
Novel Dopamine Transporter Inhibitor, CE-123, Ameliorates Spatial Memory Deficits Induced by Maternal Separation in Adolescent Rats: Impact of Sex
 , , ,
, , ,  , ,
, , 
Abstract
:1. Introduction
2. Results
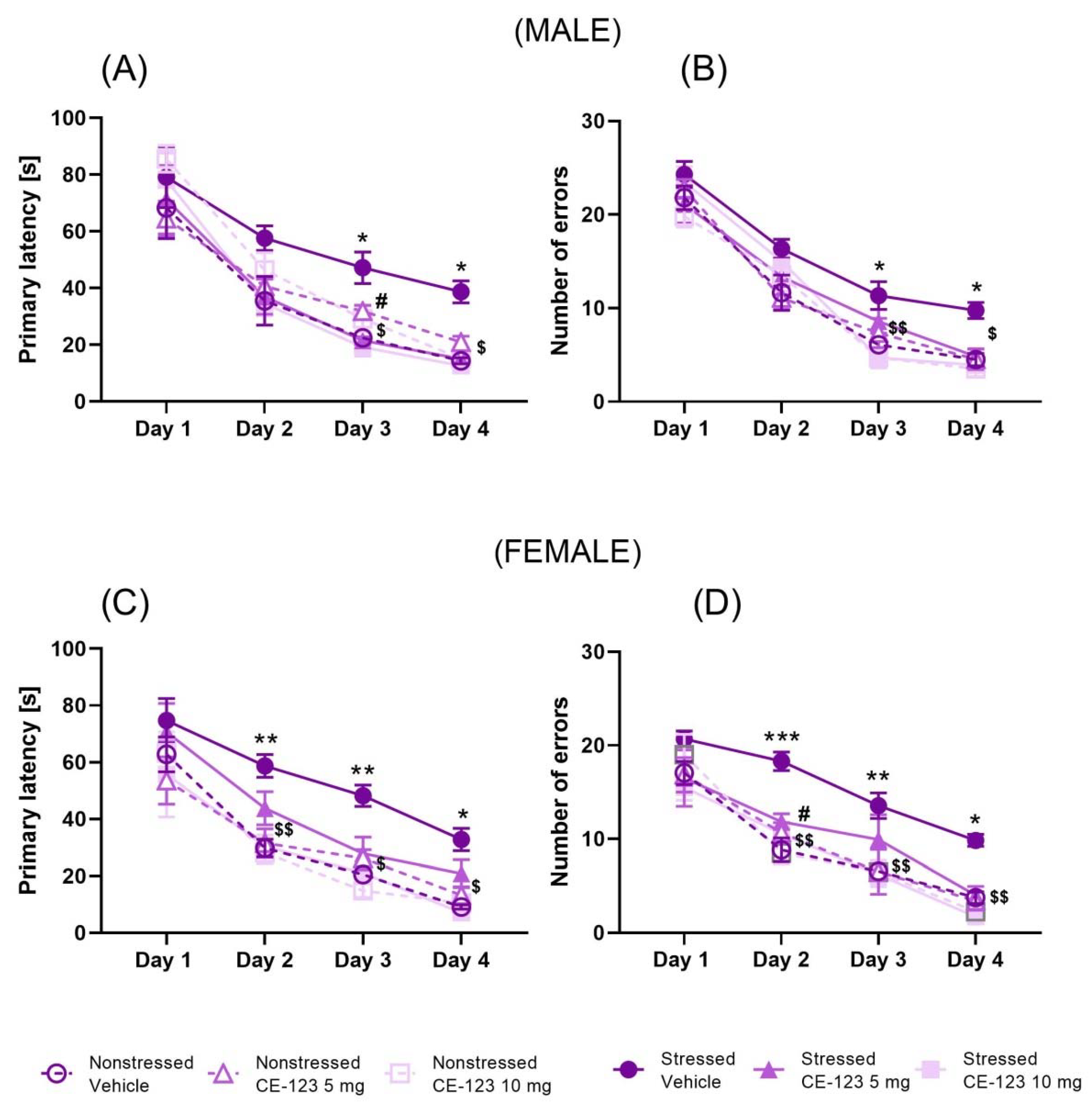
2.1. Effect of CE-123 on Acquisition of Spatial Learning of the Barnes Maze in Young Male and Female Rats with Prior MS
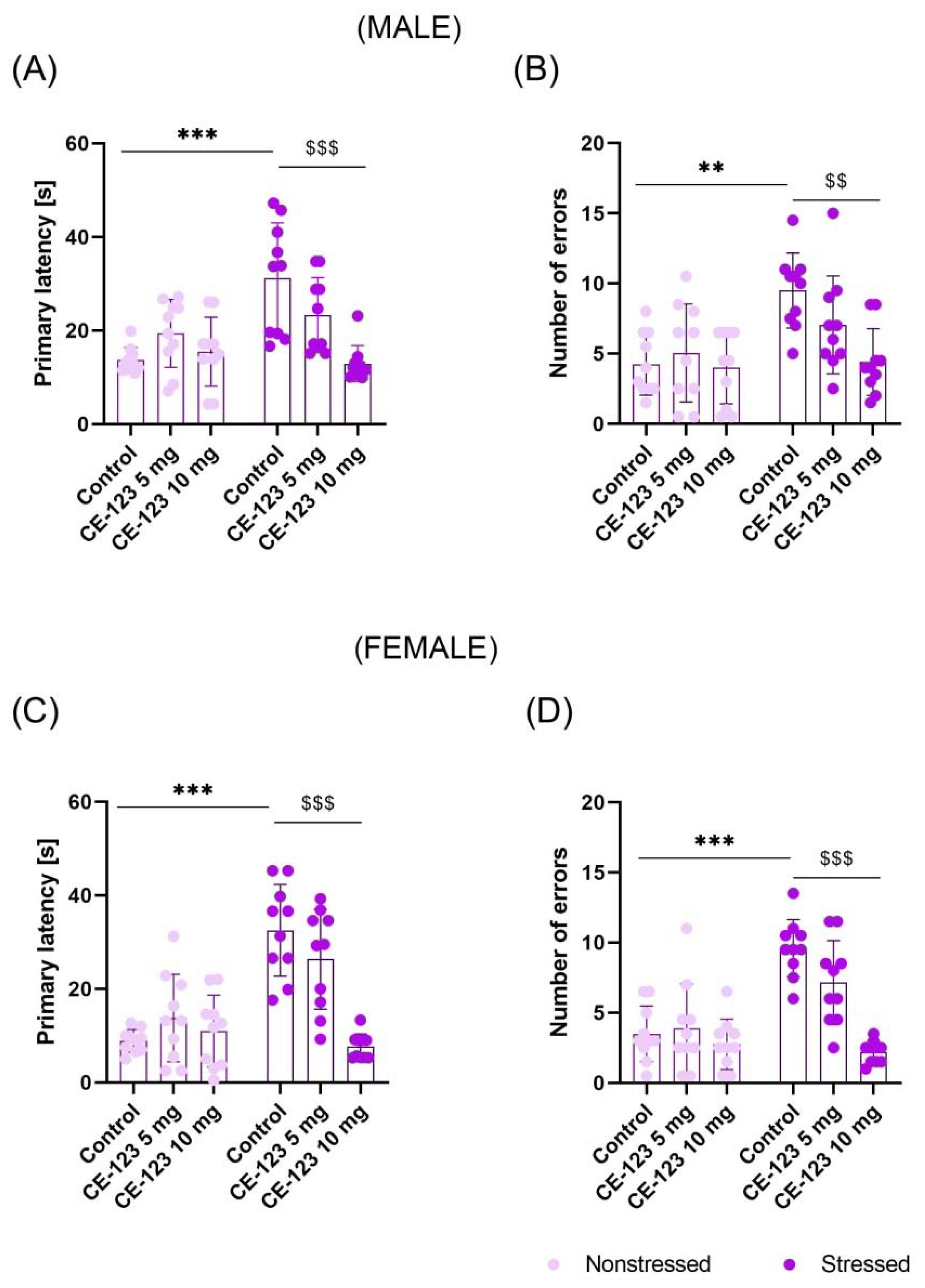
2.2. Effect of CE-123 Given during Acquisition Session on Spatial Memory Retention in the Probe-Trial of the Barnes Maze in Young Male and Female Rats with Prior MS
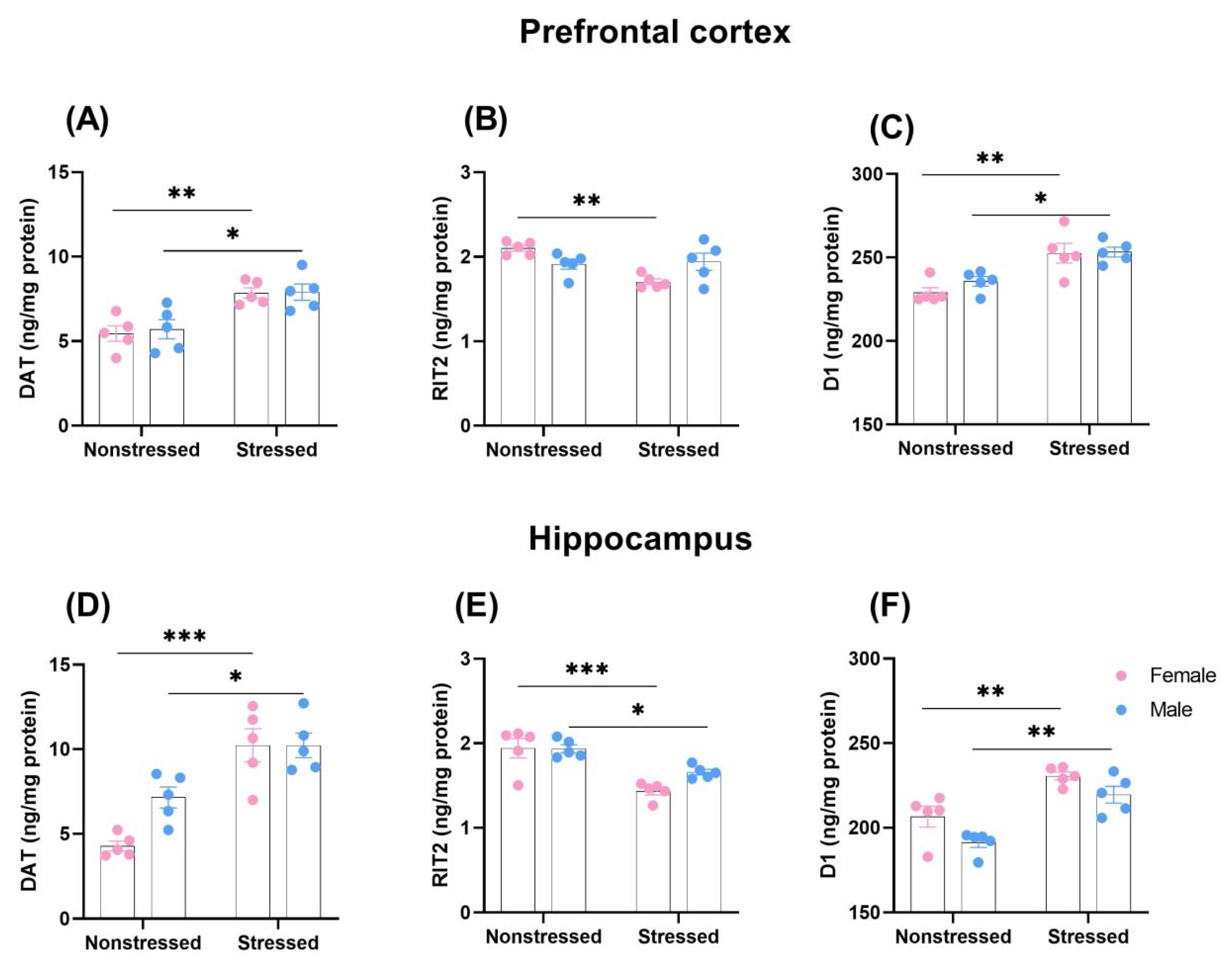
2.3. The Influence of Early MS on Dopamine Transporter (DAT), GTP-Binding Protein RIT2, and D1 Receptor Expression in the Prefrontal Cortex and Hippocampus in Adolescent Male and Female Rats
2.3.1. Prefrontal Cortex
2.3.2. Hippocampus
2.4. The Influence of CE-123 Administration before Each Acquisition Session on D1 Receptor Expression Level in the Hippocampus of MS Male and Female Adolescent Rats
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Drugs
4.3. Maternal Separation
4.4. Barnes Maze Task
4.4.1. Habituation
4.4.2. Acquisition Phase
4.4.3. Probe-Trial
4.5. ELISA Assays
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lupien, S.J.; McEwen, B.S.; Gunnar, M.R.; Heim, C. Effects of stress throughout the lifespan on the brain, behaviour and cognition. Nat. Rev. Neurosci. 2009, 10, 434–445. [Google Scholar] [CrossRef] [PubMed]
- Huot, R.L.; Plotsky, P.M.; Lenox, R.H.; McNamara, R.K. Neonatal maternal separation reduces hippocampal mossy fiber density in adult Long Evans rats. Brain Res. 2002, 950, 52–63. [Google Scholar] [CrossRef]
- Matthews, K.; Robbins, T.W. Early experience as a determinant of adult behavioural responses to reward: The effects of repeated maternal separation in the rat. Neurosci. Biobehav. Rev. 2003, 27, 45–55. [Google Scholar] [CrossRef]
- Ploj, K.; Roman, E.; Nylander, I. Long-term effects of short and long periods of maternal separation on brain opioid peptide levels in male Wistar rats. Neuropeptides 2003, 37, 149–156. [Google Scholar] [CrossRef]
- Pachenari, N.; Azizi, H.; Ghasemi, E.; Azadi, M.; Semnanian, S. Exposure to opiates in male adolescent rats alters pain perception in the male offspring. Behav. Pharmacol. 2018, 29, 255–260. [Google Scholar] [CrossRef]
- Goodman, J.B.; Freeman, E.E.; Chalmers, K.A. The relationship between early life stress and working memory in adulthood: A systematic review and meta-analysis. Memory 2019, 27, 868–880. [Google Scholar] [CrossRef]
- Pechtel, P.; Pizzagalli, D.A. Effects of early life stress on cognitive and affective function: An integrated review of human literature. Psychopharmacology 2011, 214, 55–70. [Google Scholar] [CrossRef]
- Paterniti, S.; Sterner, I.; Caldwell, C.; Bisserbe, J.C. Childhood neglect predicts the course of major depression in a tertiary care sample: A follow-up study. BMC Psychiatry 2017, 17, 113. [Google Scholar] [CrossRef]
- Wigger, A.; Neumann, I.D. Periodic maternal deprivation induces gender-dependent alterations in behavioral and neuroendocrine responses to emotional stress in adult rats. Physiol. Behav. 1999, 66, 293–302. [Google Scholar] [CrossRef]
- Kalinichev, M.; Easterling, K.W.; Plotsky, P.M.; Holtzman, S.G. Long-lasting changes in stress-induced corticosterone response and anxiety-like behaviors as a consequence of neonatal maternal separation in Long-Evans rats. Pharmacol. Biochem. Behav. 2002, 73, 131–140. [Google Scholar] [CrossRef]
- Muhammad, A.; Kolb, B. Maternal separation altered behavior and neuronal spine density without influencing amphetamine sensitization. Behav. Brain Res. 2011, 223, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, M.C.; Ogawa, S. Long-lasting consequences of neonatal maternal separation on social behaviors in ovariectomized female mice. PLoS ONE 2012, 7, e33028. [Google Scholar] [CrossRef] [PubMed]
- Niwa, M.; Matsumoto, Y.; Mouri, A.; Ozaki, N.; Nabeshima, T. Vulnerability in early life to changes in the rearing environment plays a crucial role in the aetiopathology of psychiatric disorders. Int. J. Neuropsychopharmacol. 2011, 14, 459–477. [Google Scholar] [CrossRef] [PubMed]
- Thomas, A.W.; Caporale, N.; Wu, C.; Wilbrecht, L. Early maternal separation impacts cognitive flexibility at the age of first independence in mice. Dev. Cogn. Neurosci. 2016, 18, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Chiba, A.A.; Kesner, R.P.; Reynolds, A.M. Memory for spatial location as a function of temporal lag in rats: Role of hippocampus and medial prefrontal cortex. Behav. Neural. Biol. 1994, 61, 123–131. [Google Scholar] [CrossRef]
- Morris, R.G.; Garrud, P.; Rawlins, J.N.; O’Keefe, J. Place navigation impaired in rats with hippocampal lesions. Nature 1982, 297, 681–683. [Google Scholar] [CrossRef]
- Fuge, P.; Aust, S.; Fan, Y.; Weigand, A.; Gärtner, M.; Feeser, M.; Bajbouj, M.; Grimm, S. Interaction of early life stress and corticotropin-releasing hormone receptor gene: Effects on working memory. Biol. Psychiatry 2014, 76, 888–894. [Google Scholar] [CrossRef]
- O’Keefe, J. Do hippocampal pyramidal cells signal non-spatial as well as spatial information? Hippocampus 1999, 9, 352–364. [Google Scholar] [CrossRef]
- Muzzio, I.A.; Kentros, C.; Kandel, E. What is remembered? Role of attention on the encoding and retrieval of hippocampal representations. J. Physiol. 2009, 587, 2837–2854. [Google Scholar] [CrossRef]
- Kentros, C.G.; Agnihotri, N.T.; Streater, S.; Hawkins, R.D.; Kandel, E.R. Increased attention to spatial context increases both place field stability and spatial memory. Neuron 2004, 42, 283–295. [Google Scholar] [CrossRef] [Green Version]
- Gasbarri, A.; Sulli, A.; Innocenzi, R.; Pacitti, C.; Brioni, J.D. Spatial memory impairment induced by lesion of the mesohippocampal dopaminergic system in the rat. Neuroscience 1996, 74, 1037–1044. [Google Scholar] [CrossRef]
- Mizoguchi, K.; Yuzurihara, M.; Ishige, A.; Sasaki, H.; Chui, D.H.; Tabira, T. Chronic stress induces impairment of spatial working memory because of prefrontal dopaminergic dysfunction. J. Neurosci. 2000, 20, 1568–1574. [Google Scholar] [CrossRef] [PubMed]
- Broussard, J.I.; Yang, K.; Levine, A.T.; Tsetsenis, T.; Jenson, D.; Cao, F.; Garcia, I.; Arenkiel, B.R.; Zhou, F.M.; De Biasi, M.; et al. Dopamine regulates aversive contextual learning and associated in vivo synaptic plasticity in the hippocampus. Cell Rep. 2016, 14, 1930–1939. [Google Scholar] [CrossRef] [PubMed]
- Hansen, N.; Manahan-Vaughan, D. Dopamine D1/D5 receptors mediate informational saliency that promotes persistent hippocampal long-term plasticity. Cereb. Cortex. 2014, 24, 845–858. [Google Scholar] [CrossRef] [PubMed]
- Kempadoo, K.A.; Mosharov, E.V.; Choi, S.J.; Sulzer, D.; Kandel, E.R. Dopamine release from the locus coeruleus to the dorsal hippocampus promotes spatial learning and memory. Proc. Natl. Acad. Sci. USA 2016, 113, 14835–14840. [Google Scholar] [CrossRef]
- El-Ghundi, M.; O’Dowd, B.F.; George, S.R. Insights into the role of dopamine receptor systems in learning and memory. Rev. Neurosci. 2007, 18, 37–66. [Google Scholar] [CrossRef]
- da Silva, W.C.; Köhler, C.C.; Radiske, A.; Cammarota, M. D1/D5 dopamine receptors modulate spatial memory formation. Neurobiol. Learn Mem. 2012, 97, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Xing, B.; Kong, H.; Meng, X.; Wei, S.G.; Xu, M.; Li, S.B. Dopamine D1 but not D3 receptor is critical for spatial learning and related signaling in the hippocampus. Neuroscience 2010, 169, 1511–1519. [Google Scholar] [CrossRef]
- Karunakaran, S.; Chowdhury, A.; Donato, F.; Quairiaux, C.; Michel, C.M.; Caroni, P. PV plasticity sustained through D1/5 dopamine signaling required for long-term memory consolidation. Nat. Neurosci. 2016, 19, 454–464. [Google Scholar] [CrossRef]
- Williams, G.V.; Castner, S.A. Under the curve: Critical issues for elucidating D1 receptor function in working memory. Neuroscience 2006, 139, 263–276. [Google Scholar] [CrossRef]
- Balderas, I.; Moreno-Castilla, P.; Bermudez-Rattoni, F. Dopamine D1 receptor activity modulates object recognition memory consolidation in the perirhinal cortex but not in the hippocampus. Hippocampus 2013, 23, 873–878. [Google Scholar] [CrossRef] [PubMed]
- Hotte, M.; Thuault, S.; Lachaise, F.; Dineley, K.T.; Hemmings, H.C.; Nairn, A.C.; Jay, T.M. D1 receptor modulation of memory retrieval performance is associated with changes in pCREB and pDARPP-32 in rat prefrontal cortex. Behav. Brain Res. 2006, 171, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Rossato, J.I.; Radiske, A.; Kohler, C.A.; Gonzalez, C.; Bevilaqua, L.R.; Medina, J.H.; Cammarota, M. Consolidation of object recognition memory requires simultaneous activation of dopamine D1/D5 receptors in the amygdala and medial prefrontal cortex but not in the hippocampus. Neurobiol. Learn Mem. 2013, 106, 66–70. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, A.S.; Andersen, J.; Jørgensen, T.N.; Sørensen, L.; Eriksen, J.; Loland, C.J.; Strømgaard, K.; Gether, U. SLC6 neurotransmitter transporters: Structure, function, and regulation. Pharmacol. Rev. 2011, 63, 585–640. [Google Scholar] [CrossRef] [PubMed]
- Gainetdinov, R.R.; Jones, S.R.; Fumagalli, F.; Wightman, R.M.; Caron, M.G. Re-evaluation of the role of the dopamine transporter in dopamine system homeostasis. Brain Res. Brain Res. Rev. 1998, 26, 148–153. [Google Scholar] [CrossRef]
- Kurzina, N.P.; Aristova, I.Y.; Volnova, A.B.; Gainetdinov, R.R. Deficit in working memory and abnormal behavioral tactics in dopamine transporter knockout rats during training in the 8-arm maze. Behav. Brain Res. 2020, 390, 112642. [Google Scholar] [CrossRef]
- Efimova, E.V.; Gainetdinov, R.R.; Budygin, E.A.; Sotnikova, T.D. Dopamine transporter mutant animals: A translational perspective. J. Neurogenet. 2016, 30, 5–15. [Google Scholar] [CrossRef]
- Battleday, R.M.; Brem, A.K. Modafinil for cognitive neuroenhancement in healthy non-sleep-deprived subjects: A systematic review. Eur. Neuropsychopharmacol. 2015, 25, 1865–1881. [Google Scholar] [CrossRef]
- Bobo, W.V.; Woodward, N.D.; Sim, M.Y.; Jayathilake, K.; Meltzer, H.Y. The effect of adjunctive armodafinil on cognitive performance and psychopathology in antipsychotic-treated patients with schizophrenia/schizoaffective disorder: A randomized, double-blind, placebo-controlled trial. Schizophr. Res. 2011, 130, 106–113. [Google Scholar] [CrossRef]
- Dolder, P.C.; Müller, F.; Schmid, Y.; Borgwardt, S.J.; Liechti, M.E. Direct comparison of the acute subjective, emotional, autonomic, and endocrine effects of MDMA, methylphenidate, and modafinil in healthy subjects. Psychopharmacology 2018, 235, 467–479. [Google Scholar] [CrossRef] [Green Version]
- Alam, N.; Choudhary, K. Haloperidol attenuates methylphenidate and modafinil induced behavioural sensitization and cognitive enhancement. Metab. Brain Dis. 2018, 33, 893–906. [Google Scholar] [CrossRef] [PubMed]
- Robinson, T.E.; Browman, K.E.; Crombag, H.S.; Badiani, A. Modulation of the induction or expression of psychostimulant sensitization by the circumstances surrounding drug administration. Neurosci. Biobehav. Rev. 1998, 22, 347–354. [Google Scholar] [CrossRef]
- Gerrard, P.; Malcolm, R. Mechanisms of modafinil: A review of current research. Neuropsychiatr. Dis. Treat. 2007, 3, 349–364. [Google Scholar]
- Kalaba, P.; Aher, N.Y.; Ilić, M.; Dragačević, V.; Wieder, M.; Miklosi, A.G.; Zehl, M.; Wackerlig, J.; Roller, A.; Beryozkina, T.; et al. Heterocyclic analogues of modafinil as novel, atypical dopamine transporter inhibitors. J. Med. Chem. 2017, 60, 9330–9348. [Google Scholar] [CrossRef] [PubMed]
- Kalaba, P.; Ilić, M.; Aher, N.Y.; Dragačević, V.; Wieder, M.; Zehl, M.; Wackerlig, J.; Beyl, S.; Sartori, S.B.; Ebner, K.; et al. Structure-activity relationships of novel thiazole-based modafinil analogues acting at monoamine transporters. J. Med. Chem. 2020, 63, 391–417. [Google Scholar] [CrossRef]
- Nikiforuk, A.; Kalaba, P.; Ilic, M.; Korz, V.; Dragačević, V.; Wackerlig, J.; Langer, T.; Höger, H.; Golebiowska, J.; Popik, P.; et al. A novel dopamine transporter inhibitor CE-123 improves cognitive flexibility and maintains impulsivity in healthy male rats. Front. Behav. Neurosci. 2017, 11, 222. [Google Scholar] [CrossRef]
- Kristofova, M.; Aher, Y.D.; Ilic, M.; Radoman, B.; Kalaba, P.; Dragacevic, V.; Aher, N.Y.; Leban, J.; Korz, V.; Zanon, L.; et al. A daily single dose of a novel modafinil analogue CE-123 improves memory acquisition and memory retrieval. Behav. Brain Res. 2018, 343, 83–94. [Google Scholar] [CrossRef]
- Gibula-Tarlowska, E.; Korz, V.; Lopatynska-Mazurek, M.; Chlopas-Konowalek, A.; Grochecki, P.; Kalaba, P.; Dragacevic, V.; Kotlinski, R.; Kujawski, R.; Szulc, M.; et al. CE-123, a novel dopamine transporter inhibitor, attenuates locomotor hyperactivity and improves cognitive functions in rat model of fetal alcohol spectrum disorders. Behav. Brain Res. 2021, 410, 113326. [Google Scholar] [CrossRef]
- Sagheddu, C.; Pintori, N.; Kalaba, P.; Dragačević, V.; Piras, G.; Lubec, J.; Simola, N.; De Luca, M.A.; Lubec, G.; Pistis, M. Neurophysiological and neurochemical effects of the putative cognitive enhancer (S)-CE-123 on mesocorticolimbic dopamine system. Biomolecules 2020, 10, 779. [Google Scholar] [CrossRef]
- Lee, C.H.; Della, N.G.; Chew, C.E.; Zack, D.J. Rin, a neuron-specific and calmodulin-binding small G-protein, and Rit define a novel subfamily of ras proteins. J. Neurosci. 1996, 16, 6784–6794. [Google Scholar] [CrossRef]
- Navaroli, D.M.; Stevens, Z.H.; Uzelac, Z.; Gabriel, L.; King, M.J.; Lifshitz, L.M.; Sitte, H.H.; Melikian, H.E. The plasma membrane-associated GTPase Rin interacts with the dopamine transporter and is required for protein kinase C-regulated dopamine transporter trafficking. J. Neurosci. 2011, 31, 13758–13770. [Google Scholar] [CrossRef] [PubMed]
- Fagan, R.R.; Kearney, P.J.; Sweeney, C.G.; Luethi, D.; Schoot Uiterkamp, F.E.; Schicker, K.; Alejandro, B.S.; O’Connor, L.C.; Sitte, H.H.; Melikian, H.E. Dopamine transporter trafficking and Rit2 GTPase: Mechanism of action and in vivo impact. J. Biol. Chem. 2020, 295, 5229–5244. [Google Scholar] [CrossRef] [PubMed]
- Judo, C.; Matsumoto, M.; Yamazaki, D.; Hiraide, S.; Yanagawa, Y.; Kimura, S.; Shimamura, K.; Togashi, H. Early stress exposure impairs synaptic potentiation in the rat medial prefrontal cortex underlying contextual fear extinction. Neuroscience 2010, 169, 1705–1714. [Google Scholar] [CrossRef] [PubMed]
- Sandi, C.; Pinelo-Nava, M.T. Stress and memory: Behavioral effects and neurobiological mechanisms. Neural. Plast. 2007, 2007, 78970. [Google Scholar] [CrossRef]
- Sousa, V.C.; Vital, J.; Costenla, A.R.; Batalha, V.L.; Sebastião, A.M.; Ribeiro, J.A.; Lopes, L.V. Maternal separation impairs long term-potentiation in CA1-CA3 synapses and hippocampal-dependent memory in old rats. Neurobiol. Aging. 2014, 35, 1680–1685. [Google Scholar] [CrossRef]
- Sun, X.; Zhang, Y.; Li, X.; Liu, X.; Qin, C. Early-life neglect alters emotional and cognitive behavior in a sex-dependent manner and reduces glutamatergic neuronal excitability in the prefrontal cortex. Front Psychiatry 2021, 11, 572224. [Google Scholar] [CrossRef]
- Filarowska-Jurko, J.; Komsta, L.; Smaga, I.; Surowka, P.; Marszalek-Grabska, M.; Grochecki, P.; Nizio, D.; Filip, M.; Kotlinska, J.H. Maternal separation alters ethanol drinking and reversal learning processes in adolescent rats: The impact of sex and glycine transporter type 1 (GlyT1) inhibitor. Int. J. Mol. Sci. 2022, 23, 5350. [Google Scholar] [CrossRef]
- Oomen, C.A.; Girardi, C.E.; Cahyadi, R.; Verbeek, E.C.; Krugers, H.; Joëls, M.; Lucassen, P.J. Opposite effects of early maternal deprivation on neurogenesis in male versus female rats. PLoS ONE 2009, 4, e3675. [Google Scholar] [CrossRef]
- Loi, M.; Koricka, S.; Lucassen, P.J.; Joëls, M. Age- and sex-dependent effects of early life stress on hippocampal neurogenesis. Front. Endocrinol. 2014, 5, 13. [Google Scholar] [CrossRef]
- Kosten, T.A.; Kim, J.J.; Lee, H.J. Early life manipulations alter learning and memory in rats. Neurosci. Biobehav. Rev. 2012, 36, 1985–2006. [Google Scholar] [CrossRef]
- Braun, K.; Bock, J.; Wainstock, T.; Matas, E.; Gaisler-Salomon, I.; Fegert, J.; Ziegenhain, U.; Segal, M. Experience-induced transgenerational (re-)programming of neuronal structure and functions: Impact of stress prior and during pregnancy. Neurosci. Biobehav. Rev. 2020, 117, 281–296. [Google Scholar] [CrossRef] [PubMed]
- Bock, J.; Wainstock, T.; Braun, K.; Segal, M. Stress in utero: Prenatal programming of brain plasticity and cognition. Biol. Psychiatry 2015, 78, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Matas, E.; Bock, J.; Braun, K. The impact of parent-infant interaction on epigenetic plasticity mediating synaptic adaptations in the infant brain. Psychopathology 2016, 49, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Kunzler, J.; Braun, K.; Bock, J. Early life stress and sex-specific sensitivity of the catecholaminergic systems in prefrontal and limbic regions of Octodon degus. Brain Struct. Funct. 2015, 220, 861–868. [Google Scholar] [CrossRef] [PubMed]
- Ziabreva, I.; Schnabel, R.; Poeggel, G.; Braun, K. Mother’s voice “buffers” separation-induced receptor changes in the prefrontal cortex of octodon degus. Neuroscience 2003, 119, 433–441. [Google Scholar] [CrossRef]
- Kasanova, Z.; Hernaus, D.; Vaessen, T.; van Amelsvoort, T.; Winz, O.; Heinzel, A.; Pruessner, J.; Mottaghy, F.M.; Collip, D.; Myin-Germeys, I. Early-life stress affects stress-related prefrontal dopamine activity in healthy adults, but not in individuals with psychotic disorder. PLoS ONE 2016, 11, e0150746. [Google Scholar] [CrossRef]
- Majcher-Maślanka, I.; Solarz, A.; Wędzony, K.; Chocyk, A. The effects of early-life stress on dopamine system function in adolescent female rats. Int. J. Dev. Neurosci. 2017, 57, 24–33. [Google Scholar] [CrossRef]
- Wang, M.; Datta, D.; Enwright, J.; Galvin, V.; Yang, S.T.; Paspalas, C.; Kozak, R.; Gray, D.L.; Lewis, D.A.; Arnsten, A.F.T. A novel dopamine D1 receptor agonist excites delay-dependent working memory-related neuronal firing in primate dorsolateral prefrontal cortex. Neuropharmacology 2019, 150, 46–58. [Google Scholar] [CrossRef]
- Rentesi, G.; Antoniou, K.; Marselos, M.; Syrrou, M.; Papadopoulou-Daifoti, Z.; Konstandi, M. Early maternal deprivation-induced modifications in the neurobiological, neurochemical and behavioral profile of adult rats. Behav. Brain Res. 2013, 244, 29–37. [Google Scholar] [CrossRef]
- Banqueri, M.; Gutiérrez-Menéndez, A.; Méndez, M.; Conejo, N.M.; Arias, J.L. Early life stress due to repeated maternal separation alters the working memory acquisition brain functional network. Stress 2021, 24, 87–95. [Google Scholar] [CrossRef]
- Son, G.H.; Chung, S.; Geum, D.; Kang, S.S.; Choi, W.S.; Kim, K.; Choi, S. Hyperactivity and alteration of the midbrain dopaminergic system in maternally stressed male mice offspring. Biochem. Biophys. Res. Commun. 2007, 352, 823–829. [Google Scholar] [CrossRef] [PubMed]
- Converse, A.K.; Moore, C.F.; Moirano, J.M.; Ahlers, E.O.; Larson, J.A.; Engle, J.W.; Barnhart, T.E.; Murali, D.; Christian, B.T.; DeJesus, O.T.; et al. Prenatal stress induces increased striatal dopamine transporter binding in adult nonhuman primates. Biol. Psychiatry 2013, 74, 502–510. [Google Scholar] [CrossRef] [PubMed]
- Van den Bergh, B.R.; Marcoen, A. High antenatal maternal anxiety is related to ADHD symptoms, externalizing problems, and anxiety in 8- and 9-year-olds. Child Dev. 2004, 75, 1085–1097. [Google Scholar] [CrossRef] [PubMed]
- Spencer, T.J.; Biederman, J.; Mick, E. Attention-deficit/hyperactivity disorder: Diagnosis, lifespan, comorbidities, and neurobiology. J. Pediatr. Psychol. 2007, 32, 631–642. [Google Scholar] [CrossRef] [PubMed]
- Volkow, N.D.; Wang, G.J.; Kollins, S.H.; Wigal, T.L.; Newcorn, J.H.; Telang, F.; Fowler, J.S.; Zhu, W.; Logan, J.; Ma, Y.; et al. Evaluating dopamine reward pathway in ADHD: Clinical implications. JAMA 2009, 302, 1084–1091. [Google Scholar] [CrossRef]
- Bannon, M.J. The dopamine transporter: Role in neurotoxicity and human disease. Toxicol. Appl. Pharmacol. 2005, 204, 355–360. [Google Scholar] [CrossRef]
- Turner, D. A review of the use of modafinil for attention-deficit hyperactivity disorder. Expert Rev. Neurother. 2006, 6, 455–468. [Google Scholar] [CrossRef]
- Sweeney, C.G.; Kearney, P.J.; Fagan, R.R.; Smith, L.A.; Bolden, N.C.; Zhao-Shea, R.; Rivera, I.V.; Kolpakova, J.; Xie, J.; Gao, G.; et al. Conditional, inducible gene silencing in dopamine neurons reveals a sex-specific role for Rit2 GTPase in acute cocaine response and striatal function. Neuropsychopharmacology 2020, 45, 384–393. [Google Scholar] [CrossRef]
- Chocyk, A.; Dudys, D.; Przyborowska, A.; Maćkowiak, M.; Wędzony, K. Impact of maternal separation on neural cell adhesion molecules expression in dopaminergic brain regions of juvenile, adolescent and adult rats. Pharmacol. Rep. 2010, 62, 1218–1224. [Google Scholar] [CrossRef]
- Grochecki, P.; Smaga, I.; Lopatynska-Mazurek, M.; Gibula-Tarlowska, E.; Kedzierska, E.; Listos, J.; Talarek, S.; Marszalek-Grabska, M.; Hubalewska-Mazgaj, M.; Korga-Plewko, A.; et al. Effects of mephedrone and amphetamine exposure during adolescence on spatial memory in adulthood: Behavioral and neurochemical analysis. Int. J. Mol. Sci. 2021, 22, 589. [Google Scholar] [CrossRef]
- Lopatynska-Mazurek, M.; Antolak, A.; Grochecki, P.; Gibula-Tarlowska, E.; Bodzon-Kulakowska, A.; Listos, J.; Kedzierska, E.; Suder, P.; Silberring, J.; Kotlinska, J.H. Rapamycin improves spatial learning deficits, vulnerability to alcohol addiction and altered expression of the GluN2B subunit of the NMDA receptor in adult rats exposed to ethanol during the neonatal period. Biomolecules 2021, 11, 650. [Google Scholar] [CrossRef] [PubMed]
- Harrison, F.E.; Reiserer, R.S.; Tomarken, A.J.; McDonald, M.P. Spatial and nonspatial escape strategies in the Barnes maze. Learn Mem. 2006, 13, 809–819. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Csaszar, E.; Szodorai, E.; Patil, S.; Pollak, A.; Lubec, G. The differential hippocampal phosphoproteome of Apodemus sylvaticus paralleling spatial memory retrieval in the Barnes maze. Behav. Brain Res. 2014, 264, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Marszalek-Grabska, M.; Gibula-Bruzda, E.; Bodzon-Kulakowska, A.; Suder, P.; Gawel, K.; Talarek, S.; Listos, J.; Kedzierska, E.; Danysz, W.; Kotlinska, J.H. ADX-47273, a mGlu5 receptor positive allosteric modulator, attenuates deficits in cognitive flexibility induced by withdrawal from ‘binge-like’ ethanol exposure in rats. Behav. Brain Res. 2018, 338, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Gawel, K.; Gibula, E.; Marszalek-Grabska, M.; Filarowska, J.; Kotlinska, J.H. Assessment of spatial learning and memory in the Barnes maze task in rodents-methodological consideration. Naunyn Schmiedebergs Arch. Pharmacol. 2019, 392, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| FEMALE | MALE | ||
|---|---|---|---|
| STRESSED | STRESSED + CE 10 mg | STRESSED | STRESSED + CE 10 mg |
| 230.611 ± 6.967 | 162.233 ± 2.354 *** | 219.500 ± 4.962 | 188.406 ± 8.308 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grochecki, P.; Smaga, I.; Surowka, P.; Marszalek-Grabska, M.; Kalaba, P.; Dragacevic, V.; Kotlinska, P.; Filip, M.; Lubec, G.; Kotlinska, J.H. Novel Dopamine Transporter Inhibitor, CE-123, Ameliorates Spatial Memory Deficits Induced by Maternal Separation in Adolescent Rats: Impact of Sex. Int. J. Mol. Sci. 2022, 23, 10718. https://doi.org/10.3390/ijms231810718
Grochecki P, Smaga I, Surowka P, Marszalek-Grabska M, Kalaba P, Dragacevic V, Kotlinska P, Filip M, Lubec G, Kotlinska JH. Novel Dopamine Transporter Inhibitor, CE-123, Ameliorates Spatial Memory Deficits Induced by Maternal Separation in Adolescent Rats: Impact of Sex. International Journal of Molecular Sciences. 2022; 23(18):10718. https://doi.org/10.3390/ijms231810718
Chicago/Turabian StyleGrochecki, Pawel, Irena Smaga, Paulina Surowka, Marta Marszalek-Grabska, Predrag Kalaba, Vladimir Dragacevic, Patrycja Kotlinska, Malgorzata Filip, Gert Lubec, and Jolanta H. Kotlinska. 2022. "Novel Dopamine Transporter Inhibitor, CE-123, Ameliorates Spatial Memory Deficits Induced by Maternal Separation in Adolescent Rats: Impact of Sex" International Journal of Molecular Sciences 23, no. 18: 10718. https://doi.org/10.3390/ijms231810718
APA StyleGrochecki, P., Smaga, I., Surowka, P., Marszalek-Grabska, M., Kalaba, P., Dragacevic, V., Kotlinska, P., Filip, M., Lubec, G., & Kotlinska, J. H. (2022). Novel Dopamine Transporter Inhibitor, CE-123, Ameliorates Spatial Memory Deficits Induced by Maternal Separation in Adolescent Rats: Impact of Sex. International Journal of Molecular Sciences, 23(18), 10718. https://doi.org/10.3390/ijms231810718