC |
| APPL1 | AGCCAGTGACCCTTTATATCTGC | AGGTATCCAGCCTTTCGGGTT |
| LKB1 | CTGGACTCCGAGACCTTATGC | CAAGCTGGATCACATTCCGAT |
| mTOR | CTTCTTCCGTTCTATCTC | AAGTGTCAATCTGTATGG |
| Beclin1 | GAACTCTGGAGGTCTCGCT | CACCCAGGCTCGTTCTACC |
| P62 | AGTCCAGAATTCCTGCCTGA | TTCATTCAACTTCACATGAA |
| LAMP1 | CTGCTCCTGCTGCTGCT | AATTGTGAGGCTGGG |
| Atp6v1h | CCAAGATGGACATTCGAGGTG | CACTTTGTTGGCACGAACTTC |
| Plcg1 | GAGACGCGCCAGATCACAT | AAAGTCCCGAGAAGTCTTCCC |
| Plcg2 | AAATCCGTCCGGGGAAGAAC | TCCTCCTTTGAGTCCGTTGC |
| Kcnip3 | AGTGAACTGGAGTTATCCACGG | GTGAACTTGGTCTGAGCTTGT |
| Mid1 | CTGTGACGGCACCTGTCTC | AAACGGCTGACTGTTGGTCTT |
| Ppp2cα | ATGGACGAGAAGTTGTTCACC | CAGTGACTGGACATCGAACCT |
| GAPDH | CATGGCCTTCCGTGTTCCTA | CCTGCTTCACCACCTTCTTGAT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jian, Y.; Yuan, S.; Yang, J.; Lei, Y.; Li, X.; Liu, W. Aerobic Exercise Alleviates Abnormal Autophagy in Brain Cells of APP/PS1 Mice by Upregulating AdipoR1 Levels. Int. J. Mol. Sci. 2022, 23, 9921. https://doi.org/10.3390/ijms23179921
Jian Y, Yuan S, Yang J, Lei Y, Li X, Liu W. Aerobic Exercise Alleviates Abnormal Autophagy in Brain Cells of APP/PS1 Mice by Upregulating AdipoR1 Levels. International Journal of Molecular Sciences. 2022; 23(17):9921. https://doi.org/10.3390/ijms23179921
Chicago/Turabian StyleJian, Ye, Shunling Yuan, Jialun Yang, Yong Lei, Xuan Li, and Wenfeng Liu. 2022. "Aerobic Exercise Alleviates Abnormal Autophagy in Brain Cells of APP/PS1 Mice by Upregulating AdipoR1 Levels" International Journal of Molecular Sciences 23, no. 17: 9921. https://doi.org/10.3390/ijms23179921
APA StyleJian, Y., Yuan, S., Yang, J., Lei, Y., Li, X., & Liu, W. (2022). Aerobic Exercise Alleviates Abnormal Autophagy in Brain Cells of APP/PS1 Mice by Upregulating AdipoR1 Levels. International Journal of Molecular Sciences, 23(17), 9921. https://doi.org/10.3390/ijms23179921