
Identification of the Key Functional Domains of Bombyx mori Nucleopolyhedrovirus IE1 Protein
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
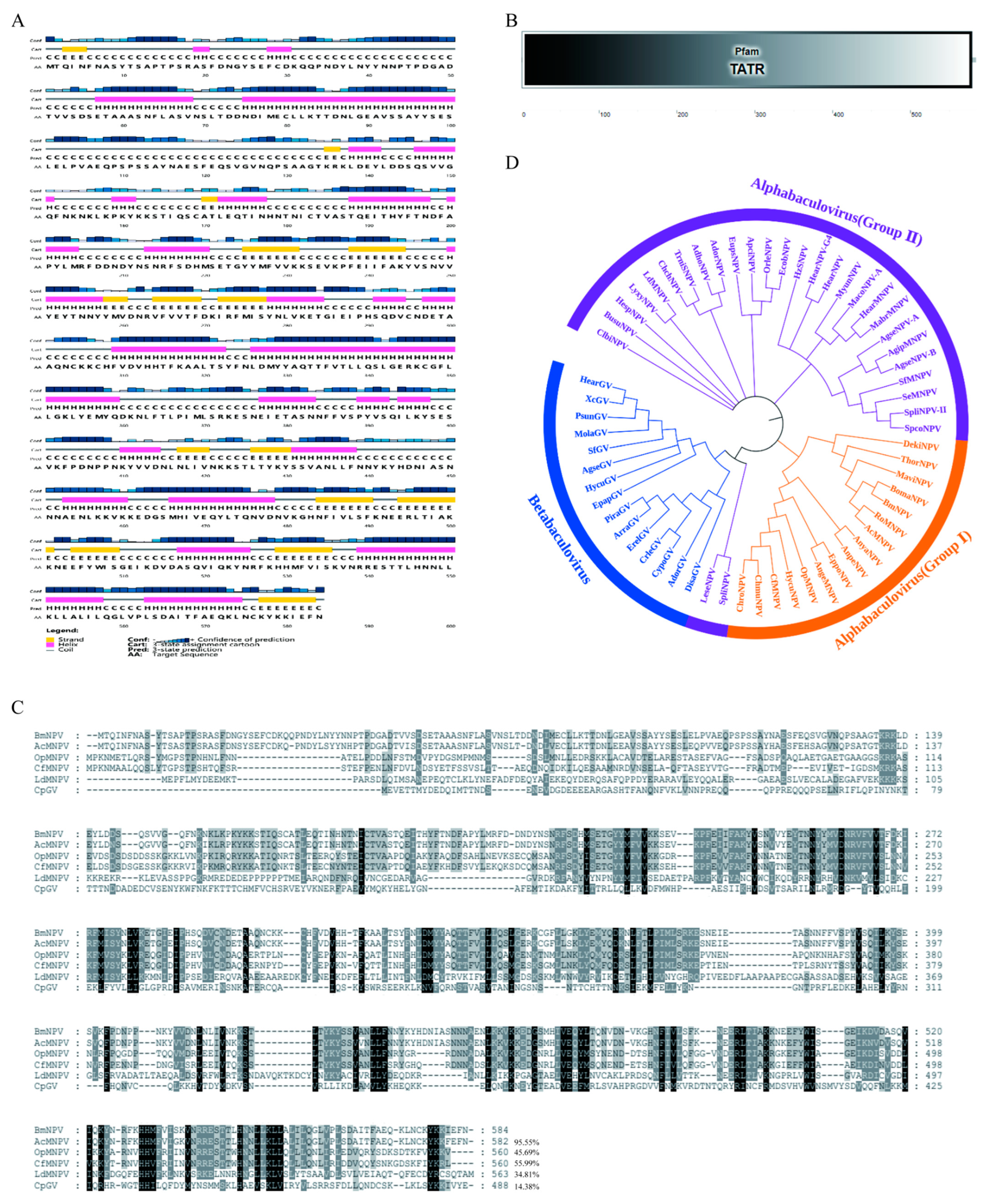
2.1. Sequence Characterization of BmNPV IE1
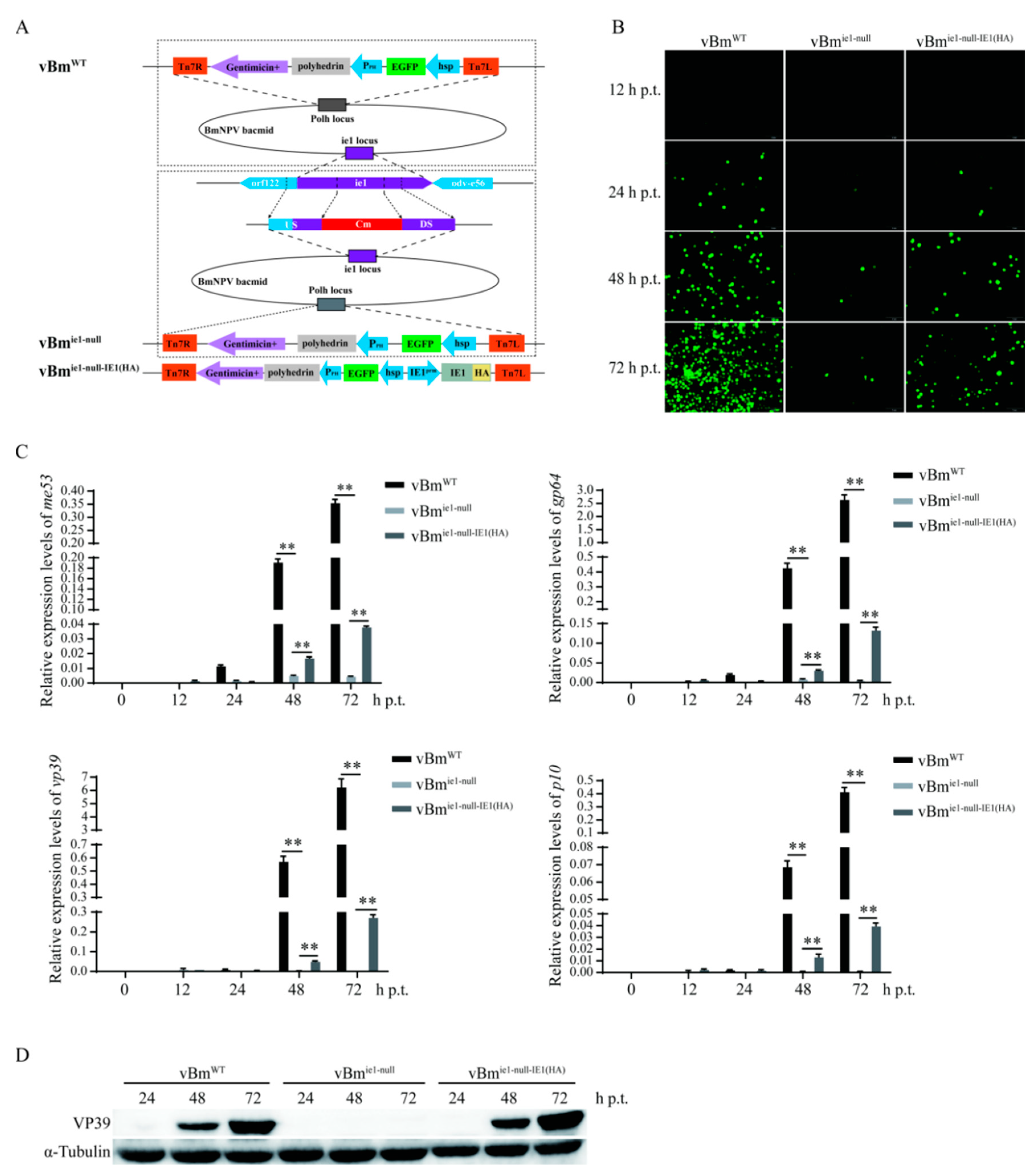
2.2. Effect of IE1 on the Proliferation of BmNPV
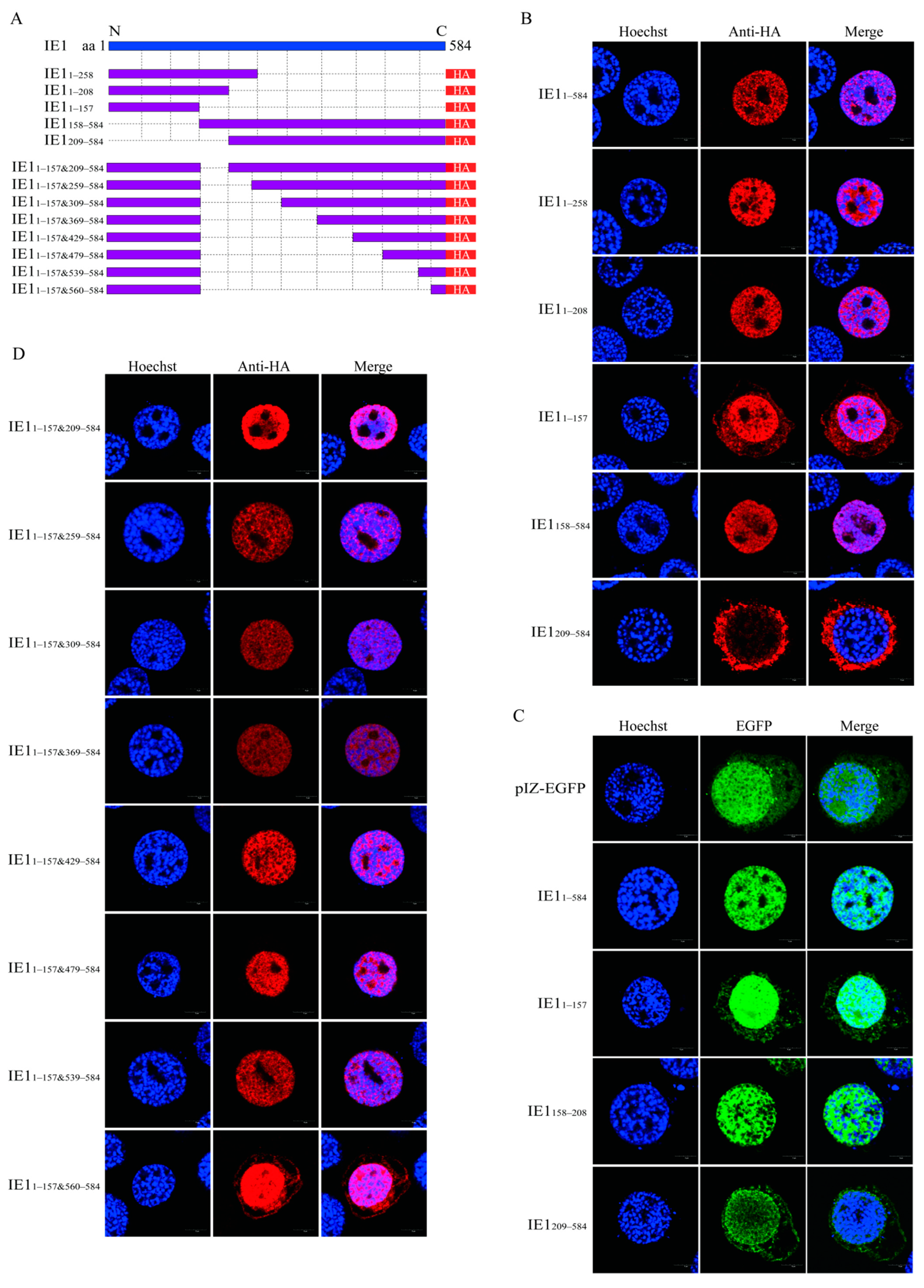
2.3. Identification of the Nuclear Localization Elements of BmNPV IE1
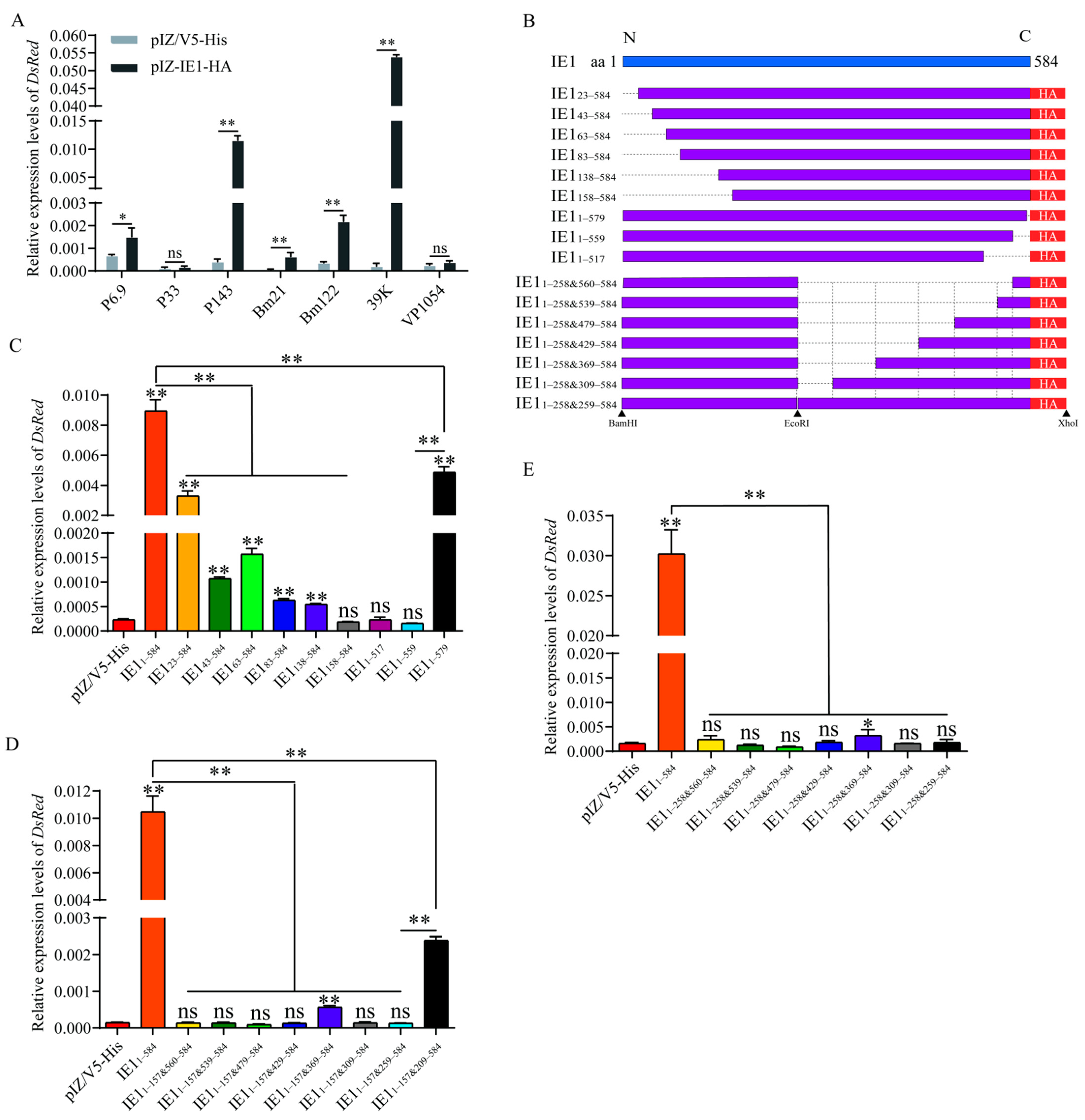
2.4. Identification of Key Transactivation Domains of BmNPV IE1
3. Discussion
4. Materials and Methods
4.1. Sequence Characterization of BmNPV IE1
4.2. Cells and Transient Transfection
4.3. Construction of ie-1 Deletion and ie-1 Repair Recombinant Viruses
4.4. Plasmid Construction
4.5. Immunofluorescence Assay
4.6. Western Blotting
4.7. Real Time Fluorescent Quantitative PCR (qRT-PCR)
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rohrmann, G.F. Introduction to the Baculoviruses, Their Taxonomy, and Evolution. In Baculovirus Molecular Biology, 4th ed.; Bethesda: Rockville, MD, USA, 2019; p. 2. [Google Scholar]
- Wang, M.; Hu, Z. Advances in Molecular Biology of Baculoviruses. Curr. Issues Mol. Biol. 2020, 34, 183–214. [Google Scholar] [CrossRef] [PubMed]
- Mansouri, M.; Berger, P. Baculovirus for gene delivery to mammalian cells: Past, present and future. Plasmid 2018, 98, 1–7. [Google Scholar] [PubMed]
- Varikkodan, M.M.; Chen, C.C.; Wu, T.Y. Recombinant Baculovirus: A Flexible Drug Screening Platform for Chikungunya Virus. Int. J. Mol. Sci. 2021, 22, 7891–7908. [Google Scholar]
- Hussain, A.G.; Wennmann, J.T.; Goergen, G.; Bryon, A.; Ros, V.I.D. Viruses of the Fall Armyworm Spodoptera frugiperda: A Review with Prospects for Biological Control. Viruses 2021, 13, 2220–2240. [Google Scholar]
- van der Merwe, M.; Jukes, M.D.; Rabalski, L.; Knox, C.; Opoku-Debrah, J.K.; Moore, S.D.; Krejmer-Rabalska, M.; Szewczyk, B.; Hill, M.P. Genome Analysis and Genetic Stability of the Cryptophlebia leucotreta Granulovirus (CrleGV-SA) after 15 Years of Commercial Use as a Biopesticide. Int. J. Mol. Sci. 2017, 18, 2327–2340. [Google Scholar]
- Lu, A.; Miller, L.K. The roles of eighteen baculovirus late expression factor genes in transcription and DNA replication. J. Virol. 1995, 69, 975–982. [Google Scholar] [PubMed]
- Kovacs, G.R.; Choi, J.; Guarino, L.A.; Summers, M.D. Functional dissection of the Autographa californica nuclear polyhedrosis virus immediate-early 1 transcriptional regulatory protein. J. Virol. 1992, 66, 7429–7437. [Google Scholar]
- Slack, J.M.; Blissard, G.W. Identification of two independent transcriptional activation domains in the Autographa californica multicapsid nuclear polyhedrosis virus IE1 protein. J. Virol. 1997, 71, 9579–9587. [Google Scholar]
- Olson, V.A.; Wetter, J.A.; Friesen, P.D. The highly conserved basic domain I of baculovirus IE1 is required for hr enhancer DNA binding and hr-dependent transactivation. J. Virol. 2003, 77, 5668–5677. [Google Scholar]
- Taggart, D.J.; Mitchell, J.K.; Friesen, P.D. A conserved N-terminal domain mediates required DNA replication activities and phosphorylation of the transcriptional activator IE1 of Autographa californica multicapsid nucleopolyhedrovirus. J. Virol. 2012, 86, 6575–6585. [Google Scholar] [PubMed]
- Olson, V.A.; Wetter, J.A.; Friesen, P.D. Oligomerization mediated by a helix-loop-helix-like domain of baculovirus IE1 is required for early promoter transactivation. J. Virol. 2001, 75, 6042–6051. [Google Scholar]
- Rodems, S.M.; Pullen, S.S.; Friesen, P.D. DNA-dependent transregulation by IE1 of Autographa californica nuclear polyhedrosis virus: IE1 domains required for transactivation and DNA binding. J. Virol. 1997, 71, 9270–9277. [Google Scholar] [PubMed]
- Olson, V.A.; Wetter, J.A.; Friesen, P.D. Baculovirus transregulator IE1 requires a dimeric nuclear localization element for nuclear import and promoter activation. J. Virol. 2002, 76, 9505–9515. [Google Scholar] [PubMed]
- Liao, N.; Dong, Z.; Zhang, X.; Qin, Q.; Luo, Y.; Huang, L.; Chen, P.; Lu, C.; Pan, M. Construction of a CRISPR/FnCas12a multi-sites editing system for inhibiting proliferation of Bombyx mori nuclearpolyhedrosisvirus. Int. J. Biol. Macromol. 2021, 193, 585–591. [Google Scholar] [PubMed]
- Yamada, Y.; Matsuyama, T.; Quan, G.X.; Kanda, T.; Tamura, T.; Sahara, K.; Asano, S.; Bando, H. Use of an N-terminal half truncated IE1 as an antagonist of IE1, an essential regulatory protein in baculovirus. Virus Res. 2002, 90, 253–261. [Google Scholar] [PubMed]
- Cao, M.Y.; Kuang, X.X.; Li, H.Q.; Lei, X.J.; Xiao, W.F.; Dong, Z.Q.; Zhang, J.; Hu, N.; Chen, T.T.; Lu, C.; et al. Screening and optimization of an efficient Bombyx mori nucleopolyhedrovirus inducible promoter. J. Biotechnol. 2016, 231, 72–80. [Google Scholar]
- Gomi, S.; Majima, K.; Maeda, S. Sequence analysis of the genome of Bombyx mori nucleopolyhedrovirus. J. Gen. Virol. 1999, 80, 1323–1337. [Google Scholar]
- Katou, Y.; Ikeda, M.; Kobayashi, M. Abortive replication of Bombyx mori nucleopolyhedrovirus in Sf9 and High Five cells: Defective nuclear transport of the virions. Virology 2006, 347, 455–465. [Google Scholar]
- Kaikkonen, M.U.; Maatta, A.I.; Ylä-Herttuala, S.; Airenne, K.J. Screening of complement inhibitors: Shielded baculoviruses increase the safety and efficacy of gene delivery. Mol. Ther. 2010, 18, 987–992. [Google Scholar]
- Murges, D.; Kremer, A.; Knebel-Mörsdorf, D. Baculovirus transactivator IE1 is functional in mammalian cells. J. Gen. Virol. 1997, 78, 1507–1510. [Google Scholar] [CrossRef]
- Efrose, R.; Swevers, L.; Iatrou, K. Baculoviruses deficient in ie1 gene function abrogate viral gene expression in transduced mammalian cells. Virology 2010, 406, 293–301. [Google Scholar] [PubMed] [Green Version]
- Liu, X.; Li, Y.; Hu, X.; Yi, Y.; Zhang, Z. Gene delivery and gene expression in vertebrate using baculovirus Bombyx mori nucleopolyhedrovirus vector. Oncotarget 2017, 8, 106017–106025. [Google Scholar] [PubMed]
- Dickison, V.L.; Willis, L.G.; Sokal, N.R.; Theilmann, D.A. Deletion of AcMNPV ac146 eliminates the production of budded virus. Virology 2012, 431, 29–39. [Google Scholar] [PubMed]
- He, L.; Shao, W.; Li, J.; Deng, F.; Wang, H.; Hu, Z.; Wang, M. Systematic analysis of nuclear localization of Autographa californica multiple nucleopolyhedrovirus proteins. J. Gen. Virol. 2021, 102, 1517–1526. [Google Scholar]
- Kaffman, A.; O’Shea, E.K. Regulation of nuclear localization: A key to a door. Annu. Rev. Cell Dev. Biol. 1999, 15, 291–339. [Google Scholar]
- Chevigné, A.; Fievez, V.; Schmit, J.C.; Deroo, S. Engineering and screening the N-terminus of chemokines for drug discovery. Biochem. Pharmacol. 2011, 82, 1438–1456. [Google Scholar]
- Hansen, L.S.; Sparre-Ulrich, A.H.; Christensen, M.; Knop, F.K.; Hartmann, B.; Holst, J.J.; Rosenkilde, M.M. N-terminally and C-terminally truncated forms of glucose-dependent insulinotropic polypeptide are high-affinity competitive antagonists of the human GIP receptor. Br. J. Pharmacol. 2016, 173, 826–838. [Google Scholar]
- Yang, B.; Gelfanov, V.M.; Perez-Tilve, D.; DuBois, B.; Rohlfs, R.; Levy, J.; Douros, J.D.; Finan, B.; Mayer, J.P.; DiMarchi, R.D. Optimization of Truncated Glucagon Peptides to Achieve Selective, High Potency, Full Antagonists. J. Med. Chem. 2021, 64, 4697–4708. [Google Scholar]
- Chang, C.L.; Hsu, S.Y.T. Development of chimeric and bifunctional antagonists for CLR/RAMP receptors. PLoS ONE 2019, 14, e0216996–e0217015. [Google Scholar]
- Kallunki, T.; Barisic, M.; Jäättelä, M.; Liu, B. How to Choose the Right Inducible Gene Expression System for Mammalian Studies? Cells 2019, 8, 796–811. [Google Scholar]
- Dong, Z.Q.; Chen, T.T.; Zhang, J.; Hu, N.; Cao, M.Y.; Dong, F.F.; Jiang, Y.M.; Chen, P.; Lu, C.; Pan, M.H. Establishment of a highly efficient virus-inducible CRISPR/Cas9 system in insect cells. Antivir. Res. 2016, 130, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Huang, L.; Dong, F.; Hu, Z.; Qin, Q.; Long, J.; Cao, M.; Chen, P.; Lu, C.; Pan, M.H. Establishment of a baculovirus-inducible CRISPR/Cas9 system for antiviral research in transgenic silkworms. Appl. Microbiol. Biotechnol. 2018, 102, 9255–9265. [Google Scholar] [CrossRef] [PubMed]
- Buchan, D.W.A.; Jones, D.T. The PSIPRED Protein Analysis Workbench: 20 years on. Nucleic Acids Res. 2019, 47, W402–W407. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Khedkar, S.; Bork, P. SMART: Recent updates, new developments and status in 2020. Nucleic Acids Res. 2021, 49, D458–D460. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.H.; Cai, X.J.; Liu, M.; Lv, J.; Tang, H.; Tan, J.; Lu, C. Establishment and characterization of an ovarian cell line of the silkworm, Bombyx mori. Tissue Cell 2010, 42, 42–46. [Google Scholar] [CrossRef]
- Dong, Z.Q.; Zhang, J.; Chen, X.M.; He, Q.; Cao, M.Y.; Wang, L.; Li, H.Q.; Xiao, W.F.; Pan, C.X.; Lu, C.; et al. Bombyx mori nucleopolyhedrovirus ORF79 is a per os infectivity factor associated with the PIF complex. Virus Res. 2014, 184, 62–70. [Google Scholar] [CrossRef]
- Dong, Z.Q.; Hu, N.; Zhang, J.; Chen, T.T.; Cao, M.Y.; Li, H.Q.; Lei, X.J.; Chen, P.; Lu, C.; Pan, M.H. Oligomerization of Baculovirus LEF-11 Is Involved in Viral DNA Replication. PLoS ONE 2015, 10, e0144930–e0144942. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, Z.-G.; Dong, Z.-Q.; Miao, J.-H.; Li, K.-J.; Wang, J.; Chen, P.; Lu, C.; Pan, M.-H. Identification of the Key Functional Domains of Bombyx mori Nucleopolyhedrovirus IE1 Protein. Int. J. Mol. Sci. 2022, 23, 10276. https://doi.org/10.3390/ijms231810276
Hu Z-G, Dong Z-Q, Miao J-H, Li K-J, Wang J, Chen P, Lu C, Pan M-H. Identification of the Key Functional Domains of Bombyx mori Nucleopolyhedrovirus IE1 Protein. International Journal of Molecular Sciences. 2022; 23(18):10276. https://doi.org/10.3390/ijms231810276
Chicago/Turabian StyleHu, Zhi-Gang, Zhan-Qi Dong, Jiang-Hao Miao, Ke-Jie Li, Jie Wang, Peng Chen, Cheng Lu, and Min-Hui Pan. 2022. "Identification of the Key Functional Domains of Bombyx mori Nucleopolyhedrovirus IE1 Protein" International Journal of Molecular Sciences 23, no. 18: 10276. https://doi.org/10.3390/ijms231810276
APA StyleHu, Z. -G., Dong, Z. -Q., Miao, J. -H., Li, K. -J., Wang, J., Chen, P., Lu, C., & Pan, M. -H. (2022). Identification of the Key Functional Domains of Bombyx mori Nucleopolyhedrovirus IE1 Protein. International Journal of Molecular Sciences, 23(18), 10276. https://doi.org/10.3390/ijms231810276