
HMG20A Inhibit Adipogenesis by Transcriptional and Epigenetic Regulation of MEF2C Expression
Abstract
:1. Introduction
2. Results
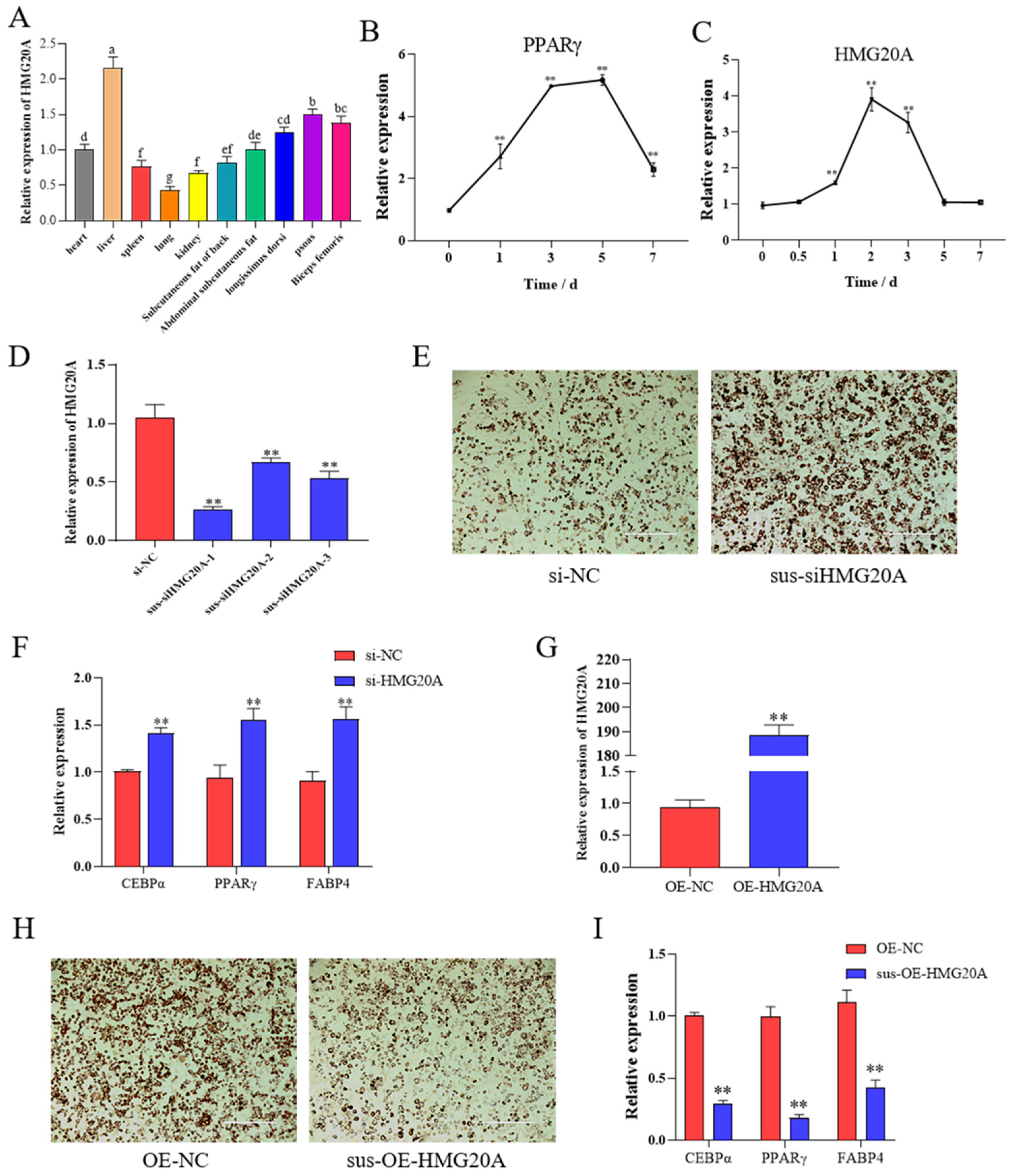
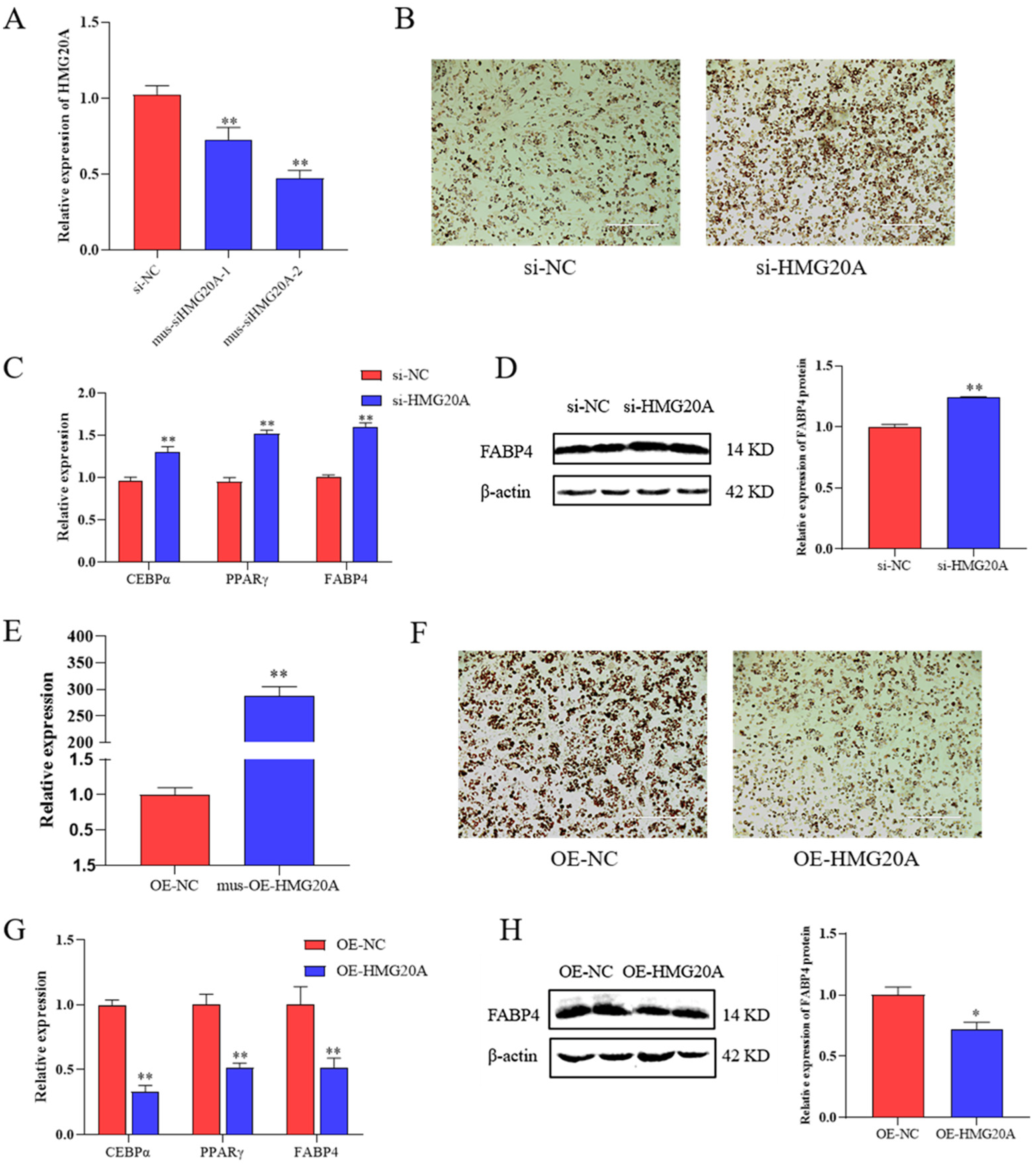
2.1. HMG20A Silencing Promotes Adipogenesis of SVF Cells
2.2. HMG20A Slows Adipogenesis in C3H10T1/2 Mesenchymal Cells
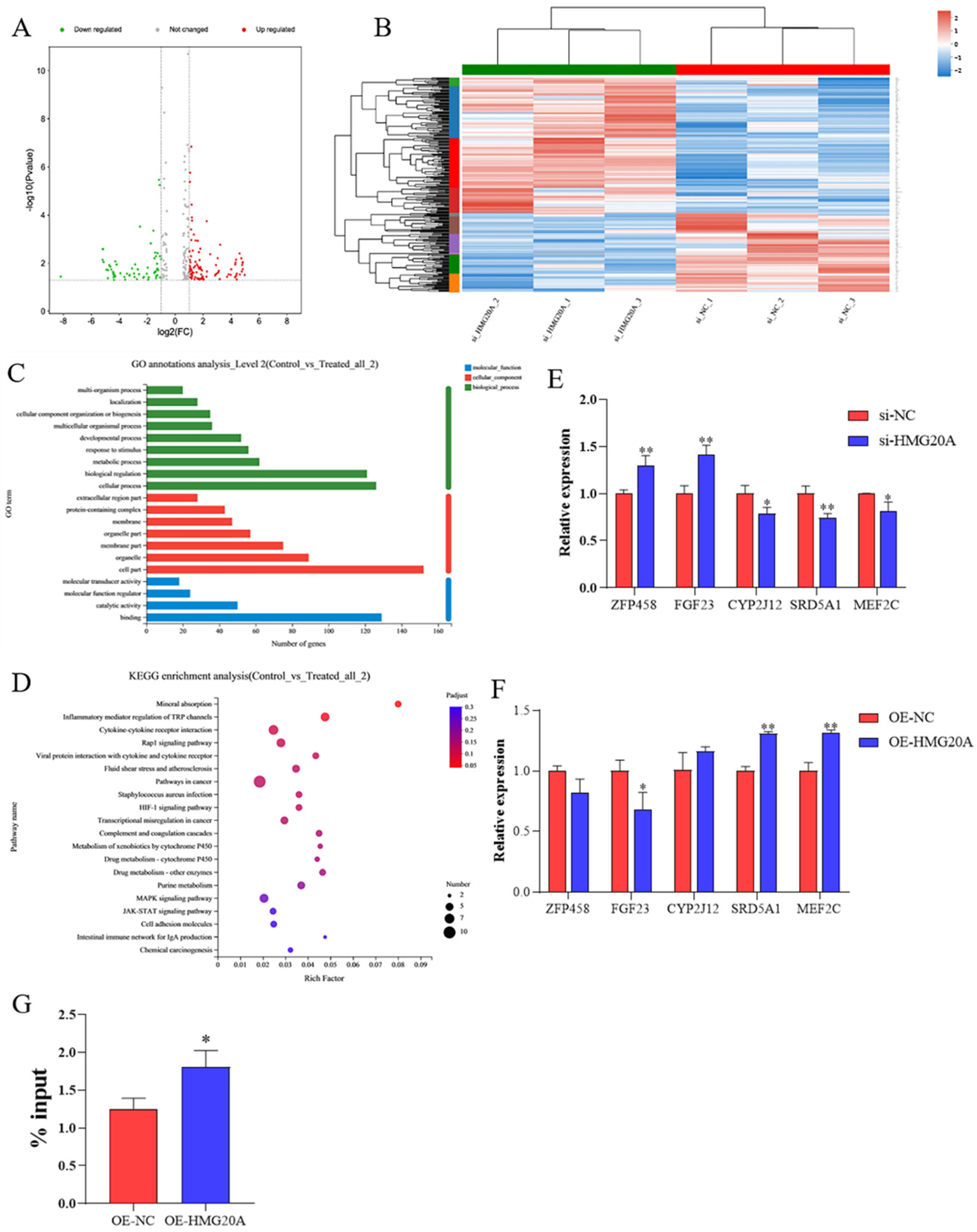
2.3. Transcriptomics Analysis Indicates MEF2C as a Target of HMG20A
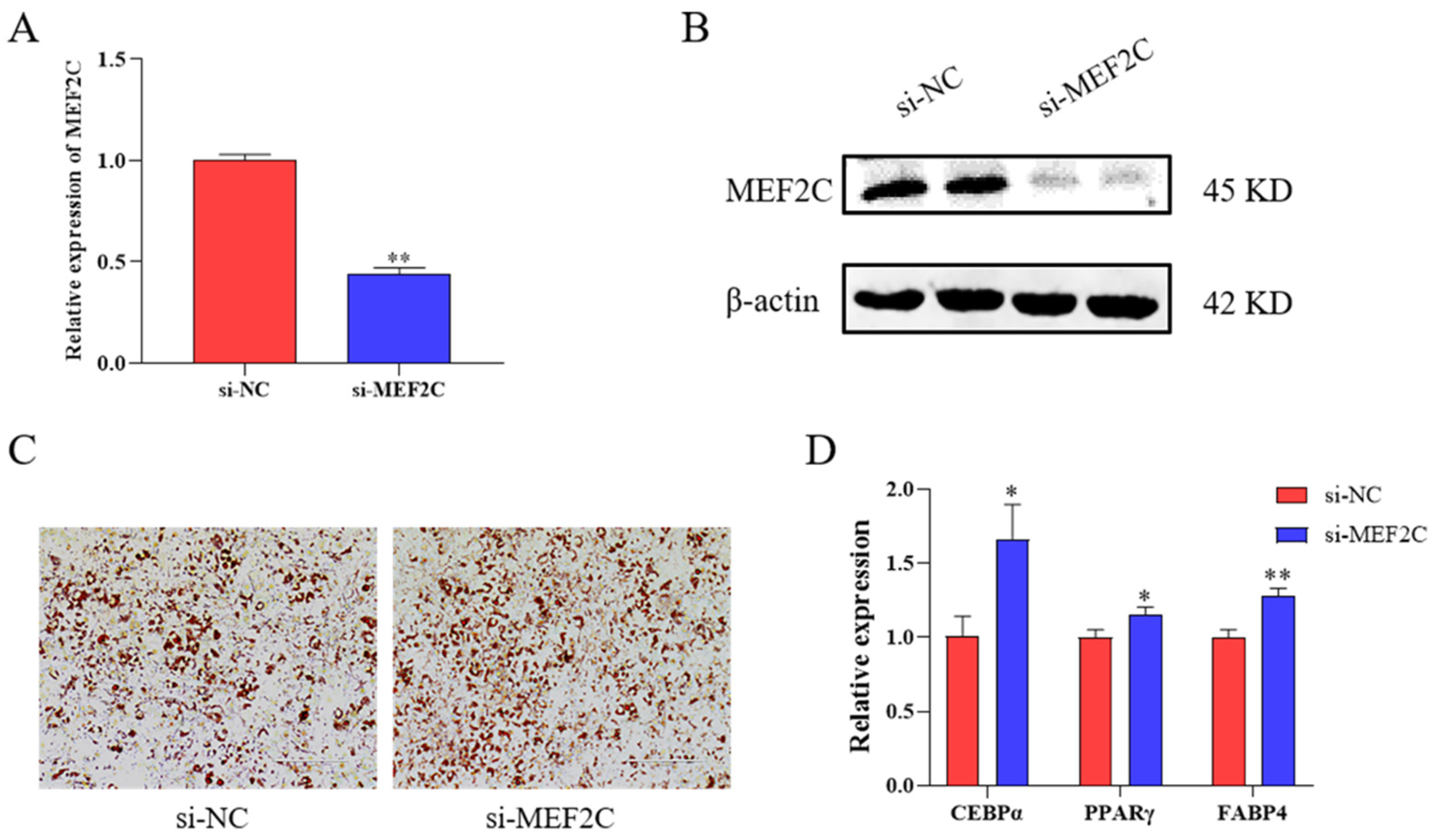
2.4. SiRNA Targeting MEF2C Promotes Adipogenesis in C3H10T1/2 Cells
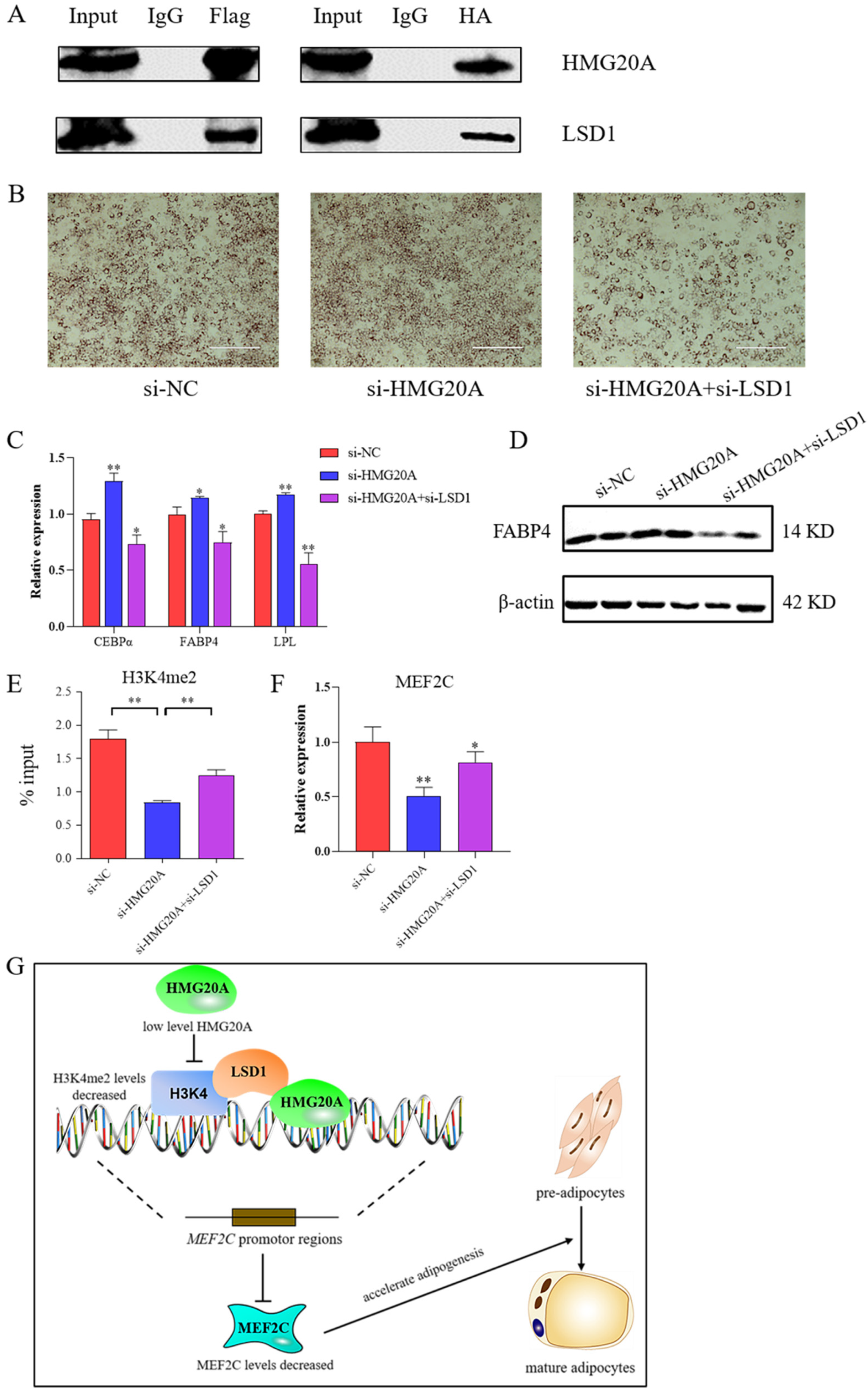
2.5. HMG20A and LSD1 Protein Interaction Exists in Adipogenesis
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Plasmid and siRNA Transfection
4.3. Adipogenic Differentiation and OIL Red O Staining
4.4. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.5. Western Blot and Co-Inmunoprecipitation (CoIP)
4.6. RNA Sequencing (RNA-Seq)
4.7. Chromatin Immunoprecipitation (ChIP) Assay
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- An, M.R.; Cg, D.; Jl, E.; Nan, C.; Cc, B. Trade openness and the obesity epidemic: A cross-national study of 175 countries during 1975–2016. Ann. Epidemiol. 2019, 37, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Flier, S. Obesity and the Regulation of Energy Balance. Cell 2001, 104, 531–543. [Google Scholar]
- Farooqi, S.I. Genetic, molecular and physiological mechanisms involved in human obesity: Society for Endocrinology Medal Lecture 2012. Clin. Endocrinol. 2015, 82, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Jaacks, L.M.; Vandevijvere, S.; Pan, A.; Mcgowan, C.J.; Wallace, C.; Imamura, F.; Mozaffarian, D.; Swinburn, B.; Ezzati, M. The obesity transition: Stages of the global epidemic. Lancet Diabetes Endocrinol. 2019, 7, 231–240. [Google Scholar] [CrossRef]
- Zhou, Y.; Peng, J.; Jiang, S. Role of histone acetyltransferases and histone deacetylases in adipocyte differentiation and adipogenesis. Eur. J. Cell Biol. 2014, 93, 170–177. [Google Scholar] [CrossRef]
- Moseti, D.; Regassa, A.; Kim, W.K. Molecular Regulation of Adipogenesis and Potential Anti-Adipogenic Bioactive Molecules. Int. J. Mol. Sci. 2016, 17, 124. [Google Scholar] [CrossRef]
- Ambele, M.; Dhanraj, P.; Giles, R.; Pepper, M. Adipogenesis: A Complex Interplay of Multiple Molecular Determinants and Pathways. Int. J. Mol. Sci. 2020, 21, 4283. [Google Scholar] [CrossRef]
- Lin, T.; Zhang, Y.; Zhang, T.; Stecker, R.A.; Yang, X. Hop2 interacts with the transcription factor CEBPα and suppresses adipocyte differentiation—ScienceDirect. J. Biol. Chem. 2021, 297, 101264. [Google Scholar] [CrossRef]
- Martínez, J.; Milagro, F.; Claycombe, K.; Schalinske, K. Epigenetics in adipose tissue, obesity, weight loss, and diabetes. Adv. Nutr. 2014, 5, 71–81. [Google Scholar] [CrossRef]
- Stricker, S.; Köferle, A.; Beck, S. From profiles to function in epigenomics. Nat. Rev. Genet. 2017, 18, 51–66. [Google Scholar] [CrossRef]
- Tessarz, P.; Kouzarides, T. Histone core modifications regulating nucleosome structure and dynamics. Nat. Rev. Mol. Cell Biol. 2014, 15, 703–708. [Google Scholar] [CrossRef] [PubMed]
- Hyun, K.; Jeon, J.; Park, K.; Kim, J. Writing, erasing and reading histone lysine methylations. Exp. Mol. Med. 2017, 49, e324. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Zhao, S. Physiology, affecting factors and strategies for control of pig meat intramuscular fat. Recent Pat. Food Nutr. Agric. 2009, 1, 59–74. [Google Scholar]
- Hocquette, J.; Gondret, F.; Baéza, E.; Médale, F.; Jurie, C.; Pethick, D. Intramuscular fat content in meat-producing animals: Development, genetic and nutritional control, and identification of putative markers. Anim. Int. J. Anim. Biosci. 2010, 4, 303–319. [Google Scholar] [CrossRef] [PubMed]
- Mateescu, R.; Garrick, D.; Garmyn, A.; VanOverbeke, D.; Mafi, G.; Reecy, J. Genetic parameters for sensory traits in longissimus muscle and their associations with tenderness, marbling score, and intramuscular fat in Angus cattle. J. Anim. Sci. 2015, 93, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Wei, H.; Song, T.; Yang, Y.; Peng, J.; Jiang, S. Transcriptome comparison between porcine subcutaneous and intramuscular stromal vascular cells during adipogenic differentiation. PLoS ONE 2013, 8, e77094. [Google Scholar]
- Malarkey, C.; Churchill, M. The high mobility group box: The ultimate utility player of a cell. Trends Biochem. Sci. 2012, 37, 553–562. [Google Scholar] [CrossRef]
- Hakimi, M.; Bochar, D.; Chenoweth, J.; Lane, W.; Mandel, G.; Shiekhattar, R. A core-BRAF35 complex containing histone deacetylase mediates repression of neuronal-specific genes. Proc. Natl. Acad. Sci. USA 2002, 99, 7420–7425. [Google Scholar] [CrossRef]
- Ceballos-Chávez, M.; Rivero, S.; García-Gutiérrez, P.; Rodríguez-Paredes, M.; García-Domínguez, M.; Bhattacharya, S.; Reyes, J. Control of neuronal differentiation by sumoylation of BRAF35, a subunit of the LSD1-CoREST histone demethylase complex. Proc. Natl. Acad. Sci. USA 2012, 109, 8085–8090. [Google Scholar] [CrossRef]
- Rivero, S.; Ceballos-Chávez, M.; Bhattacharya, S.; Reyes, J. HMG20A is required for SNAI1-mediated epithelial to mesenchymal transition. Oncogene 2015, 34, 5264–5276. [Google Scholar] [CrossRef]
- Kooner, J.; Saleheen, D.; Sim, X.; Sehmi, J.; Zhang, W.; Frossard, P.; Been, L.; Chia, K.; Dimas, A.; Hassanali, N.; et al. Genome-wide association study in individuals of South Asian ancestry identifies six new type 2 diabetes susceptibility loci. Nat. Genet. 2011, 43, 984–989. [Google Scholar] [CrossRef] [PubMed]
- Mellado-Gil, J.; Fuente-Martín, E.; Lorenzo, P.; Cobo-Vuilleumier, N.; López-Noriega, L.; Martín-Montalvo, A.; Gómez, I.; Ceballos-Chávez, M.; Gómez-Jaramillo, L.; Campos-Caro, A.; et al. The type 2 diabetes-associated HMG20A gene is mandatory for islet beta cell functional maturity. Cell Death Dis. 2018, 9, 279. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Reichel, J.; Han, C.; Zuniga-Hertz, J.; Cai, D. Astrocytic Process Plasticity and IKKβ/NF-κB in Central Control of Blood Glucose, Blood Pressure, and Body Weight. Cell Metab. 2017, 25, 1091–1102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuente-Martín, E.; Mellado-Gil, J.; Cobo-Vuilleumier, N.; Martín-Montalvo, A.; Romero-Zerbo, S.; Diaz Contreras, I.; Hmadcha, A.; Soria, B.; Martin Bermudo, F.; Reyes, J.; et al. Dissecting the Brain/Islet Axis in Metabesity. Genes 2019, 10, 350. [Google Scholar] [CrossRef] [PubMed]
- Wynder, C.; Hakimi, M.; Epstein, J.; Shilatifard, A.; Shiekhattar, R. Recruitment of MLL by HMG-domain protein iBRAF promotes neural differentiation. Nat. Cell Biol. 2005, 7, 1113–1117. [Google Scholar] [CrossRef] [PubMed]
- Verrier, L.; Vandromme, M.; Trouche, D. Histone demethylases in chromatin cross-talks. Biol. Cell 2011, 103, 381–401. [Google Scholar] [CrossRef]
- Musri, M.; Carmona, M.; Hanzu, F.; Kaliman, P.; Gomis, R.; Párrizas, M. Histone demethylase LSD1 regulates adipogenesis. J. Biol. Chem. 2010, 285, 30034–30041. [Google Scholar] [CrossRef]
- Ridgeway, A.G.; Wilton, S.; Skerjanc, I.S. Myocyte enhancer factor 2C and myogenin up-regulate each other's expression and induce the development of skeletal muscle in P19 cells. J. Biol. Chem. 2000, 275, 41–46. [Google Scholar] [CrossRef]
- Fujii, T.; Murata, K.; Mun, S.; Bae, S.; Lee, Y.; Pannellini, T.; Kang, K.; Oliver, D.; Park-Min, K.; Ivashkiv, L. MEF2C regulates osteoclastogenesis and pathologic bone resorption via c-FOS. Bone Res. 2021, 9, 4. [Google Scholar] [CrossRef]
- Lorenzo, P.; Martin Vazquez, E.; López-Noriega, L.; Fuente-Martín, E.; Mellado-Gil, J.; Franco, J.; Cobo-Vuilleumier, N.; Guerrero Martínez, J.; Romero-Zerbo, S.; Perez-Cabello, J.; et al. The metabesity factor HMG20A potentiates astrocyte survival and reactive astrogliosis preserving neuronal integrity. Theranostics 2021, 11, 6983–7004. [Google Scholar] [CrossRef]
- Nakao, S.; Yamaguchi, K.; Iwamoto, H.; Sakamoto, S.; Horimasu, Y.; Masuda, T.; Miyamoto, S.; Nakashima, T.; Ohshimo, S.; Fujitaka, K.; et al. Serum high-mobility group box 1 as a predictive marker for cytotoxic chemotherapy-induced lung injury in patients with lung cancer and interstitial lung disease. Respir. Med. 2020, 172, 106131. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.; Li, X. Expression and Prognosis Analysis of SUMOylation Regulators in Oral Squamous Cell Carcinoma Based on High-Throughput Sequencing. Front. Genet. 2021, 12, 671392. [Google Scholar] [CrossRef] [PubMed]
- Pan, R.; Yu, D.; Hu, J.; Yang, X.; Wang, C.; Zhang, L.; Xue, P.; Sun, J.; Zhang, X.; Cai, W. SFMBT1 facilitates colon cancer cell metastasis and drug resistance combined with HMG20A. Cell Death Discov. 2022, 8, 263. [Google Scholar] [CrossRef] [PubMed]
- Panda, A.; Abdelmohsen, K.; Yoon, J.; Martindale, J.; Yang, X.; Curtis, J.; Mercken, E.; Chenette, D.; Zhang, Y.; Schneider, R.; et al. RNA-binding protein AUF1 promotes myogenesis by regulating MEF2C expression levels. Mol. Cell. Biol. 2014, 34, 3106–3119. [Google Scholar] [CrossRef]
- Piasecka, A.; Sekrecki, M.; Szcześniak, M.; Sobczak, K. MEF2C shapes the microtranscriptome during differentiation of skeletal muscles. Sci. Rep. 2021, 11, 3476. [Google Scholar] [CrossRef] [PubMed]
- Loumaye, A.; Lause, P.; Zhong, X.; Zimmers, T.; Bindels, L.; Thissen, J. Activin A Causes Muscle Atrophy through MEF2C-Dependent Impaired Myogenesis. Cells 2022, 11, 1119. [Google Scholar] [CrossRef]
- Zlabinger, K.; Spannbauer, A.; Traxler, D.; Gugerell, A.; Lukovic, D.; Winkler, J.; Mester-Tonczar, J.; Podesser, B.; Gyöngyösi, M. MiR-21, MiR-29a, GATA4, and MEF2c Expression Changes in Endothelin-1 and Angiotensin II Cardiac Hypertrophy Stimulated Isl-1Sca-1c-kit Porcine Cardiac Progenitor Cells In Vitro. Cells 2019, 8, 1416. [Google Scholar] [CrossRef]
- Zhang, R.; Sui, L.; Hong, X.; Yang, M.; Li, W. MiR-448 promotes vascular smooth muscle cell proliferation and migration in through directly targeting MEF2C. Environ. Sci. Pollut. Res. Int. 2017, 24, 22294–22300. [Google Scholar] [CrossRef]
- Chen, Y.; Su, Y.; Kung, J. B Cell Development sans B Cell Receptor Responsiveness Due to Unfolded Protein Response-Triggered Mef2c Protein Degradation. J. Immunol. 2018, 201, 2885–2898. [Google Scholar] [CrossRef]
- Colomer-Lahiguera, S.; Pisecker, M.; König, M.; Nebral, K.; Pickl, W.F.; Kauer, M.O.; Haas, O.A.; Ullmann, R.; Attarbaschi, A.; Dworzak, M.N.; et al. MEF2C-dysregulated pediatric T-cell acute lymphoblastic leukemia is associated with CDKN1B deletions and a poor response to glucocorticoid therapy. Leuk. Lymphoma 2017, 58, 2895–2904. [Google Scholar] [CrossRef]
- Sharma, G.; Goalstone, M. Dominant negative FTase (DNFTalpha) inhibits ERK5, MEF2C and CREB activation in adipogenesis. Mol. Cell. Endocrinol. 2005, 245, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Kim, J.; Zhang, R.; Yang, X.; Zhang, Y.; Fang, J.; Chen, Z.; Teng, L.; Chen, X.; Ge, H.; et al. Histone Demethylase LSD1 Promotes Adipocyte Differentiation through Repressing Wnt Signaling. Cell Chem. Biol. 2016, 23, 1228–1240. [Google Scholar] [CrossRef] [PubMed]
- Sambeat, A.; Gulyaeva, O.; Dempersmier, J.; Tharp, K.; Stahl, A.; Paul, S.; Sul, H. LSD1 Interacts with Zfp516 to Promote UCP1 Transcription and Brown Fat Program. Cell Rep. 2016, 15, 2536–2549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, M.; Park, U.; Kim, J.; Choi, H.; Um, S.; Kim, E. CACUL1 reciprocally regulates SIRT1 and LSD1 to repress PPARγ and inhibit adipogenesis. Cell Death Dis. 2017, 8, 3201. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Ojeda, F.; Rupérez, A.; Gomez-Llorente, C.; Gil, A.; Aguilera, C. Cell Models and Their Application for Studying Adipogenic Differentiation in Relation to Obesity: A Review. Int. J. Mol. Sci. 2016, 17, 1040. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sequence Name | Sequence | |
|---|---|---|
| Negative Control | F: | UUCUCCGAACGUGUCACGUTT |
| R: | ACGUGACACGUUCGGAGAATT | |
| sus-siHMG20A-1 | F: | CCACUAACAAUCCAGAAUUTT |
| R: | AAUUCUGGAUUGUUAGUGGTT | |
| sus-siHMG20A-2 | F: | GGAGCGUUACAUGAAGGAATT |
| R: | UUCCUUCAUGUAACGGUCCTT | |
| sus-siHMG20A-3 | F: | CCCUAUAUUUACAGAGGAATT |
| R: | UUCCUCUGUAAAUAUAGGGTT | |
| mus-siHMG20A-1 | F: | GACCGUCAGAAAGGCAAAUTT |
| R: | AUUUGCCUUUCUGACGGUCTT | |
| mus-siHMG20A-2 | F: | GCCUGGAAGUGGAGAAAUATT |
| R: | UAUUUCUCCACUUCCAGGCTT | |
| mus-si-MEF2C | F: | CACCUGGUAACCUGAACAATT |
| R: | UUGUUCAGGUUACCAGGUGTT | |
| mus-si-LSD1 | F: | CCCAAAGAUCCAGCUGACGUUUGAA |
| R: | UUCAAACGUCAGCUGGAUCUUUGGG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, R.; Meng, S.; Ji, M.; Rong, X.; You, Z.; Cai, C.; Guo, X.; Lu, C.; Liang, G.; Cao, G.; et al. HMG20A Inhibit Adipogenesis by Transcriptional and Epigenetic Regulation of MEF2C Expression. Int. J. Mol. Sci. 2022, 23, 10559. https://doi.org/10.3390/ijms231810559
Li R, Meng S, Ji M, Rong X, You Z, Cai C, Guo X, Lu C, Liang G, Cao G, et al. HMG20A Inhibit Adipogenesis by Transcriptional and Epigenetic Regulation of MEF2C Expression. International Journal of Molecular Sciences. 2022; 23(18):10559. https://doi.org/10.3390/ijms231810559
Chicago/Turabian StyleLi, Ruixiao, Shan Meng, Mengting Ji, Xiaoyin Rong, Ziwei You, Chunbo Cai, Xiaohong Guo, Chang Lu, Guoming Liang, Guoqing Cao, and et al. 2022. "HMG20A Inhibit Adipogenesis by Transcriptional and Epigenetic Regulation of MEF2C Expression" International Journal of Molecular Sciences 23, no. 18: 10559. https://doi.org/10.3390/ijms231810559
APA StyleLi, R., Meng, S., Ji, M., Rong, X., You, Z., Cai, C., Guo, X., Lu, C., Liang, G., Cao, G., Li, B., & Yang, Y. (2022). HMG20A Inhibit Adipogenesis by Transcriptional and Epigenetic Regulation of MEF2C Expression. International Journal of Molecular Sciences, 23(18), 10559. https://doi.org/10.3390/ijms231810559