

Dioscorea nipponica Makino Rhizome Extract and Its Active Compound Dioscin Protect against Neuroinflammation and Scopolamine-Induced Memory Deficits

 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
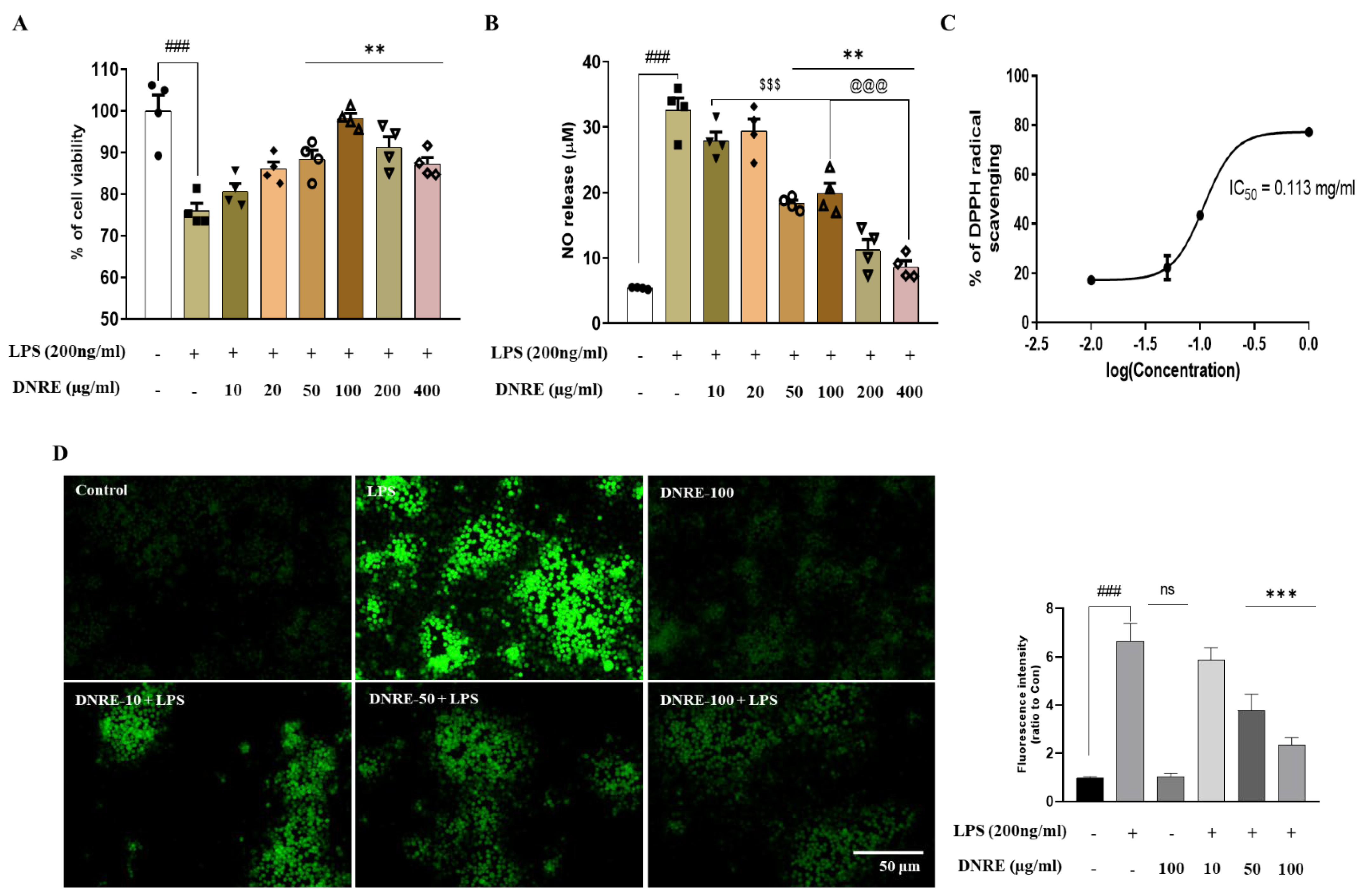
2.1. DNRE Protects Microglial Cells from LPS-Induced Stress
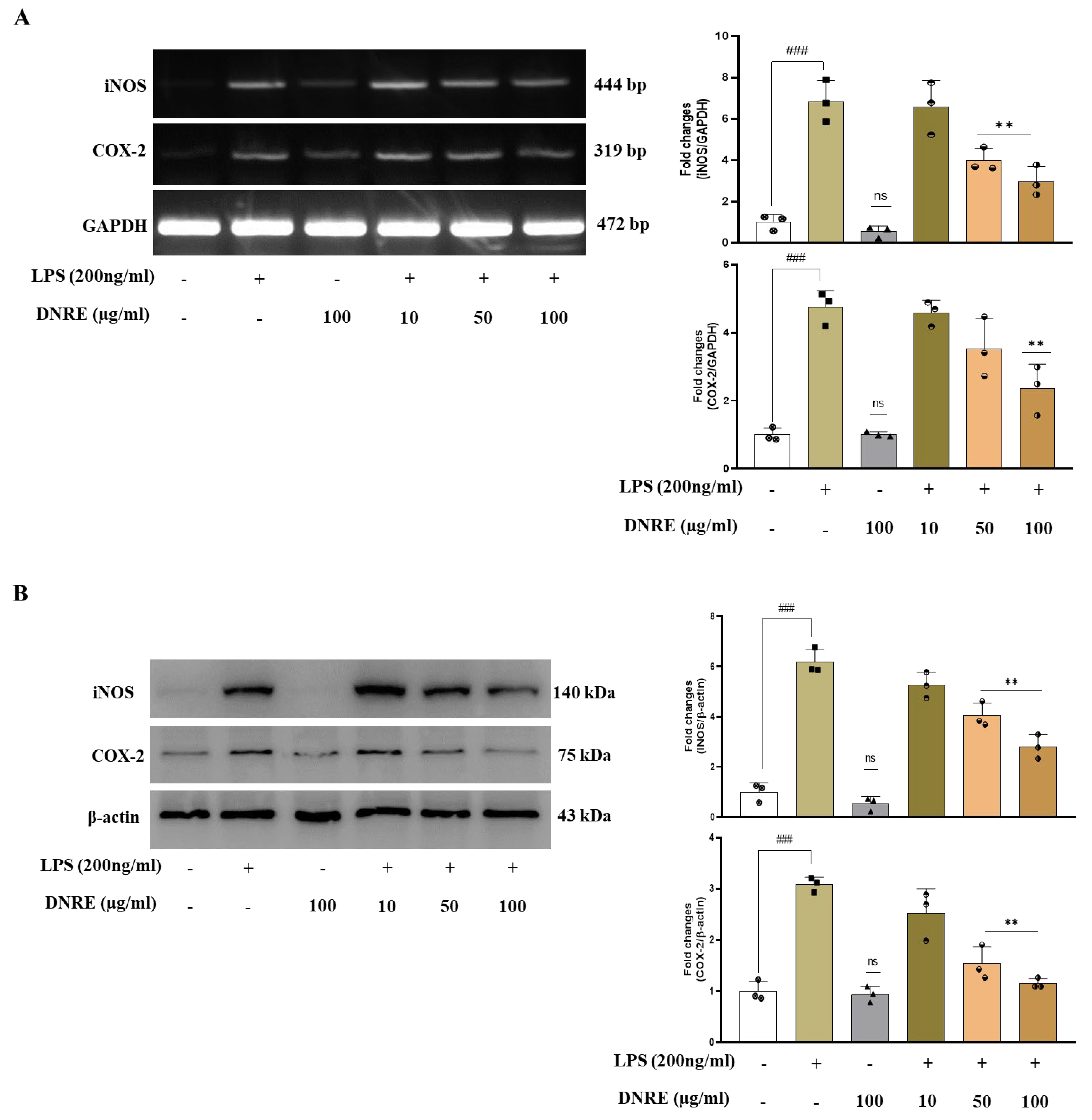
2.2. DNRE Pre-Treatment Suppresses Inflammatory Mediators following LPS Toxication
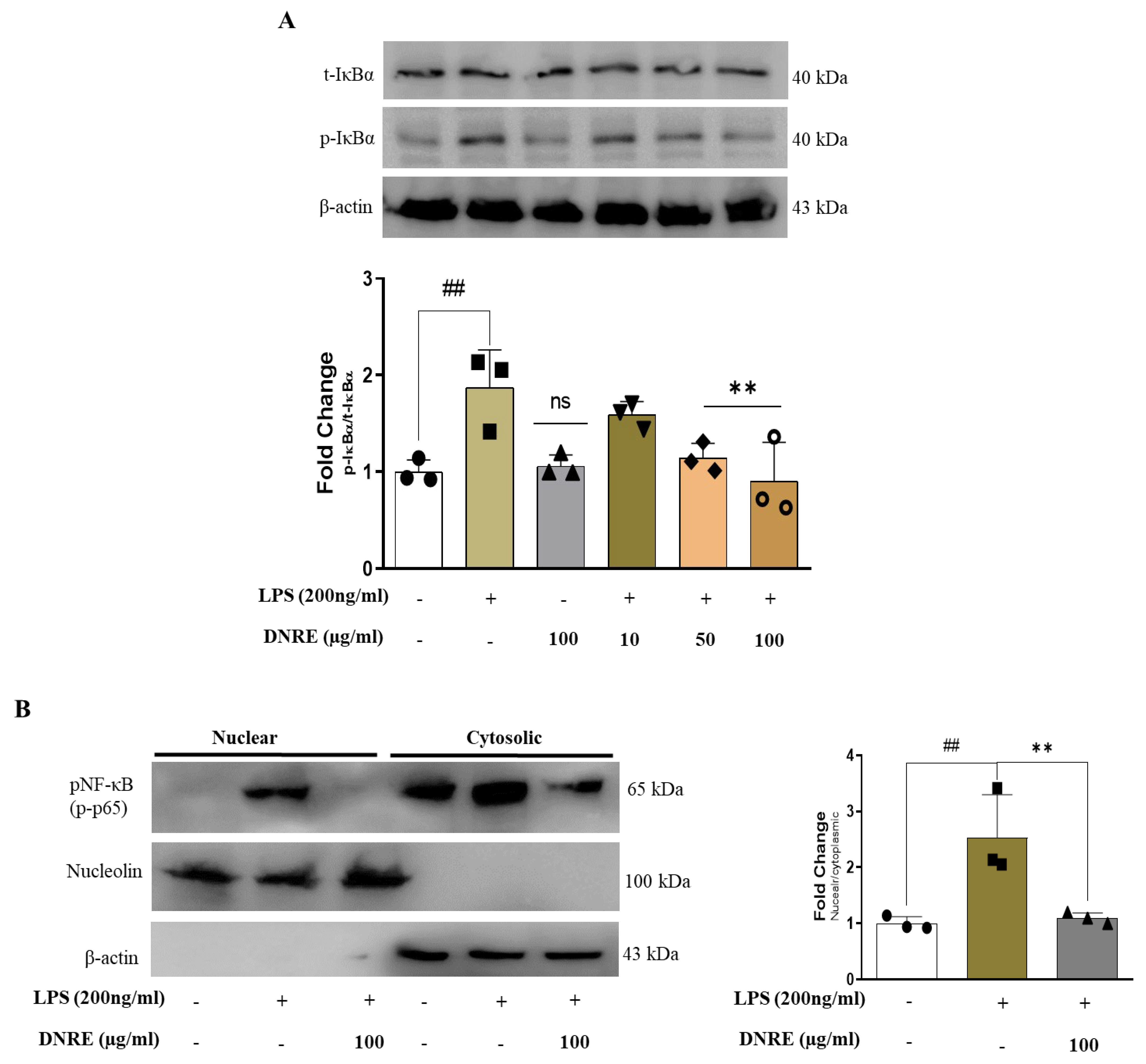
2.3. DNRE Inhibits Nuclear Translocation of NF-κB
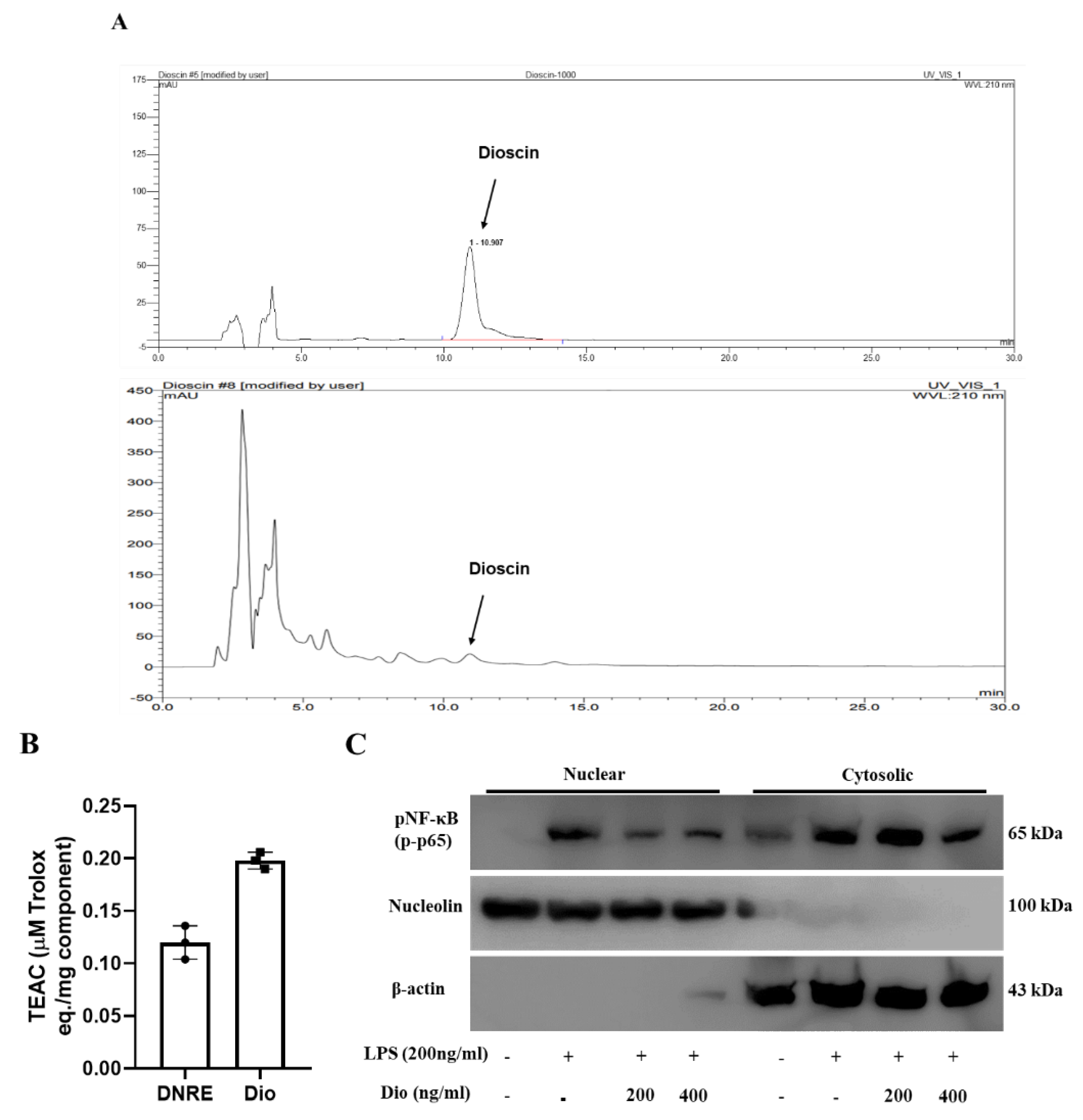
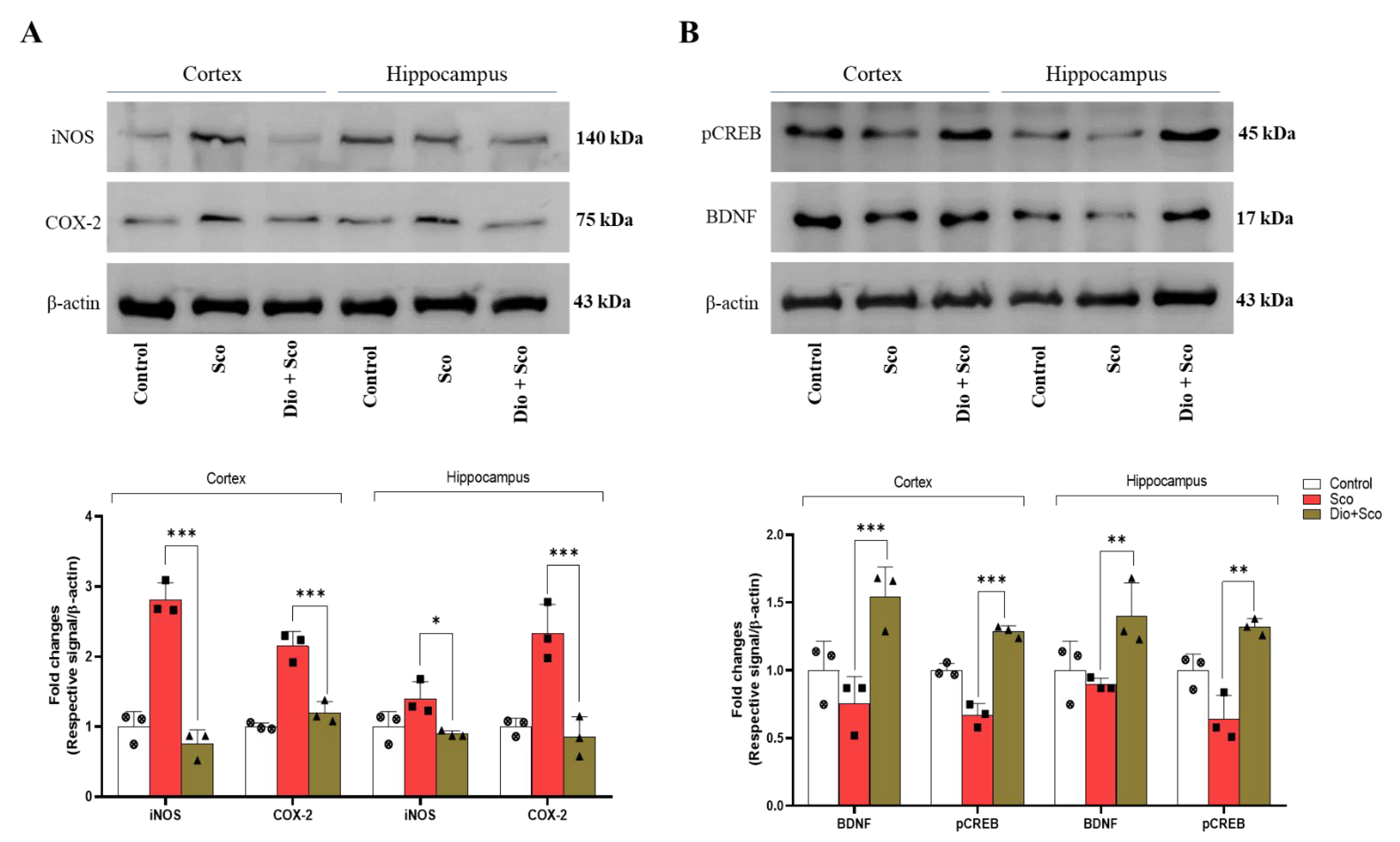
2.4. Anti-Neuroinflammatory Activities of Dioscin
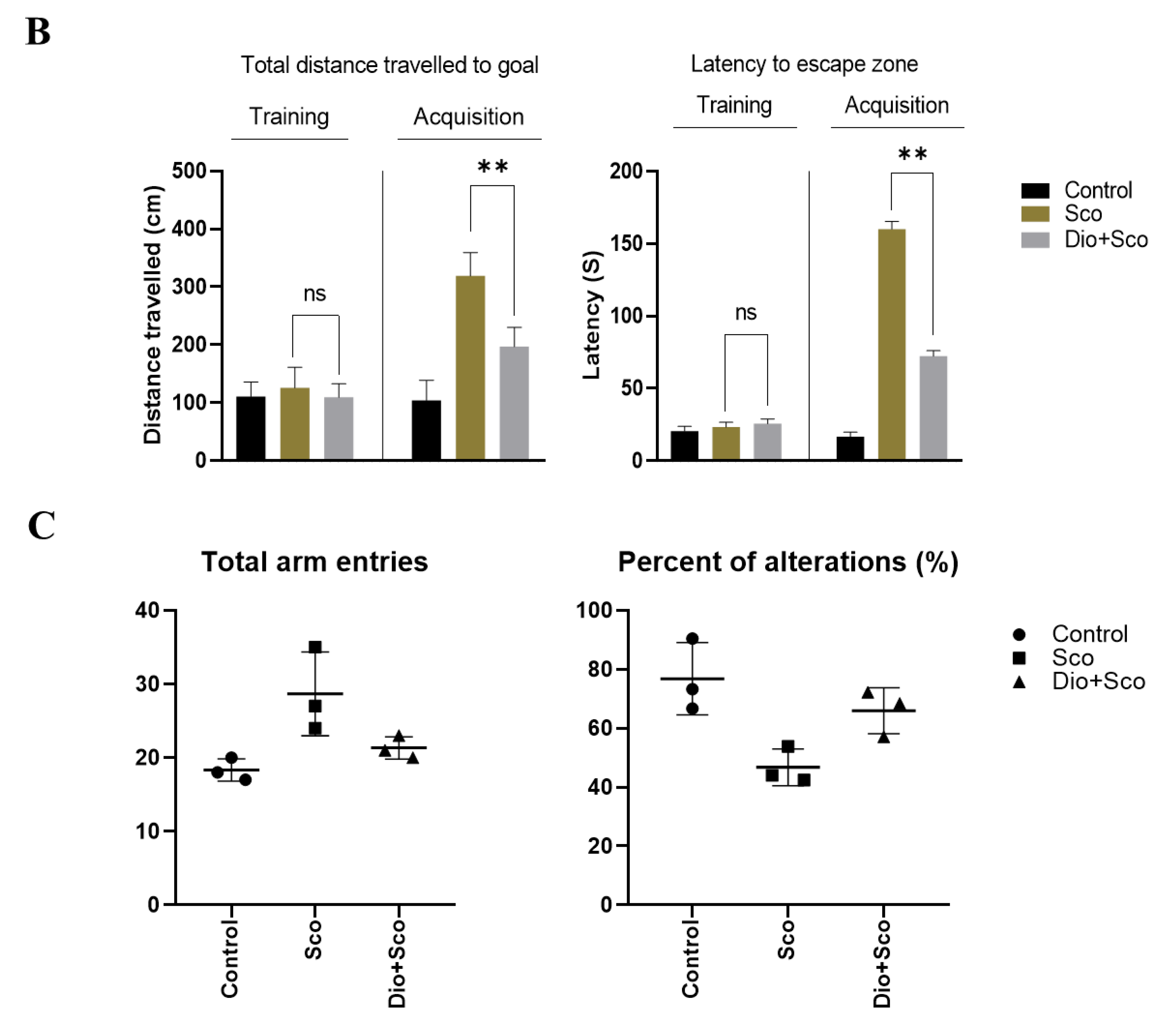
2.5. Dioscin Ameliorates Scopolamine-Induced Learning Deficits
2.6. Dioscin Ameliorates Scopolamine-Induced Neurotrophic Factor Deficits
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Cell Culture and Treatment
4.3. Cell Viability and Nitric Oxide Assay
4.4. DPPH Free Radical Scavenging Assay
4.5. DCFDA Assay
4.6. ABTS Radical Scavenging Activity
4.7. Nucleic–Cytosolic Fraction Preparation
4.8. Immunofluorescence
4.9. Determination of a Possible Active Compound in the Extract
4.10. Animals Handling and Treatment
4.11. Y-Maze Test
4.12. Morris Water Maze Test
4.13. Reverse Transcription–Polymerase Chain Reaction (RT–PCR)
4.14. Western Blot Analysis
4.15. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Glass, C.K.; Saijo, K.; Winner, B.; Marchetto, M.C.; Gage, F.H. Mechanisms underlying inflammation in neurodegeneration. Cell 2010, 140, 918–934. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Zhou, R.; Tong, Y.; Chen, P.; Shen, Y.; Miao, S.; Liu, X. Neuroprotection by dihydrotestosterone in lps-induced neuroinflammation. Neurobiol. Dis. 2020, 140, 104814. [Google Scholar] [CrossRef] [PubMed]
- Lehnardt, S.; Massillon, L.; Follett, P.; Jensen, F.E.; Ratan, R.; Rosenberg, P.A.; Volpe, J.J.; Vartanian, T. Activation of innate immunity in the cns triggers neurodegeneration through a toll-like receptor 4-dependent pathway. Proc. Natl. Acad. Sci. USA 2003, 100, 8514–8519. [Google Scholar] [CrossRef]
- Azam, S.; Jakaria, M.; Kim, I.S.; Kim, J.; Haque, M.E.; Choi, D.K. Regulation of toll-like receptor (tlr) signaling pathway by polyphenols in the treatment of age-linked neurodegenerative diseases: Focus on tlr4 signaling. Front. Immunol. 2019, 10, 1000. [Google Scholar] [CrossRef] [PubMed]
- He, P.; Yan, S.; Zheng, J.; Gao, Y.; Zhang, S.; Liu, Z.; Liu, X.; Xiao, C. Eriodictyol attenuates lps-induced neuroinflammation, amyloidogenesis, and cognitive impairments via the inhibition of nf-κb in male c57bl/6j mice and bv2 microglial cells. J. Agric. Food Chem. 2018, 66, 10205–10214. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.C.; Yeh, W.C.; Ohashi, P.S. Lps/tlr4 signal transduction pathway. Cytokine 2008, 42, 145–151. [Google Scholar] [CrossRef]
- Zhou, Y.L.; Yan, Y.M.; Li, S.Y.; He, D.H.; Xiong, S.; Wei, S.F.; Liu, W.; Hu, L.; Wang, Q.; Pan, H.F.; et al. 6-o-angeloylplenolin exerts neuroprotection against lipopolysaccharide-induced neuroinflammation in vitro and in vivo. Acta Pharmacol. Sin. 2020, 41, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Kilimann, I.; Hausner, L.; Fellgiebel, A.; Filippi, M.; Würdemann, T.J.; Heinsen, H.; Teipel, S.J. Parallel atrophy of cortex and basal forebrain cholinergic system in mild cognitive impairment. Cereb. Cortex 2017, 27, 1841–1848. [Google Scholar] [CrossRef] [PubMed]
- Hoover, D.B. Cholinergic modulation of the immune system presents new approaches for treating inflammation. Pharmacol. Ther. 2017, 179, 1–16. [Google Scholar] [CrossRef]
- Rahimzadegan, M.; Soodi, M. Comparison of memory impairment and oxidative stress following single or repeated doses administration of scopolamine in rat hippocampus. Basic Clin. Neurosci. 2018, 9, 5–14. [Google Scholar] [CrossRef] [Green Version]
- Wong-Guerra, M.; Jiménez-Martin, J.; Pardo-Andreu, G.L.; Fonseca-Fonseca, L.A.; Souza, D.O.; de Assis, A.M.; Ramirez-Sanchez, J.; Del Valle, R.M.; Nuñez-Figueredo, Y. Mitochondrial involvement in memory impairment induced by scopolamine in rats. Neurol. Res. 2017, 39, 649–659. [Google Scholar] [CrossRef]
- Cheon, S.Y.; Koo, B.N.; Kim, S.Y.; Kam, E.H.; Nam, J.; Kim, E.J. Scopolamine promotes neuroinflammation and delirium-like neuropsychiatric disorder in mice. Sci. Rep. 2021, 11, 8376. [Google Scholar] [CrossRef] [PubMed]
- Ou-Yang, S.H.; Jiang, T.; Zhu, L.; Yi, T. Dioscorea nipponica makino: A systematic review on its ethnobotany, phytochemical and pharmacological profiles. Chem. Cent. J. 2018, 12, 57. [Google Scholar] [CrossRef]
- Feng, J.F.; Tang, Y.N.; Ji, H.; Xiao, Z.G.; Zhu, L.; Yi, T. Biotransformation of dioscorea nipponica by rat intestinal microflora and cardioprotective effects of diosgenin. Oxidative Med. Cell. Longev. 2017, 2017, 4176518. [Google Scholar] [CrossRef] [PubMed]
- Woo, K.W.; Kwon, O.W.; Kim, S.Y.; Choi, S.Z.; Son, M.W.; Kim, K.H.; Lee, K.R. Phenolic derivatives from the rhizomes of dioscorea nipponica and their anti-neuroinflammatory and neuroprotective activities. J. Ethnopharmacol. 2014, 155, 1164–1170. [Google Scholar] [CrossRef] [PubMed]
- Park, K.-S.; Kim, H.J.; Hwang, J.T.; Ko, B.S. Dioscorea nipponica extracts enhance recovery from skeletal muscle atrophy by suppressing nf-κb expression. J. Funct. Foods 2020, 73, 104109. [Google Scholar] [CrossRef]
- Zhou, Q.; Sun, H.J.; Liu, S.M.; Jiang, X.H.; Wang, Q.Y.; Zhang, S.; Yu, D.H. Anti-inflammation effects of the total saponin fraction from dioscorea nipponica makino on rats with gouty arthritis by influencing mapk signalling pathway. BMC Complement. Med. Ther. 2020, 20, 261. [Google Scholar] [CrossRef]
- Huffman, W.J.; Subramaniyan, S.; Rodriguiz, R.M.; Wetsel, W.C.; Grill, W.M.; Terrando, N. Modulation of neuroinflammation and memory dysfunction using percutaneous vagus nerve stimulation in mice. Brain Stimul. 2019, 12, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; Wu, X.; Block, M.L.; Liu, Y.; Breese, G.R.; Hong, J.S.; Knapp, D.J.; Crews, F.T. Systemic lps causes chronic neuroinflammation and progressive neurodegeneration. Glia 2007, 55, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, A.R.; Yshii, L.M.; Viel, T.A.; Buck, H.S.; Mattson, M.P.; Scavone, C.; Kawamoto, E.M. Intermittent fasting attenuates lipopolysaccharide-induced neuroinflammation and memory impairment. J. Neuroinflamm. 2014, 11, 85. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.; Wang, D.; Yang, D.; Yao, J.; Tong, Y.; Chen, J. Characterization of steroidal saponins in crude extract from dioscorea nipponica makino by liquid chromatography tandem multi-stage mass spectrometry. Anal. Chim. Acta 2007, 599, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Azam, S.; Haque, M.E.; Cho, D.Y.; Kim, J.S.; Jakaria, M.; Kim, I.S.; Choi, D.K. Dioscin-mediated autophagy alleviates mpp+-induced neuronal degeneration: An in vitro parkinson’s disease model. Molecules 2022, 27, 2827. [Google Scholar] [CrossRef] [PubMed]
- Giridharan, S.; Srinivasan, M. Mechanisms of nf-κb p65 and strategies for therapeutic manipulation. J. Inflamm. Res. 2018, 11, 407–419. [Google Scholar] [CrossRef] [PubMed]
- Orihuela, R.; McPherson, C.A.; Harry, G.J. Microglial m1/m2 polarization and metabolic states. Br. J. Pharmacol. 2016, 173, 649–665. [Google Scholar] [CrossRef]
- Wang, W.Y.; Tan, M.S.; Yu, J.T.; Tan, L. Role of pro-inflammatory cytokines released from microglia in alzheimer’s disease. Ann. Transl. Med. 2015, 3, 136. [Google Scholar] [PubMed]
- Ricciotti, E.; FitzGerald, G.A. Prostaglandins and inflammation. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 986–1000. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Martin, M.; Michalek, S.M.; Katz, J. Role of mitogen-activated protein kinases and nf-kappab in the regulation of proinflammatory and anti-inflammatory cytokines by porphyromonas gingivalis hemagglutinin b. Infect. Immun. 2005, 73, 3990–3998. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. Nf-κb signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [PubMed]
- Park, K.J.; Suh, W.S.; Cha, J.M.; Park, J.E.; Woo, K.W.; Lee, K.R.; Kim, S.Y. Steroidal saponins from dioscorea nipponica rhizomes and their biological activity. Korean J. Pharmacogn. 2017, 48, 261–267. [Google Scholar]
- Yao, H.; Hu, C.; Yin, L.; Tao, X.; Xu, L.; Qi, Y.; Han, X.; Xu, Y.; Zhao, Y.; Wang, C.; et al. Dioscin reduces lipopolysaccharide-induced inflammatory liver injury via regulating tlr4/myd88 signal pathway. Int. Immunopharmacol. 2016, 36, 132–141. [Google Scholar] [CrossRef]
- Yao, H.; Sun, Y.; Song, S.; Qi, Y.; Tao, X.; Xu, L.; Yin, L.; Han, X.; Xu, Y.; Li, H.; et al. Protective effects of dioscin against lipopolysaccharide-induced acute lung injury through inhibition of oxidative stress and inflammation. Front. Pharmacol. 2017, 8, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vorhees, C.V.; Williams, M.T. Morris water maze: Procedures for assessing spatial and related forms of learning and memory. Nat. Protoc. 2006, 1, 848–858. [Google Scholar] [CrossRef]
- Castrén, E.; Antila, H. Neuronal plasticity and neurotrophic factors in drug responses. Mol. Psychiatry 2017, 22, 1085–1095. [Google Scholar] [CrossRef] [PubMed]
- Karthivashan, G.; Park, S.-Y.; Kweon, M.-H.; Kim, J.; Haque, M.E.; Cho, D.-Y.; Kim, I.-S.; Cho, E.-A.; Ganesan, P.; Choi, D.-K. Ameliorative potential of desalted salicornia europaea l. Extract in multifaceted alzheimer’s-like scopolamine-induced amnesic mice model. Sci. Rep. 2018, 8, 7174. [Google Scholar] [CrossRef] [PubMed]
- Jakaria, M.; Azam, S.; Cho, D.Y.; Haque, M.E.; Kim, I.S.; Choi, D.K. The methanol extract of allium cepa l. Protects inflammatory markers in lps-induced bv-2 microglial cells and upregulates the antiapoptotic gene and antioxidant enzymes in n27-a cells. Antioxidants 2019, 8, 348. [Google Scholar] [CrossRef]
- Tachakittirungrod, S.; Okonogi, S.; Chowwanapoonpohn, S. Study on antioxidant activity of certain plants in thailand: Mechanism of antioxidant action of guava leaf extract. Food Chem. 2007, 103, 381–388. [Google Scholar] [CrossRef]
- Runwal, G.; Stamatakou, E.; Siddiqi, F.H.; Puri, C.; Zhu, Y.; Rubinsztein, D.C. Lc3-positive structures are prominent in autophagy-deficient cells. Sci. Rep. 2019, 9, 10147. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.Z.; Zhou, H.Y.; Yan, Q. Fingerprint analysis of dioscorea nipponica by high-performance liquid chromatography with evaporative light scattering detection. Anal. Chim. Acta 2007, 582, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Zheng, L.; Yin, L.; Xu, L.; Qi, Y.; Han, X.; Xu, Y.; Liu, K.; Peng, J. Protective effects of the total saponins from dioscorea nipponica makino against carbon tetrachloride-induced liver injury in mice through suppression of apoptosis and inflammation. Int. Immunopharmacol. 2014, 19, 233–244. [Google Scholar] [CrossRef] [PubMed]
- Jo, S.H.; Kang, T.B.; Koppula, S.; Cho, D.Y.; Kim, J.S.; Kim, I.S.; Choi, D.K. Mitigating effect of lindera obtusiloba blume extract on neuroinflammation in microglial cells and scopolamine-induced amnesia in mice. Molecules 2021, 26, 2870. [Google Scholar] [CrossRef] [PubMed]
- Kraeuter, A.K.; Guest, P.C.; Sarnyai, Z. The y-maze for assessment of spatial working and reference memory in mice. Methods Mol. Biol. 2019, 1916, 105–111. [Google Scholar] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence | Size (bp) |
|---|---|---|
| iNOS | F 5′-GAG GTA CTC AGC GTC CTC CA-3′ R 5′-AGG GAG GAA AGG GAG AGA GG-3′ | 444 |
| COX-2 | F 5′-TGA GTG GTA GCC AGC AAA GC-3′ R 5′-CTG CAG TCC AGG TTC AAT GG-3′ | 319 |
| TNF-α | F 5′-TTC GAG TGA CAA GCC TGT AGC-3′ R 5′-AGA TTG ACC TCA GCG CTG AGT-3′ | 390 |
| IL-1β | F 5′-CAA GGA GAA CCA AGC AAC GA-3′ R 5′-TTG GCC GAG GAC TAA GGA GT-3′ | 428 |
| IL-6 | F 5′-GGA GGC TTA ATT ACA CAT GTT-3′ R 5′-TGA TTT CAA GAT GAA TTG GAT-3′ | 435 |
| GAPDH | F 5′- ACC ACA GTC CAT GCC ATC AC-3′ R 5′- CCA CCA CCC TGT TGC TGT AG-3′ | 472 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azam, S.; Kim, Y.-S.; Jakaria, M.; Yu, Y.-J.; Ahn, J.-Y.; Kim, I.-S.; Choi, D.-K. Dioscorea nipponica Makino Rhizome Extract and Its Active Compound Dioscin Protect against Neuroinflammation and Scopolamine-Induced Memory Deficits. Int. J. Mol. Sci. 2022, 23, 9923. https://doi.org/10.3390/ijms23179923
Azam S, Kim Y-S, Jakaria M, Yu Y-J, Ahn J-Y, Kim I-S, Choi D-K. Dioscorea nipponica Makino Rhizome Extract and Its Active Compound Dioscin Protect against Neuroinflammation and Scopolamine-Induced Memory Deficits. International Journal of Molecular Sciences. 2022; 23(17):9923. https://doi.org/10.3390/ijms23179923
Chicago/Turabian StyleAzam, Shofiul, Yon-Suk Kim, Md. Jakaria, Ye-Ji Yu, Jae-Yong Ahn, In-Su Kim, and Dong-Kug Choi. 2022. "Dioscorea nipponica Makino Rhizome Extract and Its Active Compound Dioscin Protect against Neuroinflammation and Scopolamine-Induced Memory Deficits" International Journal of Molecular Sciences 23, no. 17: 9923. https://doi.org/10.3390/ijms23179923
APA StyleAzam, S., Kim, Y. -S., Jakaria, M., Yu, Y. -J., Ahn, J. -Y., Kim, I. -S., & Choi, D. -K. (2022). Dioscorea nipponica Makino Rhizome Extract and Its Active Compound Dioscin Protect against Neuroinflammation and Scopolamine-Induced Memory Deficits. International Journal of Molecular Sciences, 23(17), 9923. https://doi.org/10.3390/ijms23179923