
Vaccine-Induced Subcutaneous Granulomas in Goats Reflect Differences in Host–Mycobacterium Interactions between BCG- and Recombinant BCG-Derivative Vaccines
 , and
, and
Abstract
:1. Introduction
2. Results
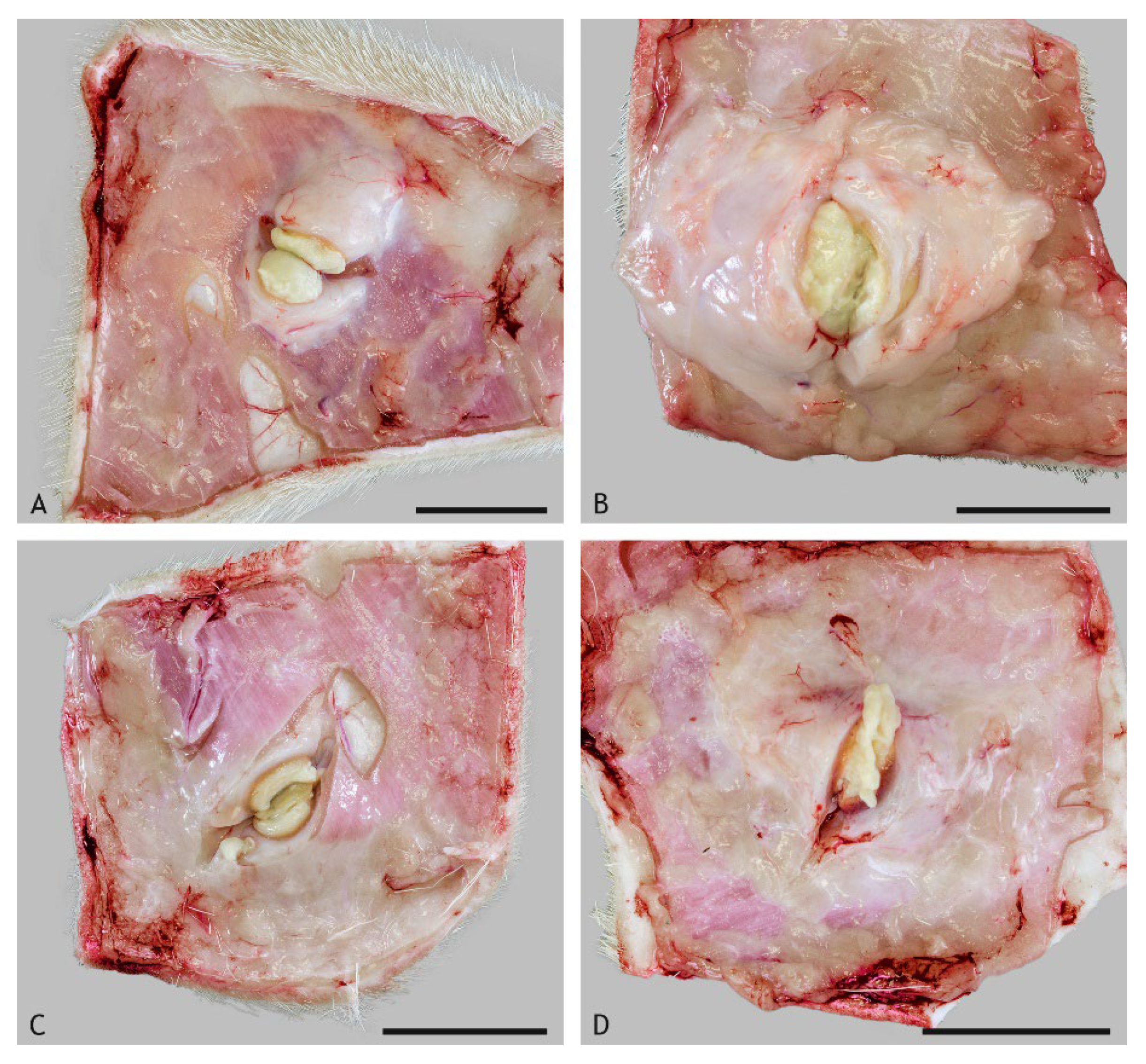
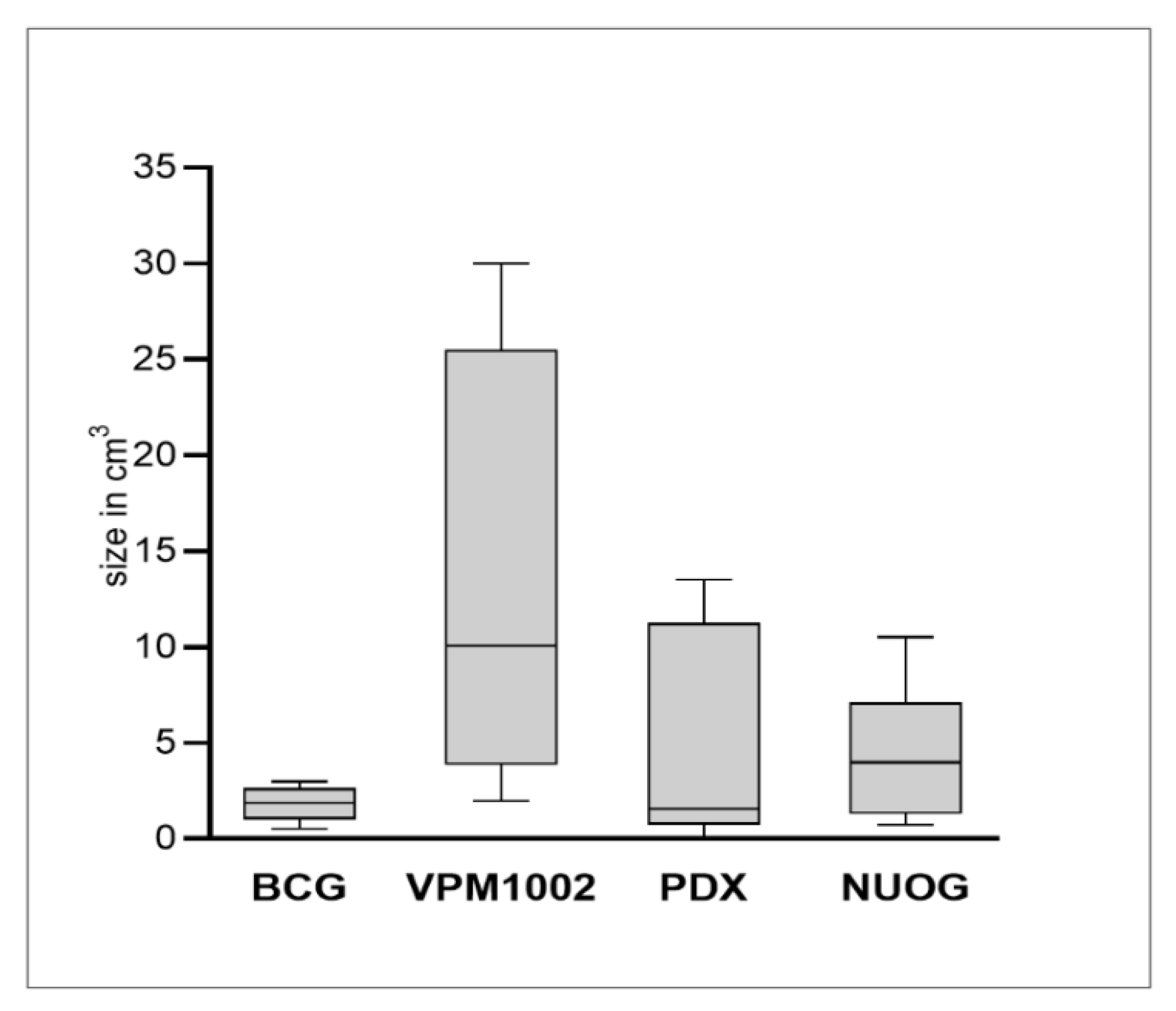
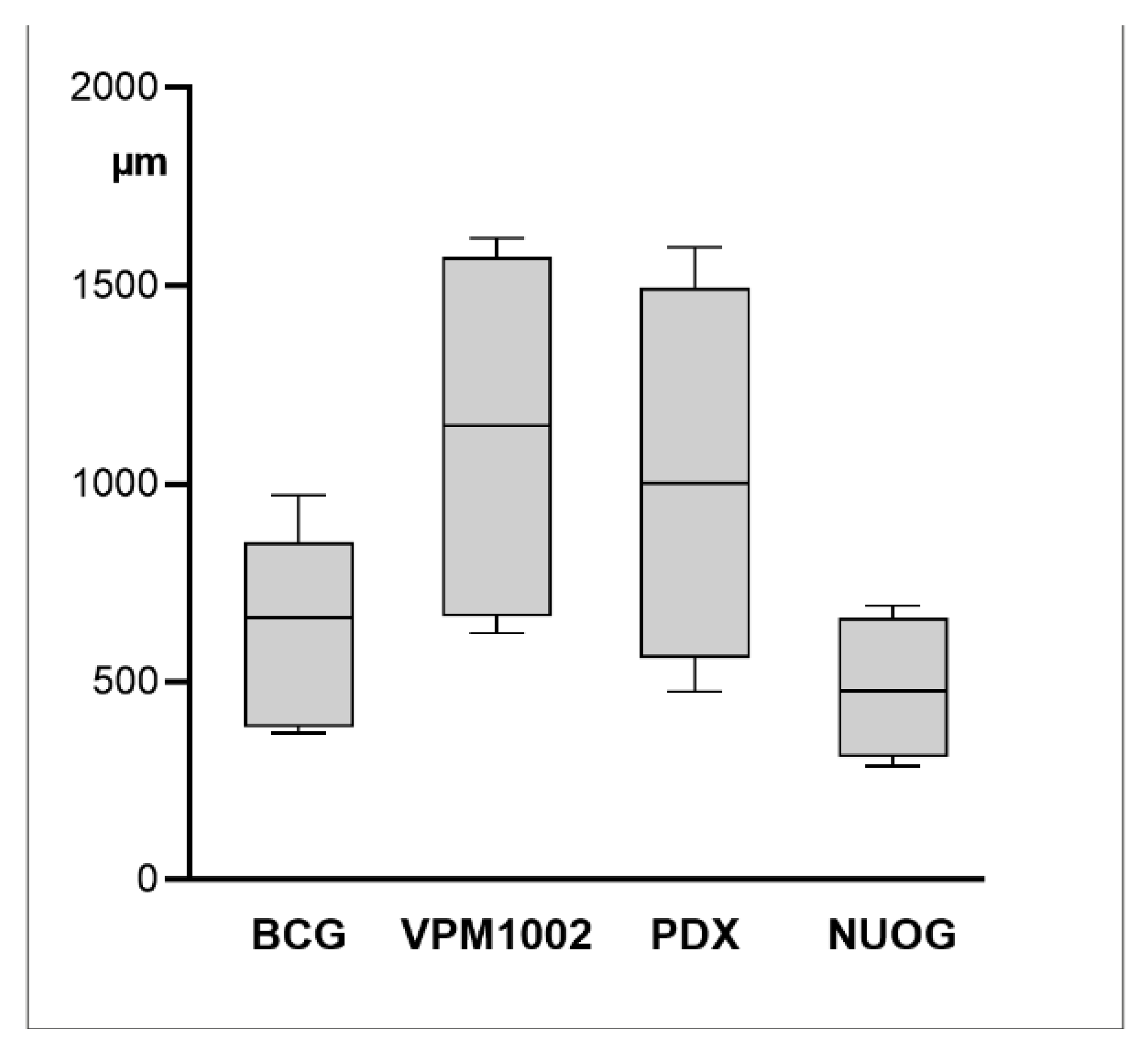
2.1. Macroscopic Findings
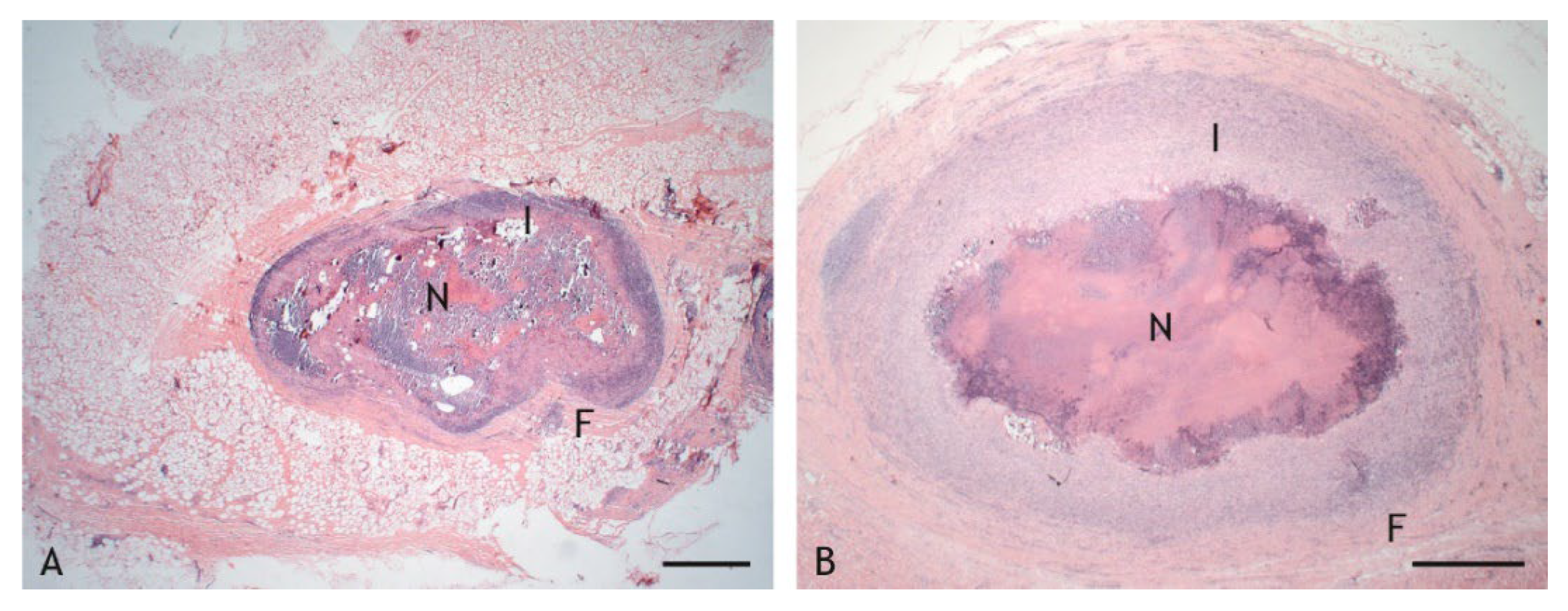
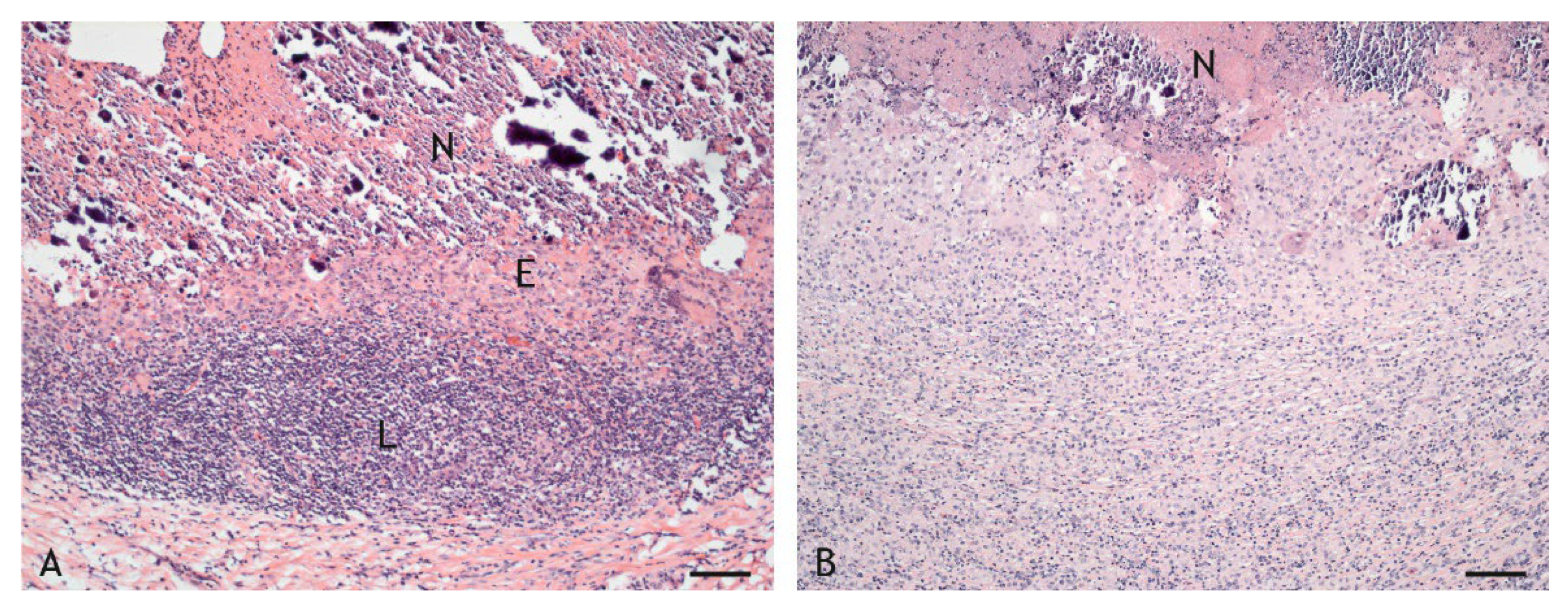
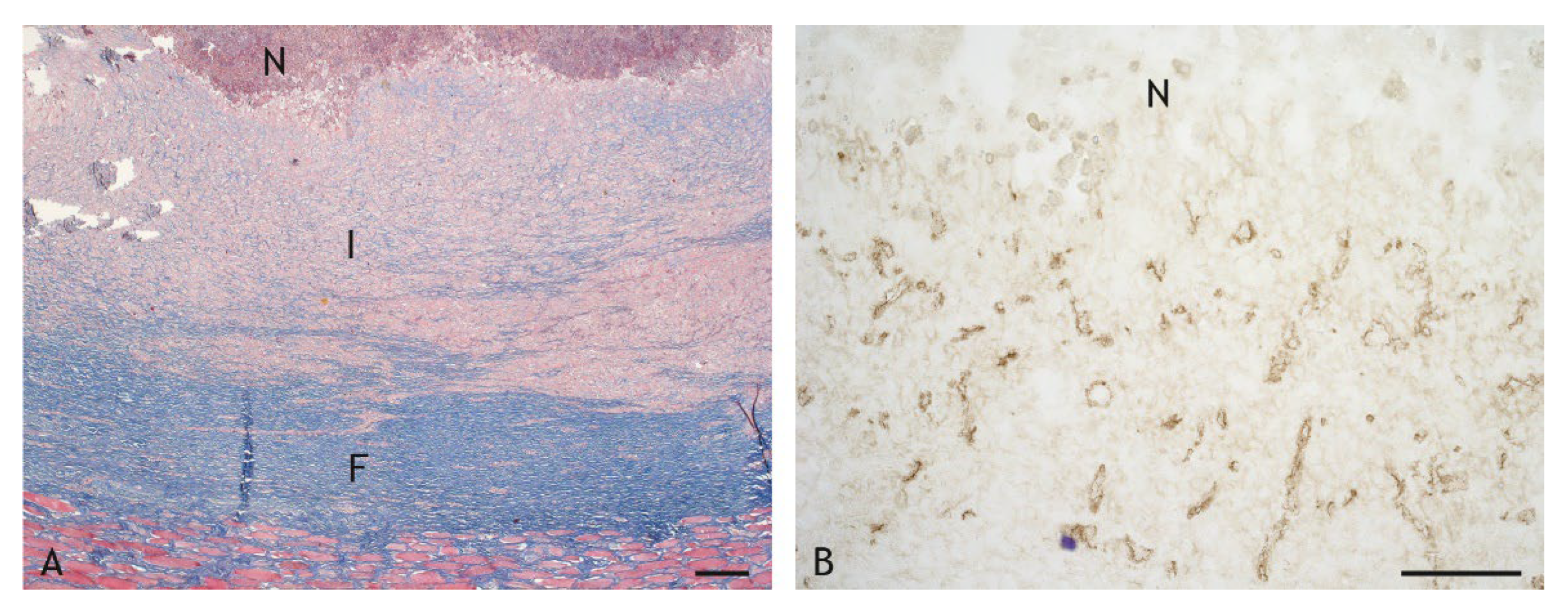
2.2. Morphologic Characteristics and Organization of Granulomas
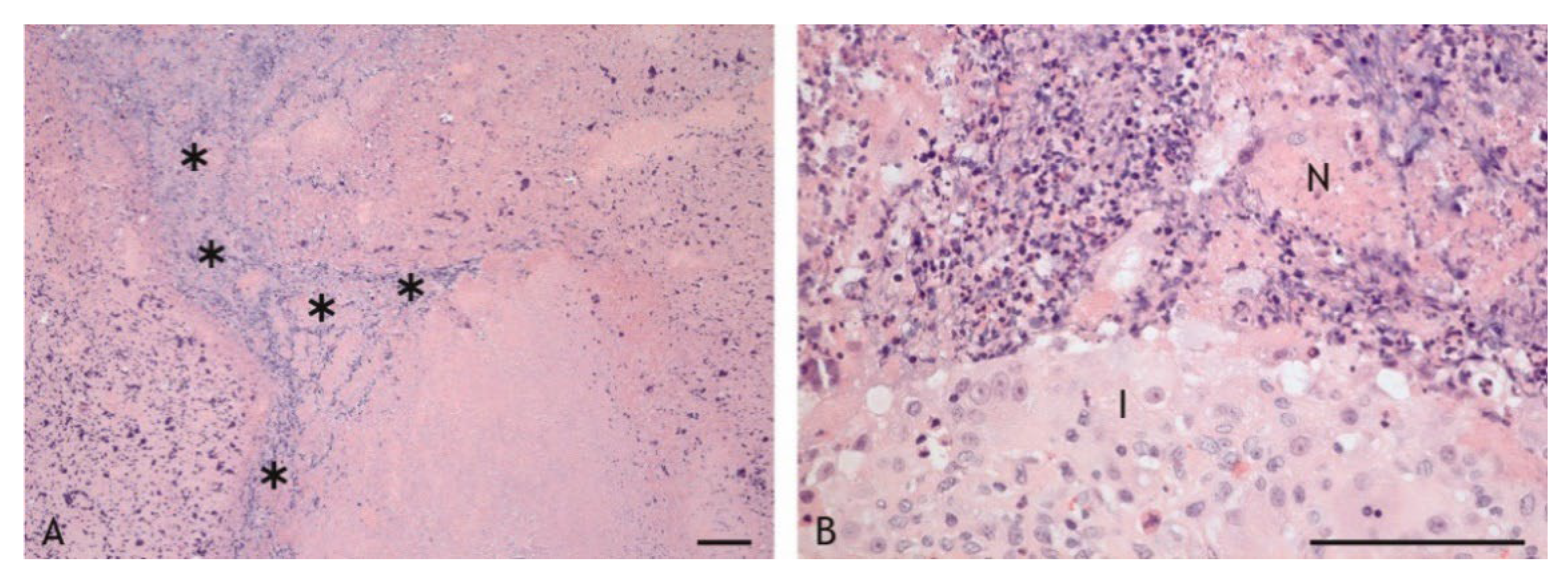
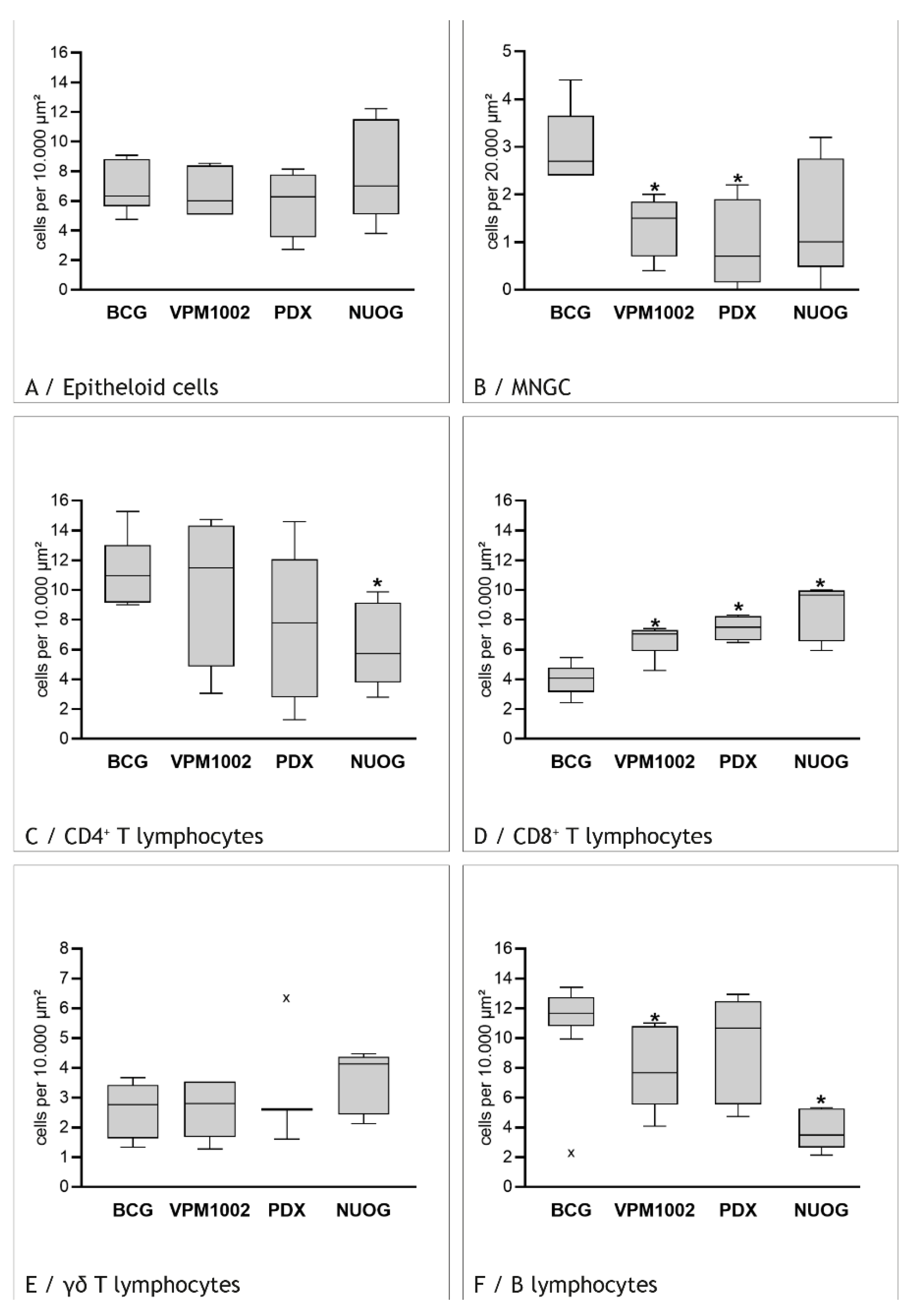
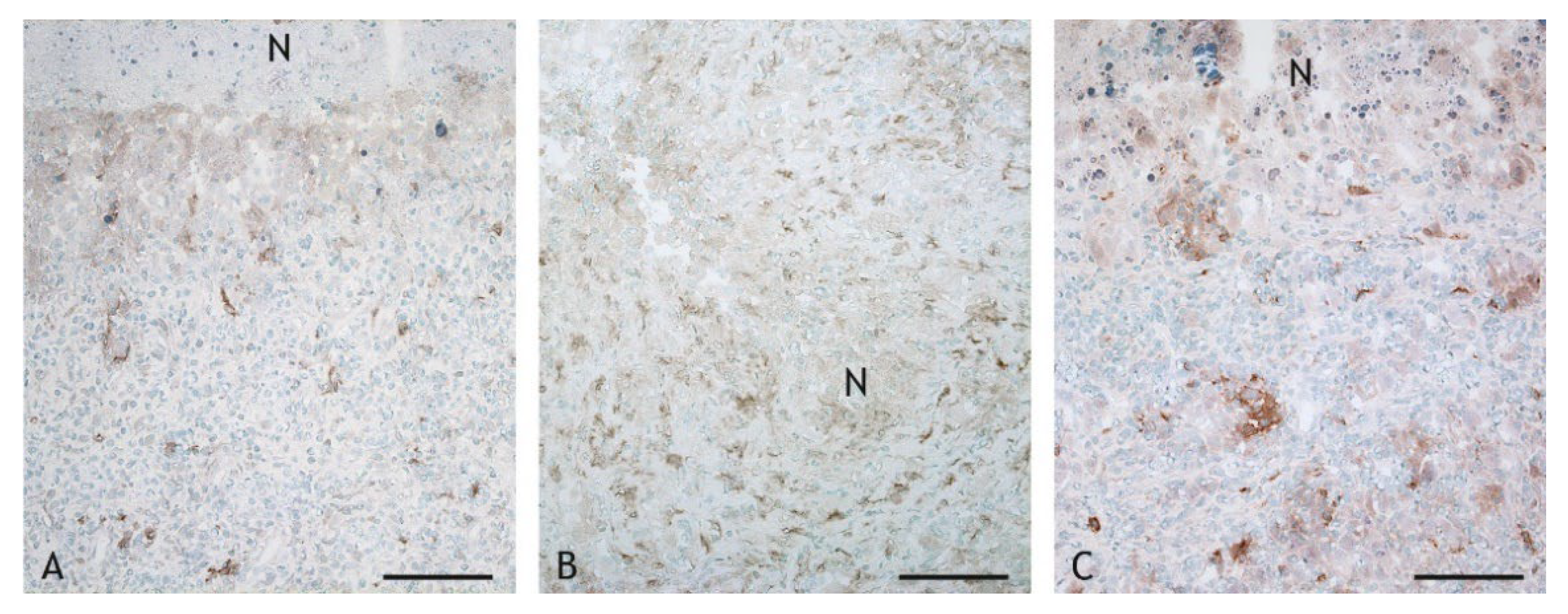
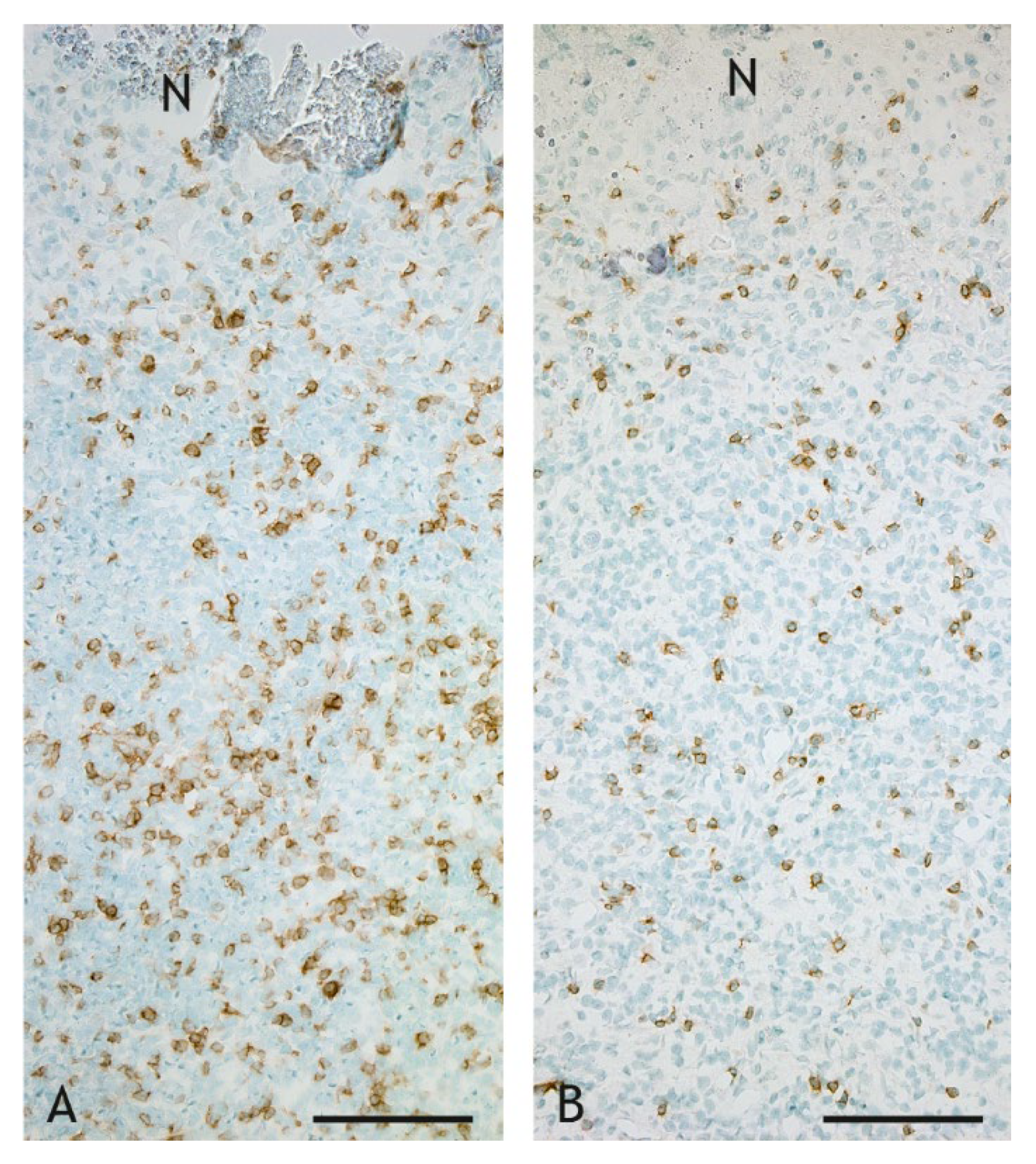
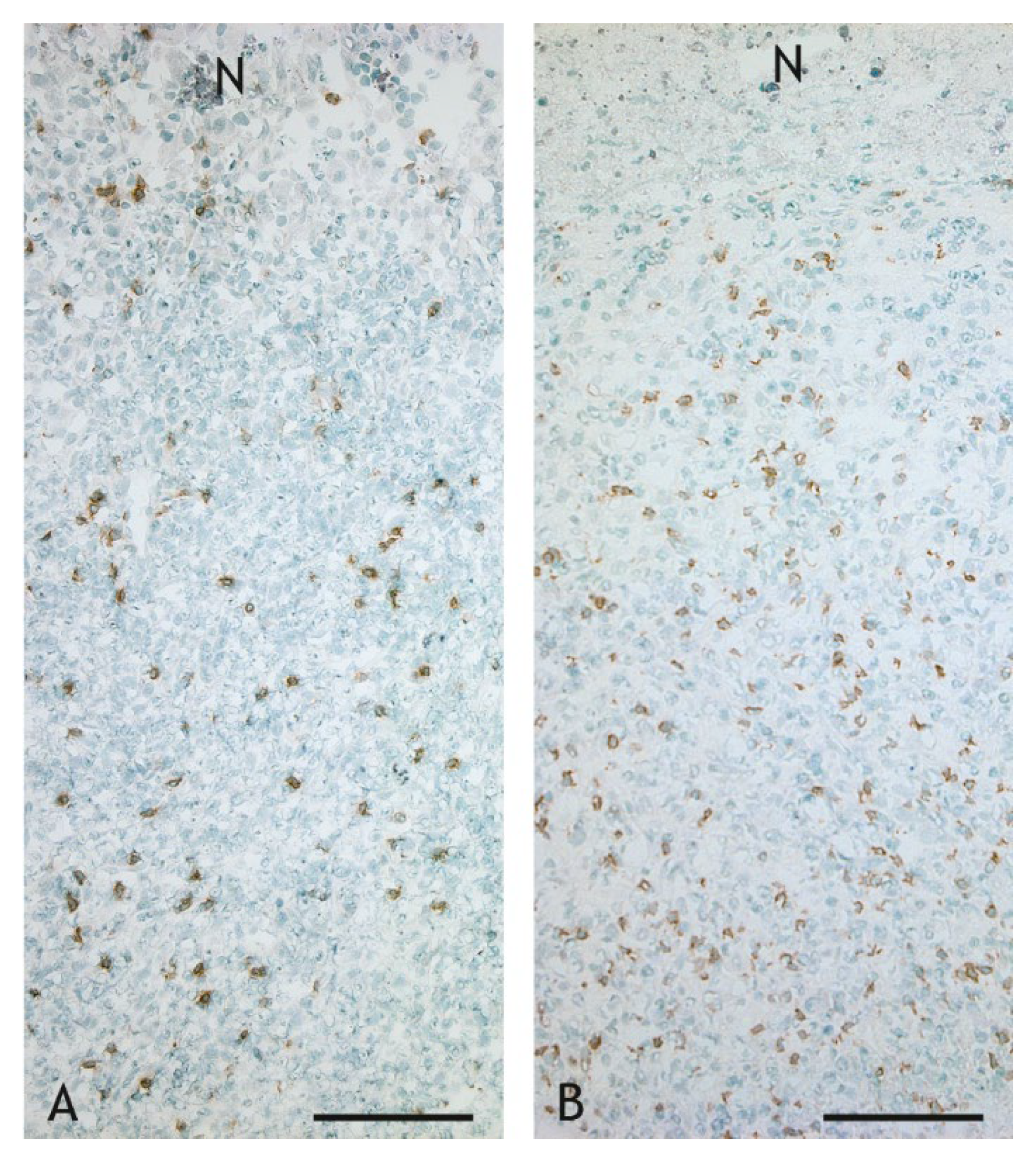
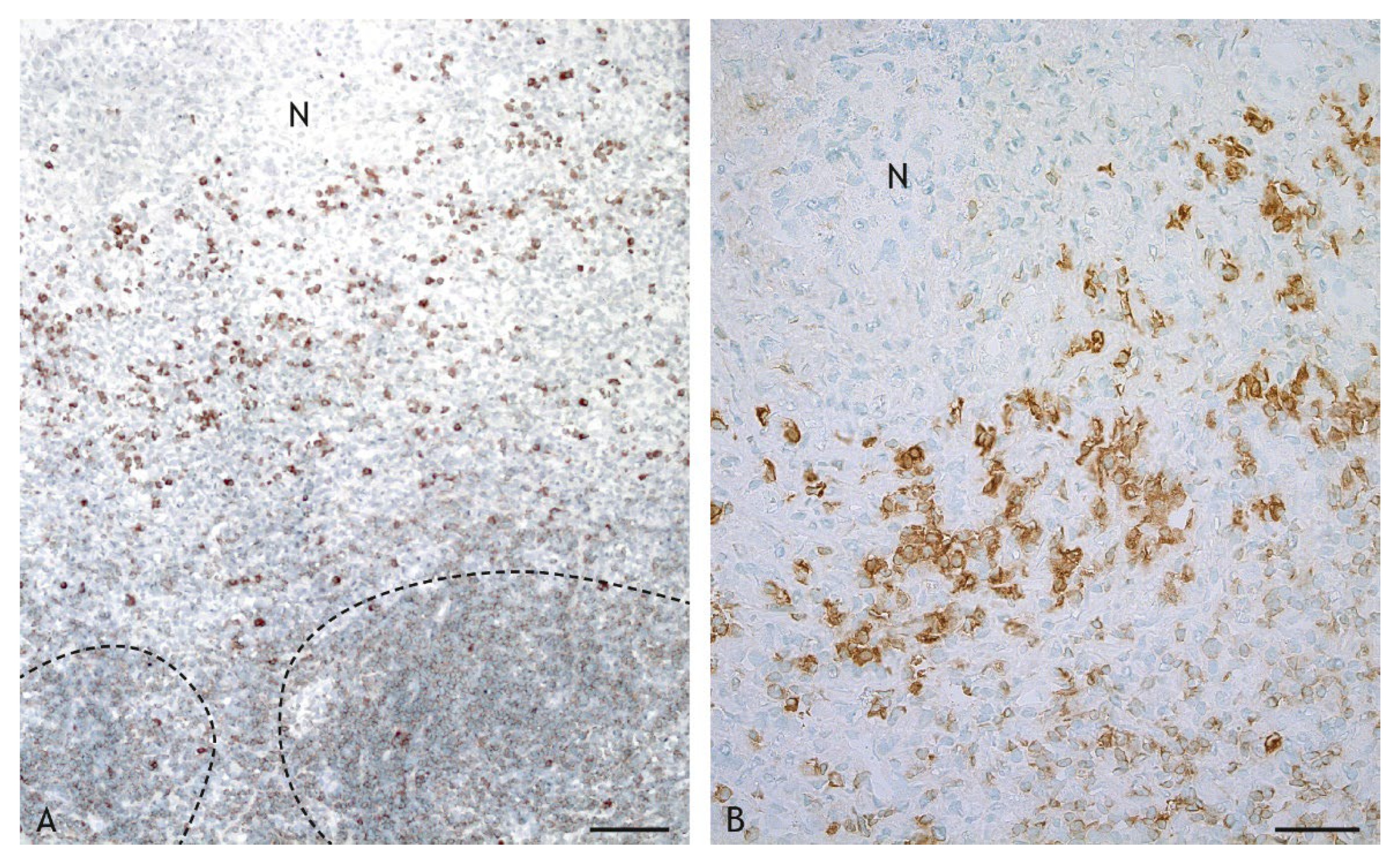
2.3. Cell Types in the Inflammatory Infiltrate
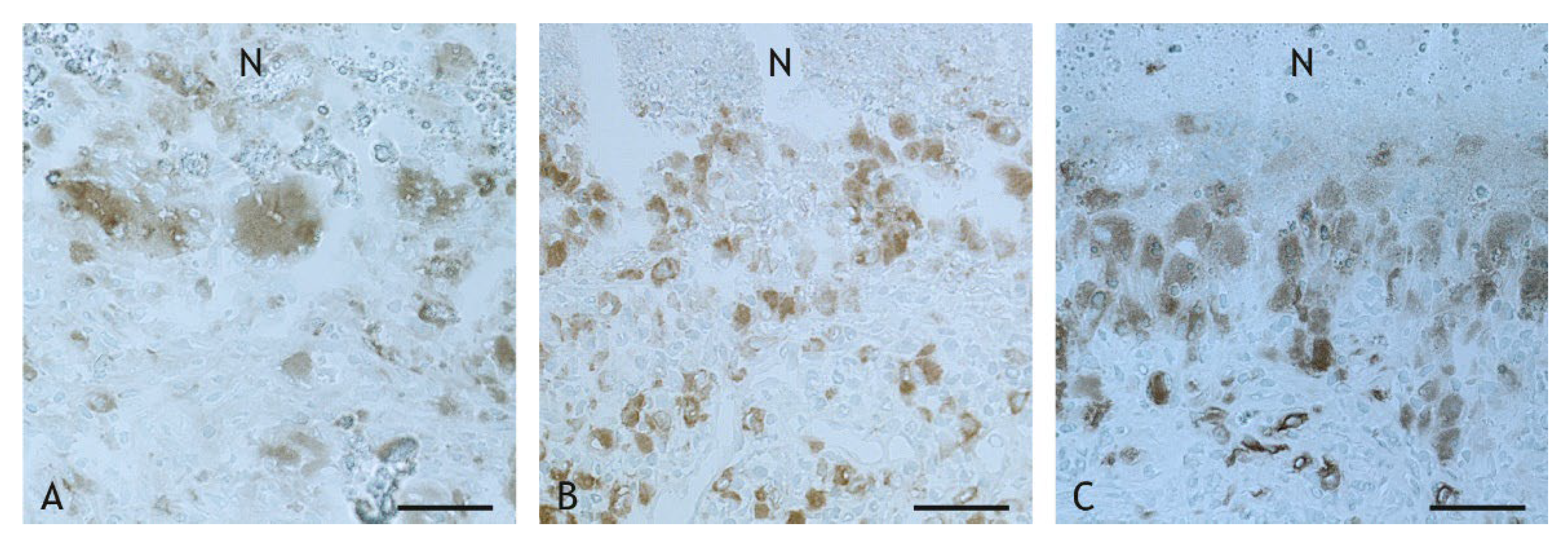
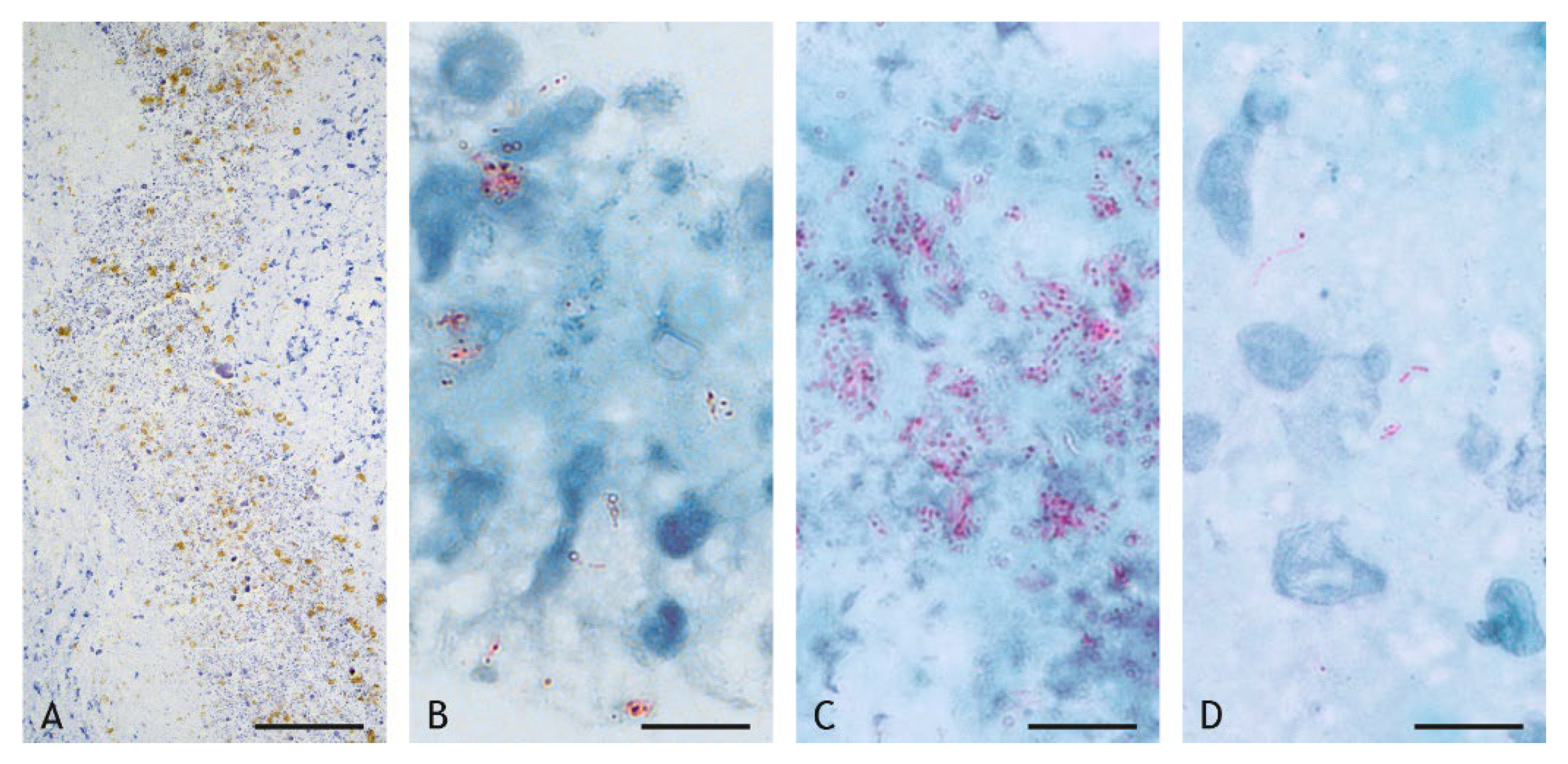
2.4. Mycobacteria in the Granuloma
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Vaccination
4.3. Clinical Examination and Findings after Vaccination
4.4. Necropsy, Gross Pathology and Tissue Samples
4.5. Histology
4.6. Immunohistochemistry
4.7. Bacterial Culture
4.8. Statistical Analysis
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Global Tuberculosis Report 2021; World Health Organization: Geneva, Switzerland, 2021. [Google Scholar]
- Jeremiah, C.; Petersen, E.; Nantanda, R.; Mungai, B.N.; Migliori, G.B.; Amanullah, F.; Lungu, P.; Ntoumi, F.; Kumarasamy, N.; Maeurer, M.; et al. The WHO Global Tuberculosis 2021 Report-not so good news and turning the tide back to end TB. Int. J. Infect. Dis. 2022, in press. [CrossRef]
- World Organization for Animal Health. Chapter 3.4.6. Bovine Tuberculosis. In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals 2018: OIE Terrestrial Manual 2018; World Organization for Animal Health: Paris, France, 2018; Available online: http://www.woah.org/fileadmin/Home/eng/Health_standards/tahm/3.04.06_BOVINE_TB.pdf (accessed on 25 July 2022).
- Domingo, M.; Vidal, E.; Marco, A. Pathology of bovine tuberculosis. Res. Vet. Sci. 2014, 97, S20–S29. [Google Scholar] [CrossRef] [PubMed]
- Pesciaroli, M.; Alvarez, J.; Boniotti, M.B.; Cagiola, M.; Di Marco, V.; Marianelli, C.; Pacciarini, M.; Pasquali, P. Tuberculosis in domestic animal species. Res. Vet. Sci. 2014, 97, S78–S85. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, S.H.E. The tuberculosis vaccine development pipeline: Present and furture priorities and challenges for research and innovation. In Essential Tuberculosis; Migliori, G.B., Raviglione, M.C., Eds.; Springer Nature Switzerland AG: Cham, Switzerland, 2021; pp. 395–405. [Google Scholar] [CrossRef]
- Lange, C.; Aaby, P.; Behr, M.A.; Donald, P.R.; Kaufmann, S.H.E.; Netea, M.G.; Mandalakas, A.M. 100 years of Mycobacterium bovis bacille Calmette-Guérin. Lancet Infect. Dis. 2022, 22, e2–e12. [Google Scholar] [CrossRef]
- Cobelens, F.; Suri, R.K.; Helinski, M.; Makanga, M.; Weinberg, A.L.; Schaffmeister, B.; Deege, F.; Hatherill, M. TB vaccine roadmap stakeholder group. Accelerating research and development of new vaccines against tuberculosis: A global roadmap. Lancet Infect. Dis. 2022, 22, e108–e120. [Google Scholar] [CrossRef]
- Puissegur, M.P.; Botanch, C.; Duteyrat, J.L.; Delsol, G.; Caratero, C.; Altare, F. An in vitro dual model of mycobacterial granulomas to investigate the molecular interactions between mycobacteria and human host cells. Cell. Microbiol. 2004, 6, 423–433. [Google Scholar] [CrossRef]
- Ray, J.C.; Flynn, J.L.; Kirschner, D.E. Synergy between individual TNF-dependent functions determines granuloma performance for controlling Mycobacterium tuberculosis infection. J. Immunol. 2009, 182, 3706–3717. [Google Scholar] [CrossRef]
- Fallahi-Sichani, M.; El-Kebir, M.; Marino, S.; Kirschner, D.E.; Linderman, J.J. Multiscale computational modeling reveals a critical role for TNF-α receptor 1 dynamics in tuberculosis granuloma formation. J. Immunol. 2011, 186, 3472–3483. [Google Scholar] [CrossRef]
- Ramakrishnan, L. Revisiting the role of the granuloma in tuberculosis. Nat. Rev. Immunol. 2012, 12, 352–366. [Google Scholar] [CrossRef]
- Oksanen, K.E.; Halfpenny, N.J.; Sherwood, E.; Harjula, S.K.; Hammarén, M.M.; Ahava, M.J.; Pajula, E.T.; Lahtinen, M.J.; Parikka, M.; Rämet, M. An adult zebrafish model for preclinical tuberculosis vaccine development. Vaccine 2013, 31, 5202–5209. [Google Scholar] [CrossRef]
- Garza-Cuartero, L.; McCarthy, E.; Brady, J.; Cassidy, J.; Hamilton, C.; Sekiya, M.; NcNair, J.; Mulcahy, G. Development of an in vitro model of the early-stage bovine tuberculous granuloma using Mycobacterium bovis-BCG. Vet. Immunol. Immunopathol. 2015, 168, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Ulrichs, T.; Kaufmann, S.H. New insights into the function of granulomas in human tuberculosis. J. Pathol. 2006, 208, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Palmer, M.V.; Kanipe, C.; Boggiatto, P.M. The bovine tuberculoid granuloma. Pathogens 2022, 11, 61. [Google Scholar] [CrossRef]
- Cardona, P.J.; Williams, A. Experimental animal modelling for TB vaccine development. Int. J. Infect. Dis. 2017, 56, 268–273. [Google Scholar] [CrossRef] [PubMed]
- Bellet, J.S.; Prose, N.S. Skin complications of Bacillus Calmette-Guérin immunization. Curr. Opin. Infect. Dis. 2005, 18, 97–100. [Google Scholar] [CrossRef] [PubMed]
- Dommergues, M.A.; de La Rocque, F.; Guy, C.; Lécuyer, A.; Jacquet, A.; Guérin, N.; Fagot, J.P.; Boucherat, M.; d’Athis, P.; Cohen, R. Local and regional adverse reactions to BCG-SSI vaccination: A 12-month cohort follow-up study. Vaccine 2009, 27, 6967–6973. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Seo, K.J.; Kim, J.J. Chest wall granuloma associated with BCG vaccination presenting as hot abscess in an immunocompetent infant. J. Cardiothorac. Surg. 2015, 10, 29. [Google Scholar] [CrossRef]
- Kigawa, T.; Kawai, M.; Yamaoka, H.; Kondoh, A.; Tojo, M.; Mabuchi, T. Two cases of subcutaneous tuberculous granuloma associated with BCG vaccination. Tokai J. Exp. Clin. Med. 2020, 45, 136–138. [Google Scholar]
- Valheim, M.; Hasvold, H.J.; Storset, A.K.; Larsen, H.J.; Press, C.M. Localisation of CD25+ cells and MHCII+ cells in lymph nodes draining Mycobacterium avium subsp. paratuberculosis vaccination granuloma and the presence of a systemic immune response. Res. Vet. Sci. 2002, 73, 77–85. [Google Scholar] [CrossRef]
- Simutis, F.J.; Cheville, N.F.; Jones, D.E. Investigation of antigen-specific T-cell responses and subcutaneous granuloma development during experimental sensitization of calves with Mycobacterium avium subsp. paratuberculosis. Am. J. Vet. Res. 2005, 66, 474–482. [Google Scholar] [CrossRef]
- Eppleston, J.; Windsor, P.A. Lesions attributed to vaccination of sheep with Gudair for the control of ovine paratuberculosis: Post farm economic impacts at slaughter. Aust. Vet. J. 2007, 85, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Palmer, M.V.; Thacker, T.C.; Waters, W.R. Vaccination with Mycobacterium bovis BCG strains Danish and Pasteur in white-tailed deer (Odocoileus virginianus) experimentally challenged with Mycobacterium bovis. Zoonoses Public Health 2009, 56, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Pérez de Val, B.; Vidal, E.; López-Soria, S.; Marco, A.; Cervera, Z.; Martín, M.; Mercader, I.; Singh, M.; Raeber, A.; Domingo, M. Assessment of safety and interferon gamma responses of Mycobacterium bovis BCG vaccine in goat kids and milking goats. Vaccine 2016, 34, 881–886. [Google Scholar] [CrossRef] [PubMed]
- Figl, J.; Köhler, H.; Wedlich, N.; Liebler-Tenorio, E.M.; Grode, L.; Parzmair, G.; Krishnamoorthy, G.; Nieuwenhiuzen, N.; Kaufmann, S.H.E.; Menge, C. Safety and immunogenicity of recombinant Bacille Calmette-Guérin strain VPM1002 and its derivatives in a goat model. Int. J. Mol. Sci. 2022, submitted.
- Grode, L.; Seiler, P.; Baumann, S.; Hess, J.; Brinkmann, V.; Nasser Eddine, A.; Mann, P.; Goosmann, C.; Bandermann, S.; Smith, D.; et al. Increased vaccine efficacy against tuberculosis of recombinant Mycobacterium bovis bacille Calmette-Guérin mutants that secrete listeriolysin. J. Clin. Investig. 2005, 115, 2472–2479. [Google Scholar] [CrossRef]
- Nieuwenhuizen, N.E.; Kulkarni, P.S.; Shaligram, U.; Cotton, M.F.; Rentsch, C.A.; Eisele, B.; Grode, L.; Kaufmann, S.H.E. The recombinant Bacille Calmette-Guérin vaccine VPM1002: Ready for clinical efficacy testing. Front. Immunol. 2017, 8, 1147. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, S.H.E. Vaccination against tuberculosis: Revamping BCG by molecular genetics guided by immunology. Front. Immunol. 2020, 11, 316. [Google Scholar] [CrossRef]
- Kaufmann, S.H.E. Vaccine development against tuberculosis over the last 140 years: Failure as part of success. Front. Microbiol. 2021, 12, 750124. [Google Scholar] [CrossRef]
- Winau, F.; Weber, S.; Sad, S.; de Diego, J.; Locatelli Hoops, S.; Breiden, B.; Sandhoff, K.; Brinkmann, V.; Kaufmann, S.H.E.; Schaible, U.E. Apoptotic vesicles crossprime CD8 T cells and protect against tuberculosis. Immunity 2006, 24, 105–117. [Google Scholar] [CrossRef]
- Saiga, H.; Nieuwenhuizen, N.; Gengenbacher, M.; Koehler, A.B.; Schuerer, S.; Moura-Alves, P.; Wagner, I.; Mollenkopf, H.J.; Dorhoi, A.; Kaufmann, S.H. The recombinant BCG ΔureC::hly vaccine targets the AIM2 inflammasome to induce autophagy and inflammation. J. Infect. Dis. 2015, 211, 1831–1841. [Google Scholar] [CrossRef]
- Desel, C.; Dorhoi, A.; Bandermann, S.; Grode, L.; Eisele, B.; Kaufmann, S.H. Recombinant BCG ΔureC hly+ induces superior protection over parental BCG by stimulating a balanced combination of type 1 and type 17 cytokine responses. J. Infect. Dis. 2011, 204, 1573–1584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farinacci, M.; Weber, S.; Kaufmann, S.H. The recombinant tuberculosis vaccine rBCG ΔureC::hly(+) induces apoptotic vesicles for improved priming of CD4(+) and CD8(+) T cells. Vaccine 2012, 30, 7608–7614. [Google Scholar] [CrossRef] [PubMed]
- Vogelzang, A.; Perdomo, C.; Zedler, U.; Kuhlmann, S.; Hurwitz, R.; Gengenbacher, M.; Kaufmann, S.H. Central memory CD4+ T cells are responsible for the recombinant Bacillus Calmette-Guérin ΔureC::hly vaccine’s superior protection against tuberculosis. J. Infect. Dis. 2014, 210, 1928–1937. [Google Scholar] [CrossRef]
- Grode, L.; Ganoza, C.A.; Brohm, C.; Weiner 3rd, J.; Eisele, B.; Kaufmann, S.H.E. Safety and immunogenicity of the recombinant BCG vaccine VPM1002 against tuberculosis in a phase I open-label randomized clinical trial. Vaccine 2013, 31, 1340–1348. [Google Scholar] [CrossRef] [PubMed]
- Loxton, A.G.; Knaul, J.K.; Grode, L.; Gutschmidt, A.; Meller, C.; Eisele, B.; Johnstone, H.; van der Spuy, G.; Maertzdorf, J.; Kaufmann, S.H.E.; et al. Safety and immunogenicity of the recombinant Mycobacterium bovis BCG Vaccine VPM1002 in HIV-unexposed newborn infants in South Africa. Clin. Vaccine Immunol. 2017, 24, e00439-16. [Google Scholar] [CrossRef] [PubMed]
- Cotton, M.F.; Madhi, S.A.; Luabeya, A.K.; Tameris, M.; Hesseling, A.C.; Shenje, J.; Schoeman, E.; Hatherill, M.; Desai, S.; Kapse, D.; et al. Safety and immunogenicity of VPM1002 versus BCG in South African newborn babies: A randomised, phase 2 non-inferiority double-blind controlled trial. Lancet Infect. Dis. 2022, in press. [Google Scholar] [CrossRef]
- Gengenbacher, M.; Vogelzang, A.; Schuerer, S.; Lazar, D.; Kaiser, P.; Kaufmann, S.H. Dietary pyridoxine controls efficacy of vitamin B6-auxotrophic tuberculosis vaccine bacillus Calmette-Guérin ΔureC::hly Δpdx1 in mice. mBio 2014, 5, e01262-14. [Google Scholar] [CrossRef]
- Gengenbacher, M.; Nieuwenhuizen, N.; Vogelzang, A.; Liu, H.; Kaiser, P.; Schuerer, S.; Lazar, D.; Wagner, I.; Mollenkopf, H.J.; Kaufmann, S.H. Deletion of nuoG from the vaccine candidate Mycobacterium bovis BCG ΔureC::hly improves protection against tuberculosis. mBio 2016, 7, e00679-16. [Google Scholar] [CrossRef]
- de Val Pérez, B.; López-Soria, S.; Nofrarías, M.; Martín, M.; Vordermeier, H.M.; Villarreal-Ramos, B.; Romera, N.; Escobar, M.; Solanes, D.; Cardona, P.J.; et al. Experimental model of tuberculosis in the domestic goat after endobronchial infection with Mycobacterium caprae. Clin. Vaccine Immunol. 2011, 18, 1872–1881. [Google Scholar] [CrossRef]
- Bezos, J.; Casal, C.; Díez-Delgado, I.; Romero, B.; Liandris, E.; Álvarez, J.; Sevilla, I.A.; Juan, L.; Domínguez, L.; Gortázar, C. Goats challenged with different members of the Mycobacterium tuberculosis complex display different clinical pictures. Vet. Immunol. Immunopathol. 2015, 167, 185–189. [Google Scholar] [CrossRef]
- Köhler, H.; Soschinka, A.; Meyer, M.; Kather, A.; Reinhold, P.; Liebler-Tenorio, E. Characterization of a caprine model for the subclinical initial phase of Mycobacterium avium subsp. paratuberculosis infection. BMC Vet. Res. 2015, 11, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schinköthe, J.; Köhler, H.; Liebler-Tenorio, E.M. Characterization of tuberculous granulomas in different stages of progression and associated tertiary lymphoid tissue in goats experimentally infected with Mycobacterium avium subsp. hominissuis. Comp. Immunol. Microbiol. Infect. Dis. 2016, 47, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Schinköthe, J.; Möbius, P.; Köhler, H.; Liebler-Tenorio, E.M. Experimental infection of goats with Mycobacterium avium subsp. hominissuis: A model for comparative tuberculosis research. J. Comp. Pathol. 2016, 155, 218–230. [Google Scholar] [CrossRef] [PubMed]
- Wangoo, A.; Johnson, L.; Gough, J.; Ackbar, R.; Inglut, S.; Hicks, D.; Spencer, Y.; Hewinson, G.; Vordermeier, M. Advanced granulomatous lesions in Mycobacterium bovis-infected cattle are associated with increased expression of type I procollagen, gammadelta (WC1+) T cells and CD 68+ cells. J. Comp. Pathol. 2005, 133, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Popescu, M.R.; Călin, G.; Strâmbu, I.; Olaru, M.; Bălăşoiu, M.; Huplea, V.; Zdrancotă, C.; Pleşea, R.M.; Enache, S.D.; Pleşea, I.E. Lymph node tuberculosis-an attempt of clinico-morphological study and review of the literature. Rom. J. Morphol. Embryol. 2014, 55 (Suppl. S2), 553–567. [Google Scholar] [PubMed]
- Ozturk-Gurgen, H.; Rieseberg, B.; Leipig-Rudolph, M.; Straubinger, R.K.; Hermanns, W. Morphology of naturally-occurring tuberculosis in cattle caused by Mycobacterium caprae. J. Comp. Pathol. 2020, 174, 120–139. [Google Scholar] [CrossRef] [PubMed]
- Palmer, M.V.; Thacker, T.C.; Kanipe, C.; Boggiatto, P.M. Heterogeneity of pulmonary granulomas in cattle experimentally infected with Mycobacterium bovis. Front. Vet. Sci. 2021, 8, 671460. [Google Scholar] [CrossRef] [PubMed]
- Palmer, M.V.; Waters, W.R.; Thacker, T.C. Lesion development and immunohistochemical changes in granulomas from cattle experimentally infected with Mycobacterium bovis. Vet. Pathol. 2007, 44, 863–874. [Google Scholar] [CrossRef]
- Reinhold, P.; Köhler, K.; Liebler-Tenorio, E. Versuchstierkunde bei Nutztieren: Einblicke in Modelle für respiratorische Erkrankungen und Infektionen. Berl. Münchn. Tierärztl. Wschr. 2018, 131, 417–443. [Google Scholar] [CrossRef]
- Silva Miranda, M.; Breiman, A.; Allain, S.; Deknuydt, F.; Altare, F. The tuberculous granuloma: An unsuccessful host defense mechanism providing a safety shelter for the bacteria? Clin. Dev. Immunol. 2012, 2012, 139127. [Google Scholar] [CrossRef]
- Martinot, A.J. Microbial offense vs host defense: Who controls the TB granuloma? Vet. Pathol. 2018, 55, 14–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velmurugan, K.; Chen, B.; Miller, J.L.; Azogue, S.; Gurses, S.; Hsu, T.; Glickman, M.; Jacobs, W.R., Jr.; Porcelli, S.A.; Briken, V. Mycobacterium tuberculosis nuoG is a virulence gene that inhibits apoptosis of infected host cells. PLoS Pathog. 2007, 3, e110. [Google Scholar] [CrossRef] [PubMed]
- Schaible, U.E.; Winau, F.; Sieling, P.A.; Fischer, K.; Collins, H.L.; Hagens, K.; Modlin, R.L.; Brinkmann, V.; Kaufmann, S.H. Apoptosis facilitates antigen presentation to T lymphocytes through MHC-I and CD1 in tuberculosis. Nat. Med. 2003, 9, 1039–1046. [Google Scholar] [CrossRef]
- Peyron, P.; Vaubourgeix, J.; Poquet, Y.; Levillain, F.; Botanch, C.; Bardou, F.; Daffé, M.; Emile, J.F.; Marchou, B.; Cardona, P.J.; et al. Foamy macrophages from tuberculous patients’ granulomas constitute a nutrient-rich reservoir for M. tuberculosis persistence. PLoS Pathog. 2008, 4, e1000204. [Google Scholar] [CrossRef]
- Ehlers, S.; Schaible, U.E. The granuloma in tuberculosis: Dynamics of a host-pathogen collusion. Front. Immunol. 2013, 3, 411. [Google Scholar] [CrossRef] [PubMed]
- Cronan, M.R.; Beerman, R.W.; Rosenberg, A.F.; Saelens, J.W.; Johnson, M.G.; Oehlers, S.H.; Sisk, D.M.; Jurcic Smith, K.L.; Medvitz, N.A.; Miller, S.E.; et al. Macrophage epithelial reprogramming underlies mycobacterial granuloma formation and promotes infection. Immunity 2016, 45, 861–876. [Google Scholar] [CrossRef] [PubMed]
- Lenaerts, A.J.; Hoff, D.; Aly, S.; Ehlers, S.; Andries, K.; Cantarero, L.; Orme, I.M.; Basaraba, R.J. Location of persisting mycobacteria in a Guinea pig model of tuberculosis revealed by r207910. Antimicrob. Agents Chemother. 2007, 51, 3338–3345. [Google Scholar] [CrossRef]
- Russell, D.G.; Cardona, P.J.; Kim, M.J.; Allain, S.; Altare, F. Foamy macrophages and the progression of the human tuberculosis granuloma. Nat. Immunol. 2009, 10, 943–948. [Google Scholar] [CrossRef]
- Kim, M.J.; Wainwright, H.C.; Locketz, M.; Bekker, L.G.; Walther, G.B.; Dittrich, C.; Visser, A.; Wang, W.; Hsu, F.F.; Wiehart, U.; et al. Caseation of human tuberculosis granulomas correlates with elevated host lipid metabolism. EMBO Mol. Med. 2010, 2, 258–274. [Google Scholar] [CrossRef]
- Borkute, R.R.; Woelke, S.; Pei, G.; Dorhoi, A. Neutrophils in tuberculosis: Cell biology, cellular networking and multitasking in host defense. Int. J. Mol. Sci. 2021, 22, 4801. [Google Scholar] [CrossRef]
- Korbel, D.S.; Schneider, B.E.; Schaible, U.E. Innate immunity in tuberculosis: Myths and truth. Microbes Infect. 2008, 10, 995–1004. [Google Scholar] [CrossRef] [PubMed]
- Flynn, J.L.; Chan, J.; Lin, P.L. Macrophages and control of granulomatous inflammation in tuberculosis. Mucosal Immunol. 2011, 4, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Wassermann, R.; Gulen, M.F.; Sala, C.; Perin, S.G.; Lou, Y.; Rybniker, J.; Schmid-Burgk, J.L.; Schmidt, T.; Hornung, V.; Cole, S.T.; et al. Mycobacterium tuberculosis differentially activates cGAS- and inflammasome-dependent intracellular immune responses through ESX-1. Cell Host Microbe 2015, 17, 799–810. [Google Scholar] [CrossRef] [PubMed]
- Queval, C.J.; Brosch, R.; Simeone, R. The macrophage: A disputed fortress in the battle against Mycobacterium tuberculosis. Front. Microbiol. 2017, 8, 2284. [Google Scholar] [CrossRef]
- van der Wel, N.; Hava, D.; Houben, D.; Fluitsma, D.; van Zon, M.; Pierson, J.; Brenner, M.; Peters, P.J. M. tuberculosis and M. leprae translocate from the phagolysosome to the cytosol in myeloid cells. Cell 2007, 129, 1287–1298. [Google Scholar] [CrossRef]
- Brodin, P.; Majlessi, L.; Marsollier, L.; de Jonge, M.I.; Bottai, D.; Demangel, C.; Hinds, J.; Neyrolles, O.; Butcher, P.D.; Leclerc, C.; et al. Dissection of ESAT-6 system 1 of Mycobacterium tuberculosis and impact on immunogenicity and virulence. Infect. Immun. 2006, 74, 88–98. [Google Scholar] [CrossRef]
- Augenstreich, J.; Arbues, A.; Simeone, R.; Haanappel, E.; Wegener, A.; Sayes, F.; Le Chevalier, F.; Chalut, C.; Malaga, W.; Guilhot, C.; et al. ESX-1 and phthiocerol dimycocerosates of Mycobacterium tuberculosis act in concert to cause phagosomal rupture and host cell apoptosis. Cell. Microbiol. 2017, 19, e12726. [Google Scholar] [CrossRef]
- Simeone, R.; Sayes, F.; Lawarée, E.; Brosch, R. Breaching the phagosome, the case of the tuberculosis agent. Cell. Microbiol. 2021, 23, e13344. [Google Scholar] [CrossRef]
- Neill, S.D.; Pollock, J.M.; Bryson, D.B.; Hanna, J. Pathogenesis of Mycobacterium bovis infection in cattle. Vet. Microbiol. 1994, 40, 41–52. [Google Scholar] [CrossRef]
- Gharun, K.; Senges, J.; Seidl, M.; Lösslein, A.; Kolter, J.; Lohrmann, F.; Fliegauf, M.; Elgizouli, M.; Alber, M.; Vavra, M.; et al. Mycobacteria exploit nitric oxide-induced transformation of macrophages into permissive giant cells. EMBO Rep. 2017, 18, 2144–2159. [Google Scholar] [CrossRef]
- Brooks, P.J.; Glogauer, M.; McCulloch, C.A. An overview of the derivation and function of multinucleated giant cells and their role in pathologic processes. Am. J. Pathol. 2019, 189, 1145–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Queval, C.J.; Fearns, A.; Botella, L.; Smyth, A.; Schnettger, L.; Mitermite, M.; Wooff, E.; Villarreal-Ramos, B.; Garcia-Jimenez, W.; Heunis, T.; et al. Macrophage-specific responses to human- and animal-adapted tubercle bacilli reveal pathogen and host factors driving multinucleated cell formation. PLoS Pathog. 2021, 17, e1009410. [Google Scholar] [CrossRef] [PubMed]
- Lay, G.; Poquet, Y.; Salek-Peyron, P.; Puissegur, M.P.; Botanch, C.; Bon, H.; Levillain, F.; Duteyrat, J.L.; Emile, J.F.; Altare, F. Langhans giant cells from M. tuberculosis-induced human granulomas cannot mediate mycobacterial uptake. J. Pathol. 2007, 211, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Palmer, M.V.; Thacker, T.C.; Waters, W.R. Multinucleated giant cell cytokine expression in pulmonary granulomas of cattle experimentally infected with Mycobacterium bovis. Vet. Immunol. Immunopathol. 2016, 180, 34–39. [Google Scholar] [CrossRef]
- Hope, J.C.; Thom, M.L.; McCormick, P.A.; Howard, C.J. Interaction of antigen presenting cells with mycobacteria. Vet. Immunol. Immunopathol. 2004, 100, 187–195. [Google Scholar] [CrossRef]
- Philips, J.A.; Ernst, J.D. Tuberculosis pathogenesis and immunity. Annu. Rev. Pathol. 2012, 7, 353–384. [Google Scholar] [CrossRef]
- Tailleux, L.; Neyrolles, O.; Honoré-Bouakline, S.; Perret, E.; Sanchez, F.; Abastado, J.P.; Lagrange, P.H.; Gluckman, J.C.; Rosenzwajg, M.; Herrmann, J.L. Constrained intracellular survival of Mycobacterium tuberculosis in human dendritic cells. J. Immunol. 2003, 170, 1939–1948. [Google Scholar] [CrossRef] [PubMed]
- Lozza, L.; Farinacci, M.; Faé, K.; Bechtle, M.; Stäber, M.; Dorhoi, A.; Bauer, M.; Ganoza, C.; Weber, S.; Kaufmann, S.H. Crosstalk between human DC subsets promotes antibacterial activity and CD8+ T-cell stimulation in response to bacille Calmette-Guérin. Eur. J. Immunol. 2014, 44, 80–92. [Google Scholar] [CrossRef] [PubMed]
- Stenger, S.; Hanson, D.A.; Teitelbaum, R.; Dewan, P.; Niazi, K.R.; Froelich, C.J.; Ganz, T.; Thoma-Uszynski, S.; Melián, A.; Bogdan, C.; et al. An antimicrobial activity of cytolytic T cells mediated by granulysin. Science 1998, 282, 121–125. [Google Scholar] [CrossRef]
- Thoma-Uszynski, S.; Stenger, S.; Modlin, R.L. CTL-mediated killing of intracellular Mycobacterium tuberculosis is independent of target cell nuclear apoptosis. J. Immunol. 2000, 165, 5773–5779. [Google Scholar] [CrossRef]
- Woodworth, J.S.; Behar, S.M. Mycobacterium tuberculosis-specific CD8+ T cells and their role in immunity. Crit. Rev. Immunol. 2006, 26, 317–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hein, W.R.; Dudler, L. TCR gamma delta+ cells are prominent in normal bovine skin and express a diverse repertoire of antigen receptors. Immunology 1997, 91, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Yirsaw, A.; Baldwin, C.L. Goat γδ T cells. Dev. Comp. Immunol. 2021, 114, 103809. [Google Scholar] [CrossRef]
- McGill, J.L.; Sacco, R.E.; Baldwin, C.L.; Telfer, J.C.; Palmer, M.V.; Waters, W.R. The role of gamma delta T cells in immunity to Mycobacterium bovis infection in cattle. Vet. Immunol. Immunopathol. 2014, 159, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Falini, B.; Flenghi, L.; Pileri, S.; Pelicci, P.; Fagioli, M.; Martelli, M.F.; Moretta, L.; Ciccone, E. Distribution of T cells bearing different forms of the T cell receptor gamma/delta in normal and pathological human tissues. J. Immunol. 1989, 143, 2480–2488. [Google Scholar]
- Ulrichs, T.; Kosmiadi, G.A.; Trusov, V.; Jörg, S.; Pradl, L.; Titukhina, M.; Mishenko, V.; Gushina, N.; Kaufmann, S.H. Human tuberculous granulomas induce peripheral lymphoid follicle-like structures to orchestrate local host defence in the lung. J. Pathol. 2004, 204, 217–228. [Google Scholar] [CrossRef]
- Warren, R.M.; Gey van Pittius, N.C.; Barnard, M.; Hesseling, A.; Engelke, E.; de Kock, M.; Gutierrez, M.C.; Chege, G.K.; Victor, T.C.; Hoal, E.G.; et al. Differentiation of Mycobacterium tuberculosis complex by PCR amplification of genomic regions of difference. Int. J. Tuberc. Lung Dis. 2006, 10, 818–822. [Google Scholar]
- Anonymous. Tuberkulose der Rinder (Mycobacterium bovis und Mycobacterium caprae): Amtliche Methode und Falldefinition 2021. Available online: https://www.openagrar.de/receive/openagrar_mods_00054079 (accessed on 25 July 2022).
- Palmer, M.V.; Thacker, T.C.; Waters, W.R. Analysis of cytokine gene expression using a novel chromogenic in-situ hybridization method in pulmonary granulomas of cattle infected experimentally by aerosolized Mycobacterium bovis. J. Comp. Pathol. 2015, 153, 150–159. [Google Scholar] [CrossRef]
- Palmer, M.V. Emerging understanding of tuberculosis and the granuloma by comparative analysis in humans, cattle, Zebrafish, and nonhuman primates. Vet. Pathol. 2018, 55, 8–10. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BCG | VPM1002 | PDX | NUOG | ||||
|---|---|---|---|---|---|---|---|
| Distrib. | Amount | Distrib. | Amount | Distrib. | Amount | Distrib. | Amount |
| S | +++ | S | ++ | S | ++ | D | +++ |
| D | +++ | S | ++ | --- | --- | D | +++ |
| D | ++ | D | +++ | --- | --- | D | +++ |
| D | ++ | D | ++ | D | + | D | +++ |
| D | +++ | S | ++ | D | + | D | +++ |
| D | ++ | S | ++ | D | + | D | +++ |
| Vaccine | Overall Amount in Granulomas of Individual Goats | Distribution of Lipid Droplets | Size of Lipid Droplets |
|---|---|---|---|
| BCG | +++ | in epithelioid cells and MNGC, esp. close to necrosis, less in periphery, in central necrosis | small lipid droplets, numerous lipid droplets per cell, sometimes confluent |
| +++ | |||
| +++ | |||
| +++ | |||
| +++ | |||
| +++ | |||
| VPM1002 | + | in epithelioid cells | pleomorphic, few per cell |
| + | |||
| + | |||
| + | |||
| + | |||
| + | |||
| PDX | + | in epithelioid cells | pleomorphic, few per cell |
| --- | |||
| --- | |||
| - | |||
| - | |||
| - | |||
| NUOG | ++ | in epithelioid cells | highly variable in size and shape, some large |
| ++ | |||
| --- | |||
| ++ | in epithelioid cells | highly variable in size and shape, some large | |
| ++ | |||
| ++ |
| Vaccine | In Caseous Necrosis | In Inflammatory Infiltrate | Cultural Isolation | ||
|---|---|---|---|---|---|
| AFB | AG | AFB | AG | ||
| BCG | +++ | +++ | + | + | c |
| + | + | - | - | c | |
| +++ | +++ | + | + | - | |
| +++ | +++ | - | - | c | |
| + | ++ | + | - | - | |
| + | + | - | - | - | |
| VPM1002 | +++ | +++ | +++ | +++ | - |
| ++ | ++ | ++ | ++ | - | |
| +++ | +++ | - | - | - | |
| ++ | +++ | + | +++ | c | |
| +++ | +++ | - | + | c | |
| + | ++ | - | ++ | - | |
| PDX | - | + | - | + | - |
| ++ | +++ | - | - | - | |
| - | - | - | - | - | |
| ++ | --- | ++ | +++ | - | |
| NUOG | + | + | - | - | - |
| - | - | - | - | - | |
| - | + | + | ++ | - | |
| +++ | +++ | + | + | c | |
| +++ | +++ | - | - | - | |
| ++ | +++ | + | +++ | - | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liebler-Tenorio, E.M.; Heyl, J.; Wedlich, N.; Figl, J.; Köhler, H.; Krishnamoorthy, G.; Nieuwenhuizen, N.E.; Grode, L.; Kaufmann, S.H.E.; Menge, C. Vaccine-Induced Subcutaneous Granulomas in Goats Reflect Differences in Host–Mycobacterium Interactions between BCG- and Recombinant BCG-Derivative Vaccines. Int. J. Mol. Sci. 2022, 23, 10992. https://doi.org/10.3390/ijms231910992
Liebler-Tenorio EM, Heyl J, Wedlich N, Figl J, Köhler H, Krishnamoorthy G, Nieuwenhuizen NE, Grode L, Kaufmann SHE, Menge C. Vaccine-Induced Subcutaneous Granulomas in Goats Reflect Differences in Host–Mycobacterium Interactions between BCG- and Recombinant BCG-Derivative Vaccines. International Journal of Molecular Sciences. 2022; 23(19):10992. https://doi.org/10.3390/ijms231910992
Chicago/Turabian StyleLiebler-Tenorio, Elisabeth M., Johannes Heyl, Nadine Wedlich, Julia Figl, Heike Köhler, Gopinath Krishnamoorthy, Natalie E. Nieuwenhuizen, Leander Grode, Stefan H. E. Kaufmann, and Christian Menge. 2022. "Vaccine-Induced Subcutaneous Granulomas in Goats Reflect Differences in Host–Mycobacterium Interactions between BCG- and Recombinant BCG-Derivative Vaccines" International Journal of Molecular Sciences 23, no. 19: 10992. https://doi.org/10.3390/ijms231910992
APA StyleLiebler-Tenorio, E. M., Heyl, J., Wedlich, N., Figl, J., Köhler, H., Krishnamoorthy, G., Nieuwenhuizen, N. E., Grode, L., Kaufmann, S. H. E., & Menge, C. (2022). Vaccine-Induced Subcutaneous Granulomas in Goats Reflect Differences in Host–Mycobacterium Interactions between BCG- and Recombinant BCG-Derivative Vaccines. International Journal of Molecular Sciences, 23(19), 10992. https://doi.org/10.3390/ijms231910992