
Altered Surface Expression of Insulin-Degrading Enzyme on Monocytes and Lymphocytes from COVID-19 Patients Both at Diagnosis and after Hospital Discharge

 ,
, 
 , , , ,
, , , , 
Abstract
:1. Introduction
2. Results
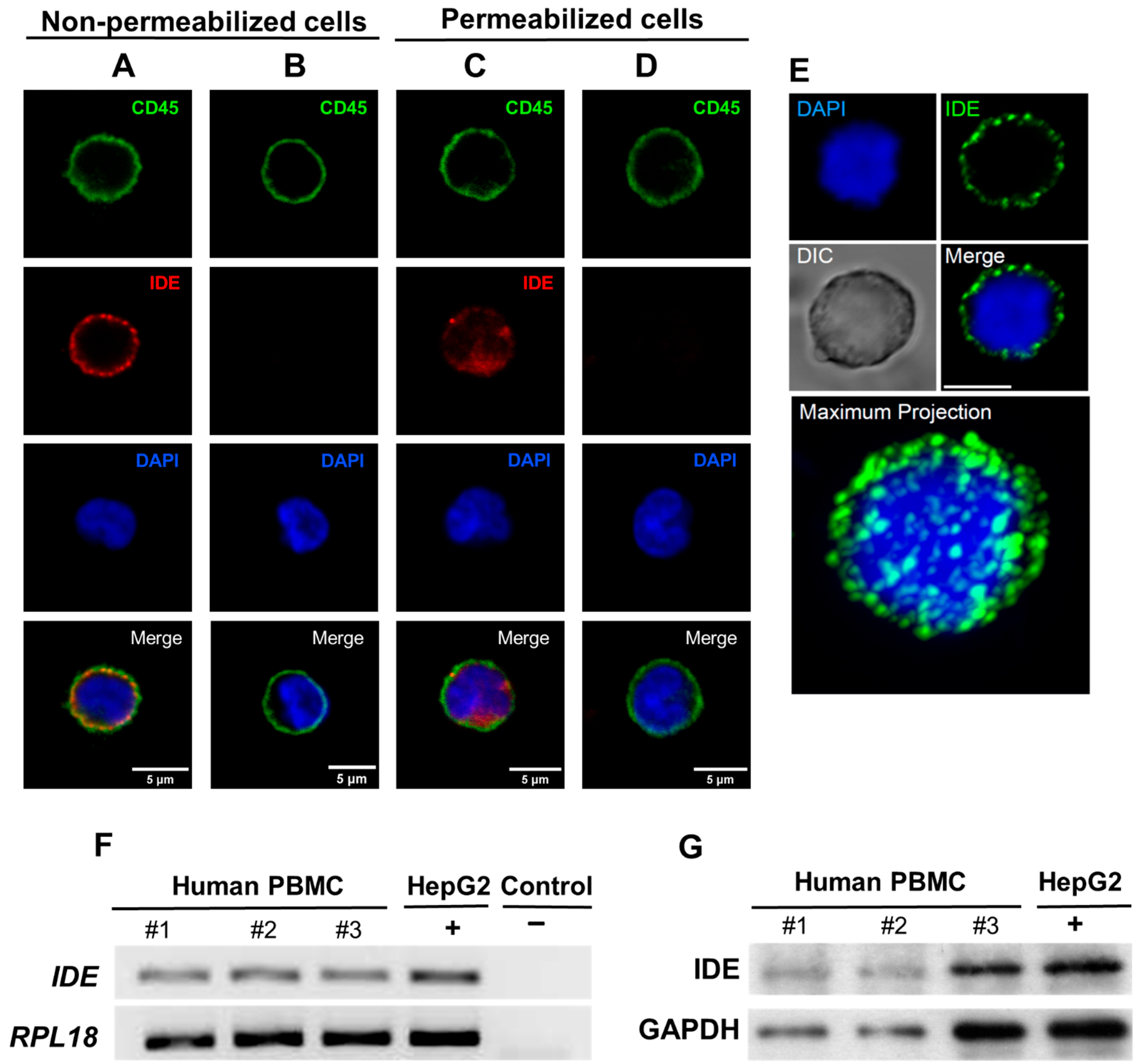
2.1. IDE Is Expressed on the Surface of Human PBMC
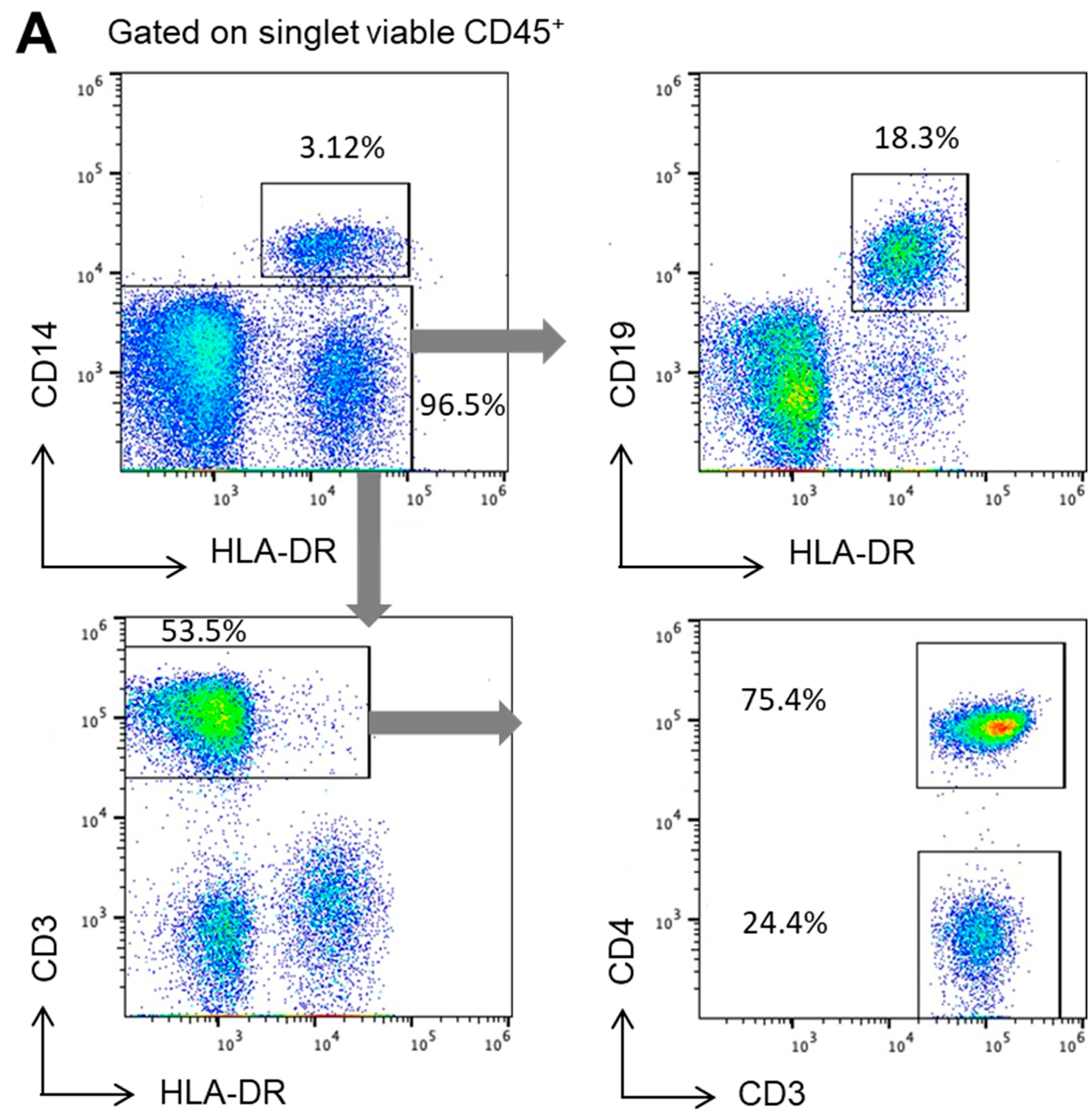
2.2. IDE Surface Expression Varies among the Different Subsets of Human PBMCs
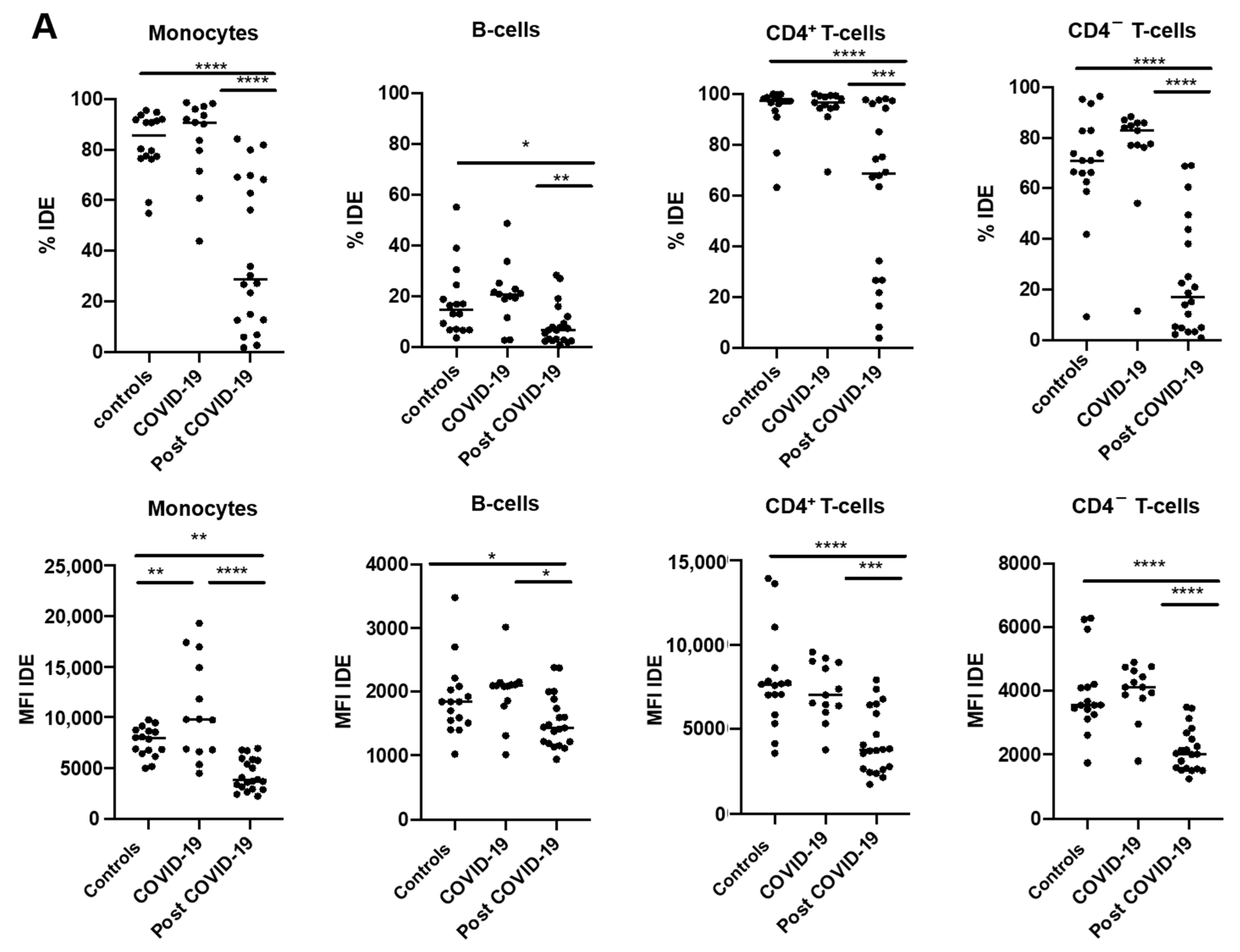
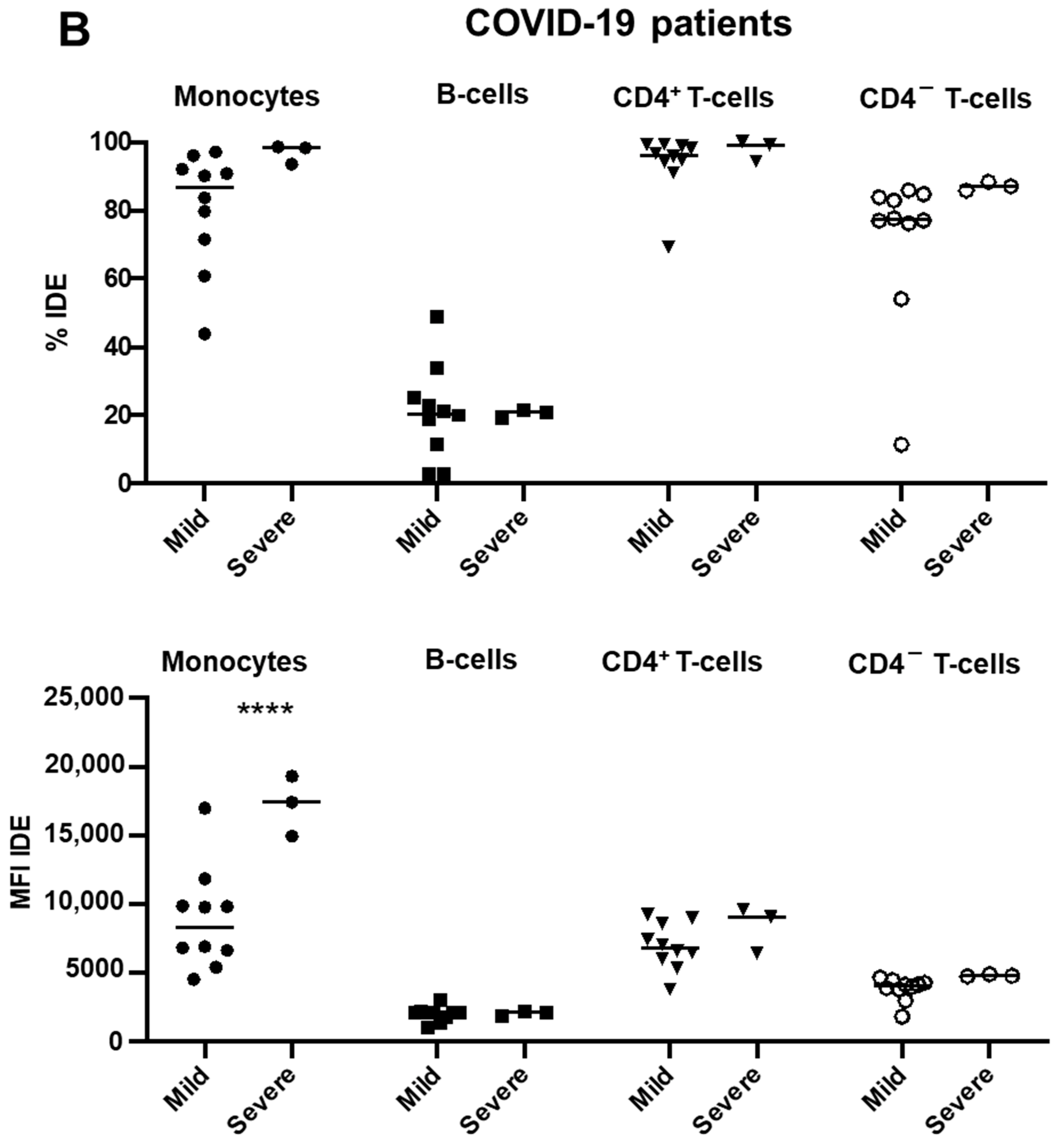
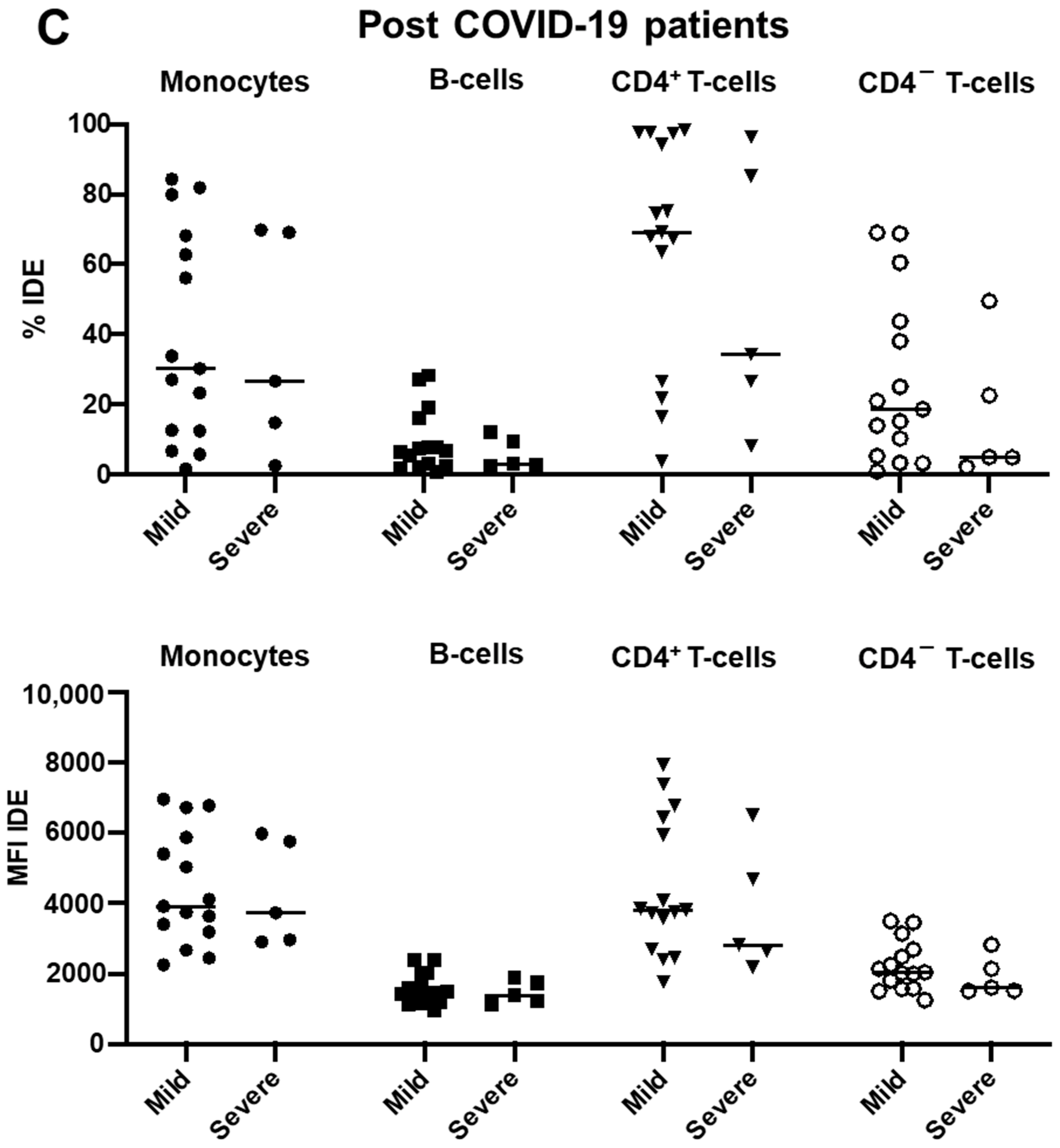
2.3. IDE Surface Expression Is Increased on Monocytes from COVID-19 Patients and Decreased in All Studied Subsets following COVID-19 Recovery
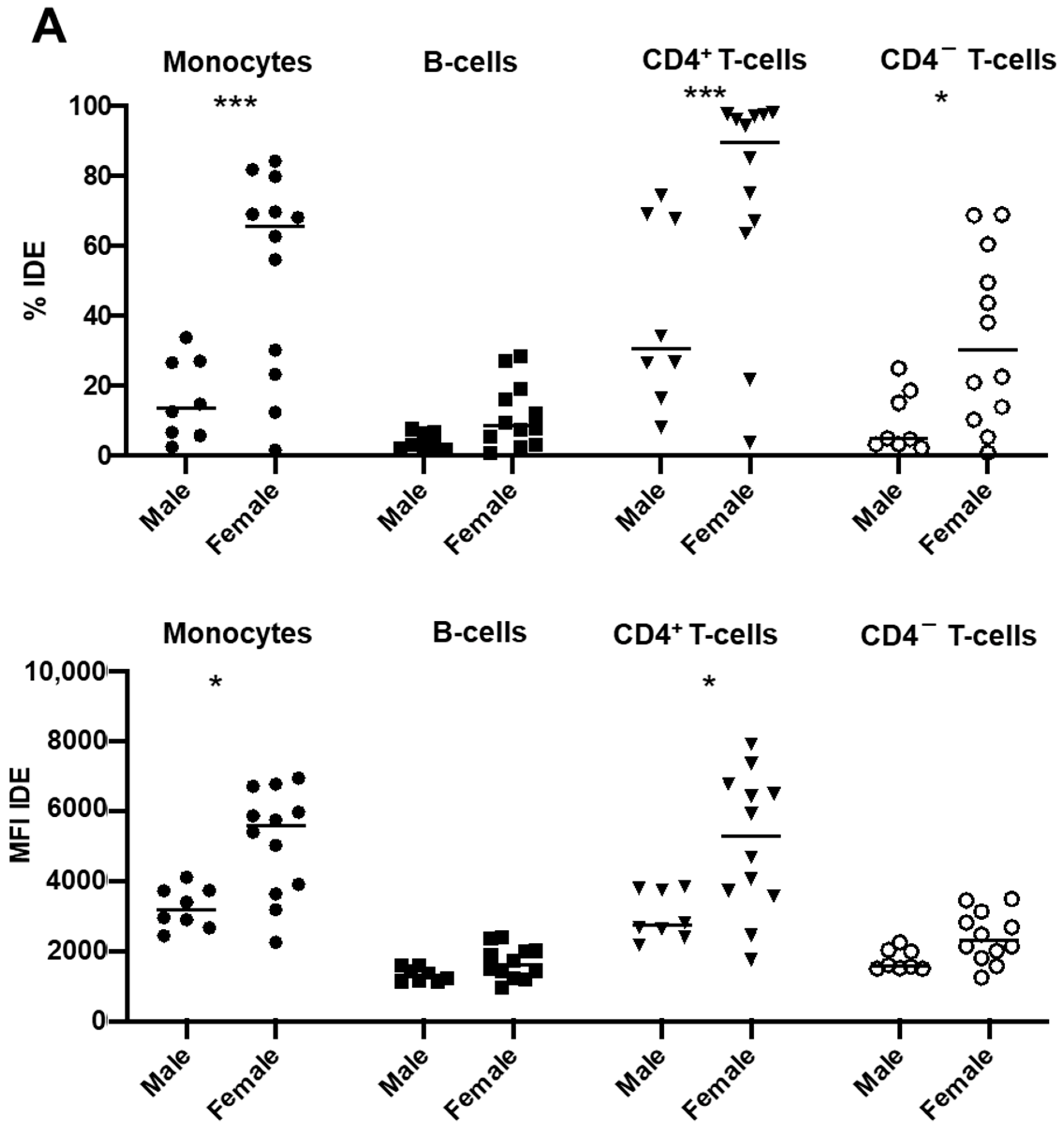
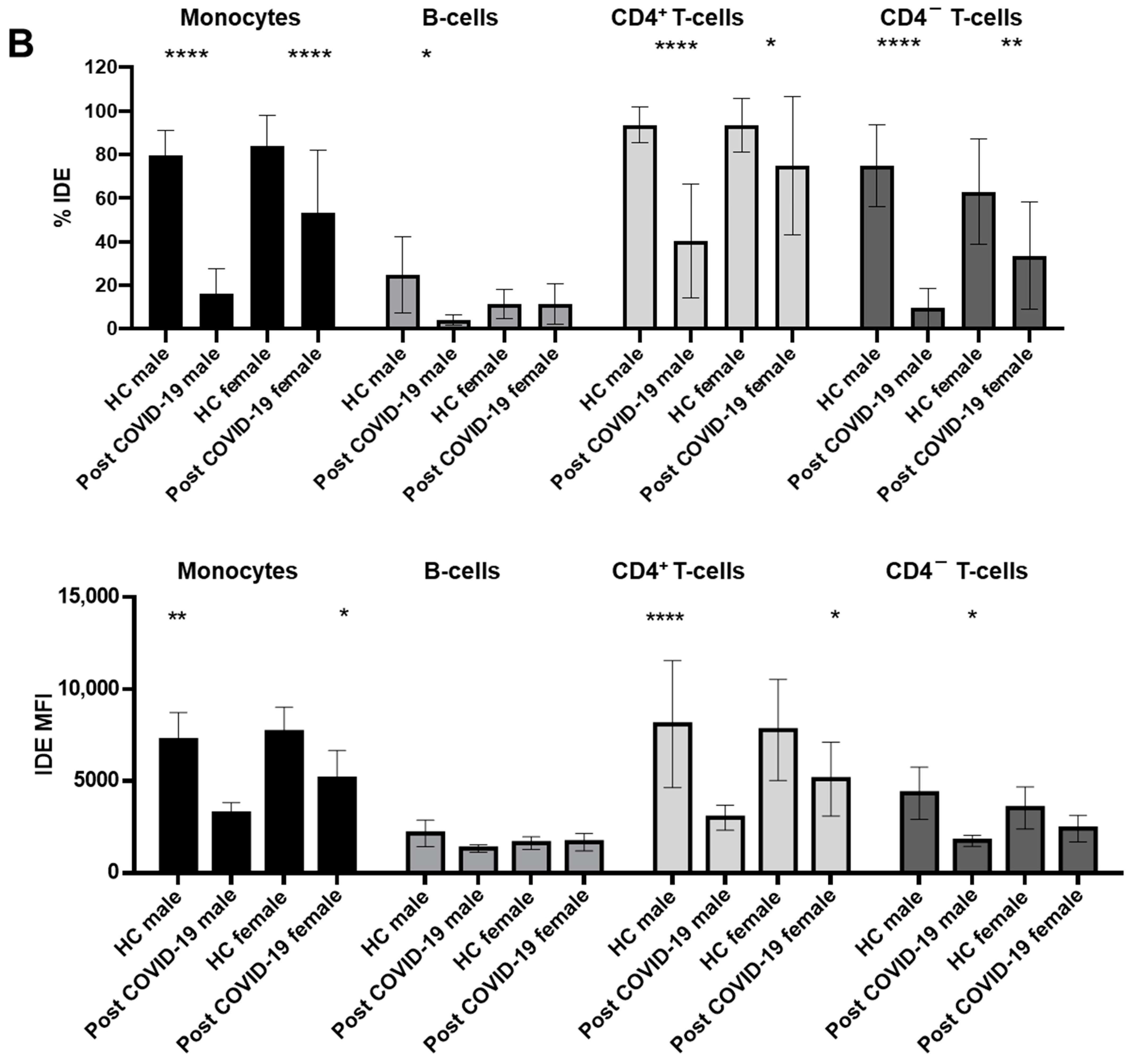
2.4. Gender Modulates IDE Surface Expression in Post-COVID-19 Patients
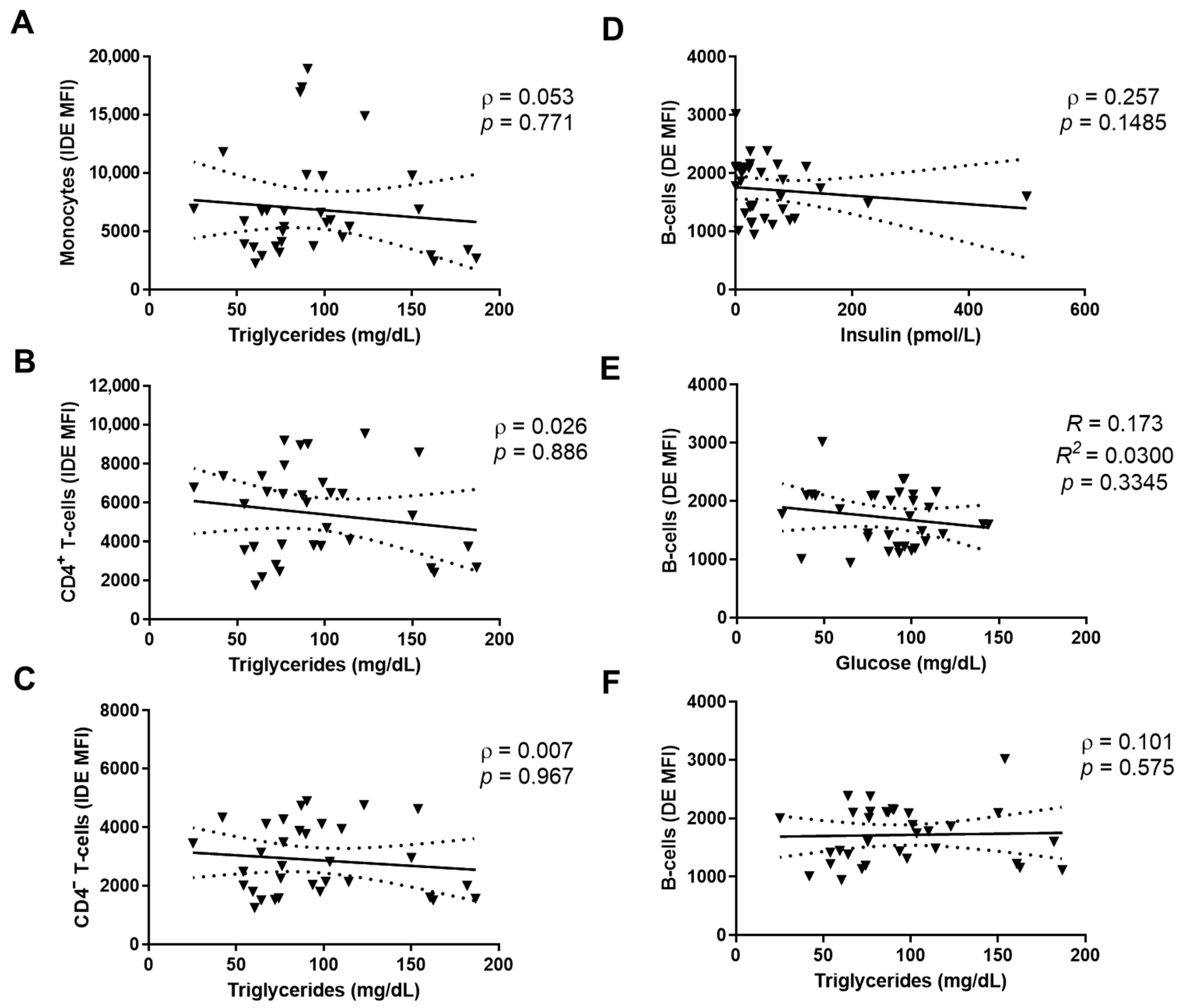
2.5. IDE Surface Expression Is Associated with Altered Metabolic Parameters
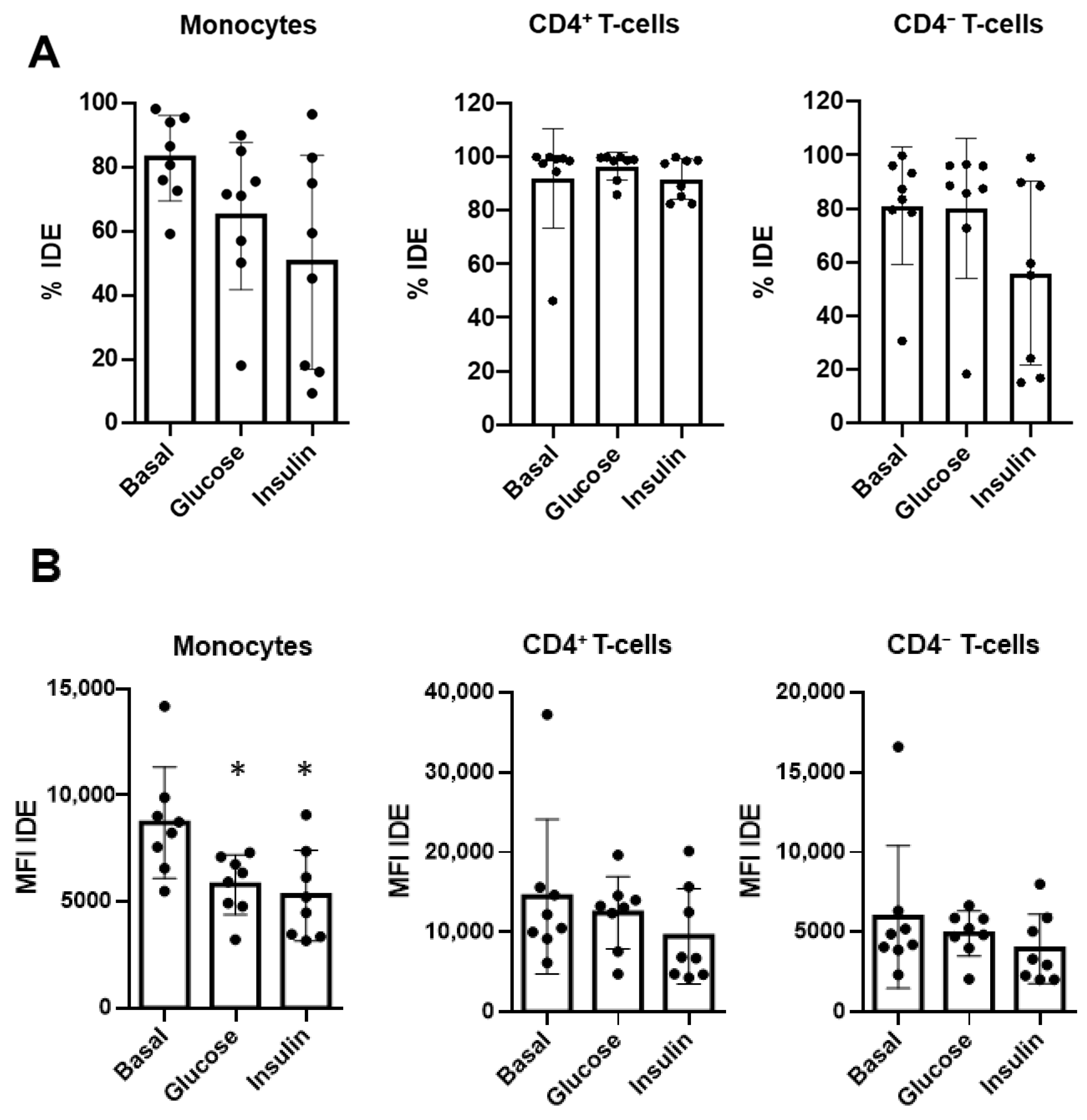
2.6. Glucose and Insulin Reduce Surface IDE Expression on Circulating Classical Monocytes
3. Discussion
4. Materials and Methods
4.1. Patient Recruitment
4.2. Biological Material
4.3. Antibody Labelling
4.4. Flow Cytometry and Data Analysis
4.5. Plasma Biochemistry and Insulin Assessments
4.6. Cell Culture
4.7. Confocal Microscopy
4.8. PCR and Electrophoresis in Agarose Gel
4.9. Western Blot Analyses
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- González-Casimiro, C.M.; Merino, B.; Casanueva-Álvarez, E.; Postigo-Casado, T.; Cámara-Torres, P.; Fernández-Díaz, C.M.; Leissring, M.A.; Cózar-Castellano, I.; Perdomo, G. Modulation of Insulin Sensitivity by Insulin-Degrading Enzyme. Biomedicines 2021, 9, 86. [Google Scholar] [CrossRef] [PubMed]
- Corraliza-Gómez, M.; Lillo, C.; Cózar-Castellano, I.; Arranz, E.; Sanchez, D.; Ganfornina, M.D. Evolutionary Origin of Insulin-Degrading Enzyme and Its Subcellular Localization and Secretion Mechanism: A Study in Microglial Cells. Cells 2022, 11, 227. [Google Scholar] [CrossRef] [PubMed]
- Mirsky, I.A.; Broh-Kahn, R.H. The inactivation of insulin by tissue extracts; the distribution and properties of insulin inactivating extracts. Arch. Biochem. 1949, 20, 1–9. [Google Scholar] [PubMed]
- Leissring, M.A.; González-Casimiro, C.M.; Merino, B.; Suire, C.N.; Perdomo, G. Targeting Insulin-Degrading Enzyme in Insulin Clearance. Int. J. Mol. Sci. 2021, 22, 2235. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Ali, M.A.; Cohen, J.I. Insulin degrading enzyme is a cellular receptor mediating varicella-zoster virus infection and cell-to-cell spread. Cell 2006, 127, 305–316. [Google Scholar] [CrossRef]
- Ali, M.A.; Li, Q.; Fischer, E.R.; Cohen, J.I. The insulin degrading enzyme binding domain of varicella-zoster virus (VZV) glycoprotein E is important for cell-to-cell spread and VZV infectivity, while a glycoprotein I binding domain is essential for infection. Virology 2009, 386, 270–279. [Google Scholar] [CrossRef]
- Li, Q.; Krogmann, T.; Ali, M.A.; Tang, W.J.; Cohen, J.I. The amino terminus of varicella-zoster virus (VZV) glycoprotein E is required for binding to insulin-degrading enzyme, a VZV receptor. J. Virol. 2007, 81, 8525–8532. [Google Scholar] [CrossRef]
- Berarducci, B.; Rajamani, J.; Zerboni, L.; Che, X.; Sommer, M.; Arvin, A.M. Functions of the unique N-terminal region of glycoprotein E in the pathogenesis of varicella-zoster virus infection. Proc. Natl. Acad. Sci. USA 2010, 107, 282–287. [Google Scholar] [CrossRef]
- Nash, Y.; Ganoth, A.; Borenstein-Auerbach, N.; Levy-Barazany, H.; Goldsmith, G.; Kopelevich, A.; Pozyuchenko, K.; Sakhneny, L.; Lazdon, E.; Blanga-Kanfi, S.; et al. From virus to diabetes therapy: Characterization of a specific insulin-degrading enzyme inhibitor for diabetes treatment. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2021, 35, e21374. [Google Scholar] [CrossRef]
- Hahn, F.; Schmalen, A.; Setz, C.; Friedrich, M.; Schlößer, S.; Kölle, J.; Spranger, R.; Rauch, P.; Fraedrich, K.; Reif, T.; et al. Proteolysis of mature HIV-1 p6 Gag protein by the insulin-degrading enzyme (IDE) regulates virus replication in an Env-dependent manner. PLoS ONE 2017, 12, e0174254. [Google Scholar] [CrossRef]
- Schmalen, A.; Karius-Fischer, J.; Rauch, P.; Setz, C.; Korn, K.; Henklein, P.; Fossen, T.; Schubert, U. The N-Terminus of the HIV-1 p6 Gag Protein Regulates Susceptibility to Degradation by IDE. Viruses 2018, 10, 710. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Zhang, Y.Y.; Chiu, S.; Hu, Z.; Lan, K.H.; Cha, H.; Sodroski, C.; Zhang, F.; Hsu, C.S.; Thomas, E.; et al. Integrative functional genomics of hepatitis C virus infection identifies host dependencies in complete viral replication cycle. PLoS Pathog. 2014, 10, e1004163. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Li, F.; Shi, Z.L. Origin and evolution of pathogenic coronaviruses. Nat. Rev. Microbiol. 2019, 17, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Ashour, H.M.; Elkhatib, W.F.; Rahman, M.M.; Elshabrawy, H.A. Insights into the Recent 2019 Novel Coronavirus (SARS-CoV-2) in Light of Past Human Coronavirus Outbreaks. Pathogens 2020, 9, 186. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.Y.; Zhang, W.; Roehrl, V.B.; Roehrl, M.W.; Roehrl, M.H. An Autoantigen-ome from HS-Sultan B-Lymphoblasts Offers a Molecular Map for Investigating Autoimmune Sequelae of COVID-19. Biorxiv Prepr. Serv. Biol. 2021. preprint. [Google Scholar] [CrossRef]
- Gordon, D.E.; Jang, G.M.; Bouhaddou, M.; Xu, J.; Obernier, K.; White, K.M.; O’Meara, M.J.; Rezelj, V.V.; Guo, J.Z.; Swaney, D.L.; et al. A SARS-CoV-2 protein interaction map reveals targets for drug repurposing. Nature 2020, 583, 459–468. [Google Scholar] [CrossRef]
- Montefusco, L.; Ben Nasr, M.; D’Addio, F.; Loretelli, C.; Rossi, A.; Pastore, I.; Daniele, G.; Abdelsalam, A.; Maestroni, A.; Dell’Acqua, M.; et al. Acute and long-term disruption of glycometabolic control after SARS-CoV-2 infection. Nat. Metab. 2021, 3, 774–785. [Google Scholar] [CrossRef]
- Duckworth, W.C. Insulin degradation: Mechanisms, products, and significance. Endocr. Rev. 1988, 9, 319–345. [Google Scholar] [CrossRef]
- Seta, K.A.; Roth, R.A. Overexpression of insulin degrading enzyme: Cellular localization and effects on insulin signaling. Biochem. Biophys. Res. Commun. 1997, 231, 167–171. [Google Scholar] [CrossRef]
- Vekrellis, K.; Ye, Z.; Qiu, W.Q.; Walsh, D.; Hartley, D.; Chesneau, V.; Rosner, M.R.; Selkoe, D.J. Neurons regulate extracellular levels of amyloid beta-protein via proteolysis by insulin-degrading enzyme. J. Neurosci. Off. J. Soc. Neurosci. 2000, 20, 1657–1665. [Google Scholar] [CrossRef] [Green Version]
- Song, E.S.; Jang, H.; Guo, H.F.; Juliano, M.A.; Juliano, L.; Morris, A.J.; Galperin, E.; Rodgers, D.W.; Hersh, L.B. Inositol phosphates and phosphoinositides activate insulin-degrading enzyme, while phosphoinositides also mediate binding to endosomes. Proc. Natl. Acad. Sci. USA 2017, 114, E2826–E2835. [Google Scholar] [CrossRef] [PubMed]
- McIlhinney, R.A. Membrane targeting via protein N-myristoylation. Methods Mol. Biol. 1998, 88, 211–225. [Google Scholar] [CrossRef] [PubMed]
- Udenwobele, D.I.; Su, R.C.; Good, S.V.; Ball, T.B.; Varma Shrivastav, S.; Shrivastav, A. Myristoylation: An Important Protein Modification in the Immune Response. Front. Immunol. 2017, 8, 751. [Google Scholar] [CrossRef]
- Rubartelli, A.; Cozzolino, F.; Talio, M.; Sitia, R. A novel secretory pathway for interleukin-1 beta, a protein lacking a signal sequence. EMBO J. 1990, 9, 1503–1510. [Google Scholar] [CrossRef]
- Hospital, V.; Chesneau, V.; Balogh, A.; Joulie, C.; Seidah, N.G.; Cohen, P.; Prat, A. N-arginine dibasic convertase (nardilysin) isoforms are soluble dibasic-specific metalloendopeptidases that localize in the cytoplasm and at the cell surface. Biochem. J. 2000, 349, 587–597. [Google Scholar] [CrossRef] [PubMed]
- Nishi, E.; Prat, A.; Hospital, V.; Elenius, K.; Klagsbrun, M. N-arginine dibasic convertase is a specific receptor for heparin-binding EGF-like growth factor that mediates cell migration. EMBO J. 2001, 20, 3342–3350. [Google Scholar] [CrossRef] [PubMed]
- Crack, P.J.; Wu, T.J.; Cummins, P.M.; Ferro, E.S.; Tullai, J.W.; Glucksman, M.J.; Roberts, J.L. The association of metalloendopeptidase EC 3.4.24.15 at the extracellular surface of the AtT-20 cell plasma membrane. Brain Res. 1999, 835, 113–124. [Google Scholar] [CrossRef]
- Kandel, J.; Bossy-Wetzel, E.; Radvanyi, F.; Klagsbrun, M.; Folkman, J.; Hanahan, D. Neovascularization is associated with a switch to the export of bFGF in the multistep development of fibrosarcoma. Cell 1991, 66, 1095–1104. [Google Scholar] [CrossRef]
- Allen, S.J.; Crown, S.E.; Handel, T.M. Chemokine: Receptor structure, interactions, and antagonism. Annu. Rev. Immunol. 2007, 25, 787–820. [Google Scholar] [CrossRef]
- Ren, M.; Guo, Q.; Guo, L.; Lenz, M.; Qian, F.; Koenen, R.R.; Xu, H.; Schilling, A.B.; Weber, C.; Ye, R.D.; et al. Polymerization of MIP-1 chemokine (CCL3 and CCL4) and clearance of MIP-1 by insulin-degrading enzyme. EMBO J. 2010, 29, 3952–3966. [Google Scholar] [CrossRef] [Green Version]
- Chevrier, S.; Zurbuchen, Y.; Cervia, C.; Adamo, S.; Raeber, M.E.; de Souza, N.; Sivapatham, S.; Jacobs, A.; Bachli, E.; Rudiger, A.; et al. A distinct innate immune signature marks progression from mild to severe COVID-19. Cell Rep. Med. 2021, 2, 100166. [Google Scholar] [CrossRef] [PubMed]
- Merino, B.; Fernández-Díaz, C.M.; Parrado-Fernández, C.; González-Casimiro, C.M.; Postigo-Casado, T.; Lobatón, C.D.; Leissring, M.A.; Cózar-Castellano, I.; Perdomo, G. Hepatic insulin-degrading enzyme regulates glucose and insulin homeostasis in diet-induced obese mice. Metabolism 2020, 113, 154352. [Google Scholar] [CrossRef] [PubMed]
- Ortega Moreno, L.; Fernández-Tomé, S.; Chaparro, M.; Marin, A.C.; Mora-Gutiérrez, I.; Santander, C.; Baldan-Martin, M.; Gisbert, J.P.; Bernardo, D. Profiling of Human Circulating Dendritic Cells and Monocyte Subsets Discriminates Between Type and Mucosal Status in Patients With Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2021, 27, 268–274. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| COVID-19 | Post-COVID-19 | |||
|---|---|---|---|---|
| Female | Male | Female | Male | |
| Number of patients (n) | 13 | 20 | ||
| 5 | 8 | 12 | 8 | |
| Glucose (mg/dL) | 67.0 ± 8.0 | 98.5 ± 4.2 * | ||
| 64.2 ± 13.5 | 68.8 ± 9.8 | 93.0 ± 3.6 * | 106.8 ± 8.3 * | |
| Insulin (pmol/L) | 22.7 ± 9.4 | 85.9 ± 23.9 * | ||
| 20.8 ± 11.6 | 24.0 ± 13.4 | 67.7 ± 17.3 * | 113.2 ± 52.5 * | |
| Cholesterol (mg/dL) | 102.2 ± 6.0 | 125.9 ± 3.9 * | ||
| 107.7 ± 8.6 | 98.8 ± 7.9 | 124.6 ± 4.6 | 127.8 ± 6.9 * | |
| Triglycerides (mg/dL) | 98.0 ± 8.3 | 93.1 ± 10.0 | ||
| 74.8 ± 8.8 | 112.4 ± 9.1 $ | 72.0 ± 6.9 | 124.7 ± 17.5 $ | |
| Gender | Age | Days in Hospital | Severity | |
|---|---|---|---|---|
| Control.1 | Male | 46 | n/a | n/a |
| Control.2 | Male | 45 | n/a | n/a |
| Control.3 | Male | 60 | n/a | n/a |
| Control.4 | Male | 64 | n/a | n/a |
| Control.5 | Male | 62 | n/a | n/a |
| Control.6 | Male | 66 | n/a | n/a |
| Control.7 | Male | 65 | n/a | n/a |
| Control.8 | Female | 46 | n/a | n/a |
| Control.9 | Female | 60 | n/a | n/a |
| Control.10 | Female | 41 | n/a | n/a |
| Control.11 | Female | 63 | n/a | n/a |
| Control.12 | Female | 45 | n/a | n/a |
| Control.13 | Female | 63 | n/a | n/a |
| Control.14 | Female | 45 | n/a | n/a |
| Control.15 | Female | 61 | n/a | n/a |
| COVID-19.1 | Male | 47 | 6 | mild |
| COVID-19.2 | Male | 32 | 3 | mild |
| COVID-19.3 | Male | 75 | 4 | mild |
| COVID-19.4 | Male | 62 | 4 | mild |
| COVID-19.5 | Male | 61 | 4 | mild |
| COVID-19.6 | Male | 33 | 0 | mild |
| COVID-19.7 | Male | 89 | Exitus | severe |
| COVID-19.8 | Male | 83 | Exitus | severe |
| COVID-19.9 | Female | 62 | 0 | mild |
| COVID-19.10 | Female | 35 | 0 | mild |
| COVID-19.11 | Female | 52 | 0 | mild |
| COVID-19.12 | Female | 82 | 0 | mild |
| COVID-19.13 | Female | 88 | 40 | severe |
| Post-COVID-19.1 | Male | 62 | 6 | mild |
| Post-COVID-19.2 | Male | 75 | 4 | mild |
| Post-COVID-19.3 | Male | 51 | 6 | mild |
| Post-COVID-19.4 | Male | 47 | 6 | mild |
| Post-COVID-19.5 | Male | 75 | 4 | mild |
| Post-COVID-19.6 | Male | 41 | 23 | severe |
| Post-COVID-19.7 | Male | 75 | 15 | severe |
| Post-COVID-19.8 | Male | 64 | 12 | severe |
| Post-COVID-19.9 | Female | 64 | 8 | mild |
| Post-COVID-19.10 | Female | 79 | 2 | mild |
| Post-COVID-19.11 | Female | 91 | 7 | mild |
| Post-COVID-19.12 | Female | 59 | 8 | mild |
| Post-COVID-19.13 | Female | 62 | 5 | mild |
| Post-COVID-19.14 | Female | 58 | 5 | mild |
| Post-COVID-19.15 | Female | 42 | 3 | mild |
| Post-COVID-19.16 | Female | 56 | 4 | mild |
| Post-COVID-19.17 | Female | 59 | 3 | mild |
| Post-COVID-19.18 | Female | 59 | 7 | mild |
| Post-COVID-19.19 | Female | 81 | 11 | severe |
| Post-COVID-19.20 | Female | 51 | 16 | severe |
| Specificity | Source | Conjugate | Clone | Manufacturer |
|---|---|---|---|---|
| CD3 | Mouse | APC | HIT3a | Biolegend |
| CD14 | Mouse | Alexa700 | 61D3 | eBioscience |
| CD19 | Mouse | PE-Cy5 | HIB19 | Becton Dickinson |
| CD45 | Mouse | PE-Cy7 | HI30 | Becton Dickinson |
| CD45 | Mouse | FITC | 304038 | Biolegend |
| HLA-DR | Mouse | BV510 | L243 | Biolegend |
| IDE | Rabbit | n/a | AB9210 | Sigma-Aldrich |
| Rabbit IgG | Donkey | FITC | Poly4064 | Biolegend |
| Rabbit IgG | Goat | Alexa488 | A11070 | Invitrogen |
| Rabbit IgG | Goat | Alexa594 | A11012 | Invitrogen |
| GAPDH | Mouse | n/a | MAB374 | Sigma-Aldrich |
| Rabbit IgG | Donkey | HRP | 711-035-152 | Jackson Immunoresearch |
| Mouse IgG | Sheep | HRP | NA9310 | Amersham |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Casimiro, C.M.; Arribas-Rodríguez, E.; Fiz-López, A.; Casas, J.; Gutiérrez, S.; Tellería, P.; Novoa, C.; Rojo-Rello, S.; Tamayo, E.; Orduña, A.; et al. Altered Surface Expression of Insulin-Degrading Enzyme on Monocytes and Lymphocytes from COVID-19 Patients Both at Diagnosis and after Hospital Discharge. Int. J. Mol. Sci. 2022, 23, 11070. https://doi.org/10.3390/ijms231911070
González-Casimiro CM, Arribas-Rodríguez E, Fiz-López A, Casas J, Gutiérrez S, Tellería P, Novoa C, Rojo-Rello S, Tamayo E, Orduña A, et al. Altered Surface Expression of Insulin-Degrading Enzyme on Monocytes and Lymphocytes from COVID-19 Patients Both at Diagnosis and after Hospital Discharge. International Journal of Molecular Sciences. 2022; 23(19):11070. https://doi.org/10.3390/ijms231911070
Chicago/Turabian StyleGonzález-Casimiro, Carlos M., Elisa Arribas-Rodríguez, Aida Fiz-López, Javier Casas, Sara Gutiérrez, Pablo Tellería, Cristina Novoa, Silvia Rojo-Rello, Eduardo Tamayo, Antonio Orduña, and et al. 2022. "Altered Surface Expression of Insulin-Degrading Enzyme on Monocytes and Lymphocytes from COVID-19 Patients Both at Diagnosis and after Hospital Discharge" International Journal of Molecular Sciences 23, no. 19: 11070. https://doi.org/10.3390/ijms231911070
APA StyleGonzález-Casimiro, C. M., Arribas-Rodríguez, E., Fiz-López, A., Casas, J., Gutiérrez, S., Tellería, P., Novoa, C., Rojo-Rello, S., Tamayo, E., Orduña, A., Dueñas, C., Bernardo, D., & Perdomo, G. (2022). Altered Surface Expression of Insulin-Degrading Enzyme on Monocytes and Lymphocytes from COVID-19 Patients Both at Diagnosis and after Hospital Discharge. International Journal of Molecular Sciences, 23(19), 11070. https://doi.org/10.3390/ijms231911070