


Response of Rice with Overlapping Growth Stages to Water Stress by Assimilates Accumulation and Transport and Starch Synthesis of Superior and Inferior Grains
 ,
,
Abstract
:1. Introduction
2. Results
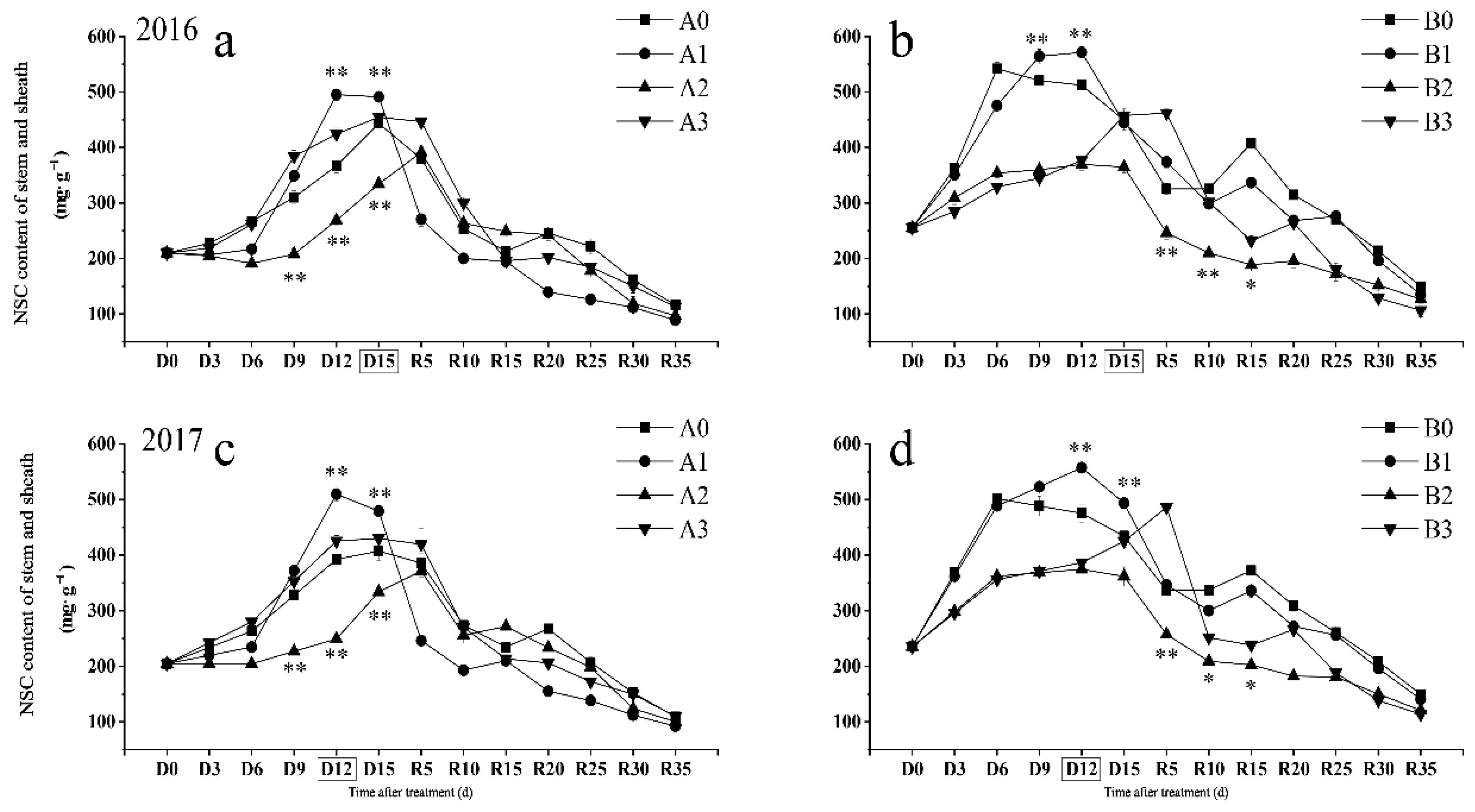
2.1. Changes in NSC Content Response to Drought Stress in Stems and Sheaths
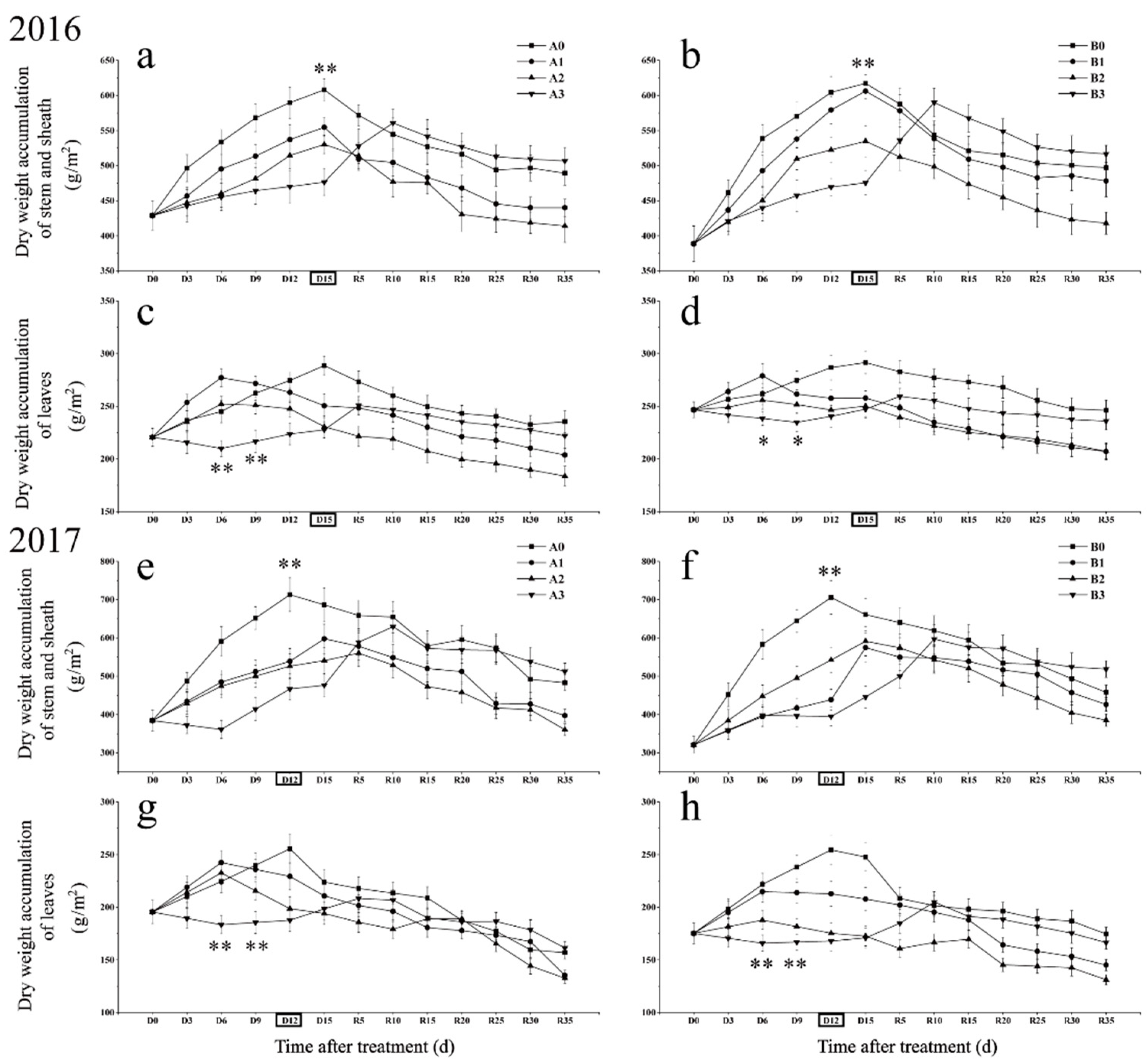
2.2. Differences in Dry Matter Accumulation in Stems and Leaves under Different Drought Stress
2.3. Difference in Grain Filling between Superior and Inferior Grains under Drought Stress
2.4. Effects of Drought Stress on Panicle Traits of Rice with Overlapping Growth Stages
2.5. Effects of Drought Stress on Yield Components in Rice
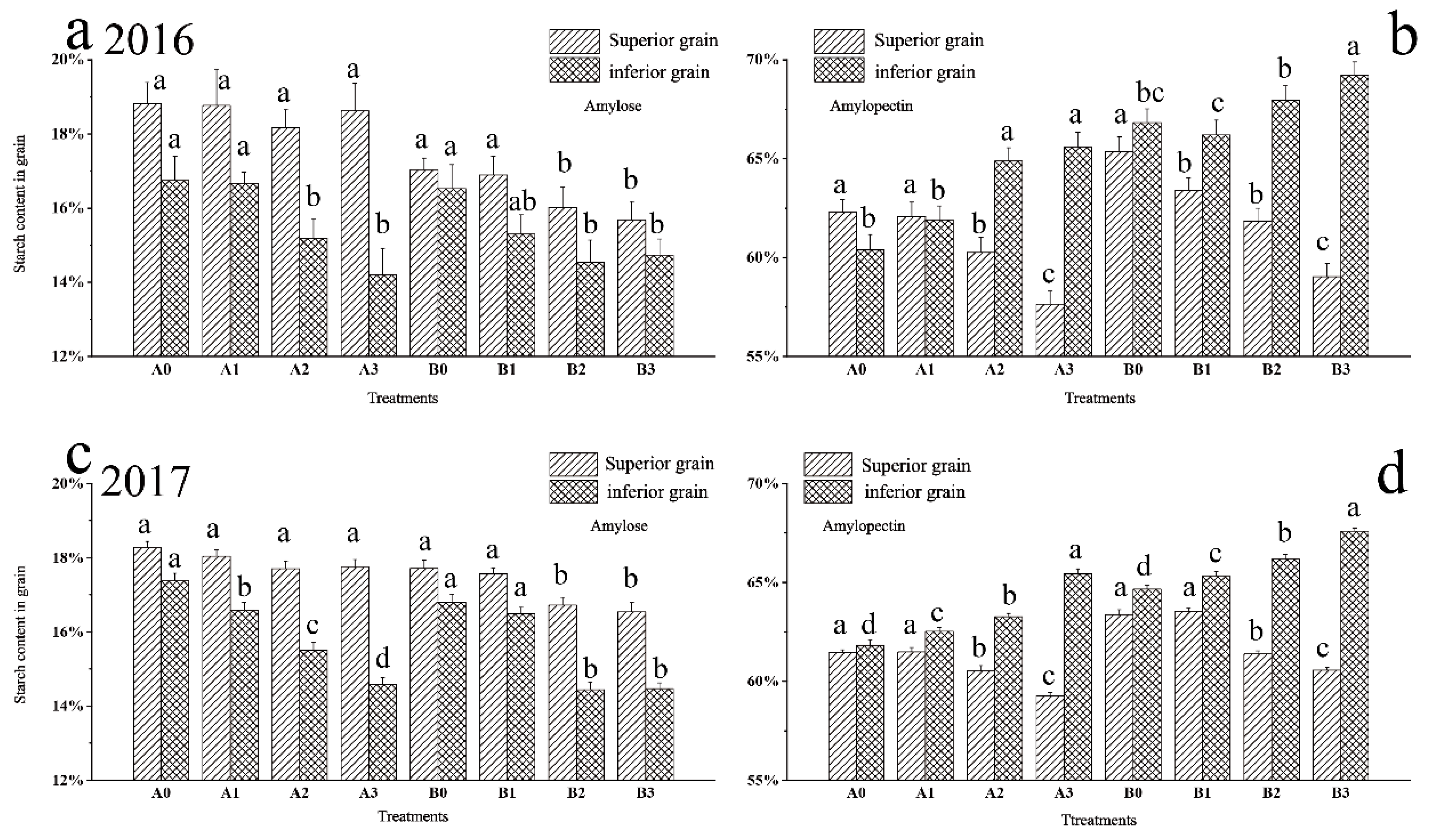
2.6. Difference between Starch Composition of Superior and Inferior Grains
2.7. Physiological Differences in Starch Synthesis between Superior and Inferior Grains
2.7.1. Soluble Starch Synthase
2.7.2. Granule-Bound Starch Synthetase
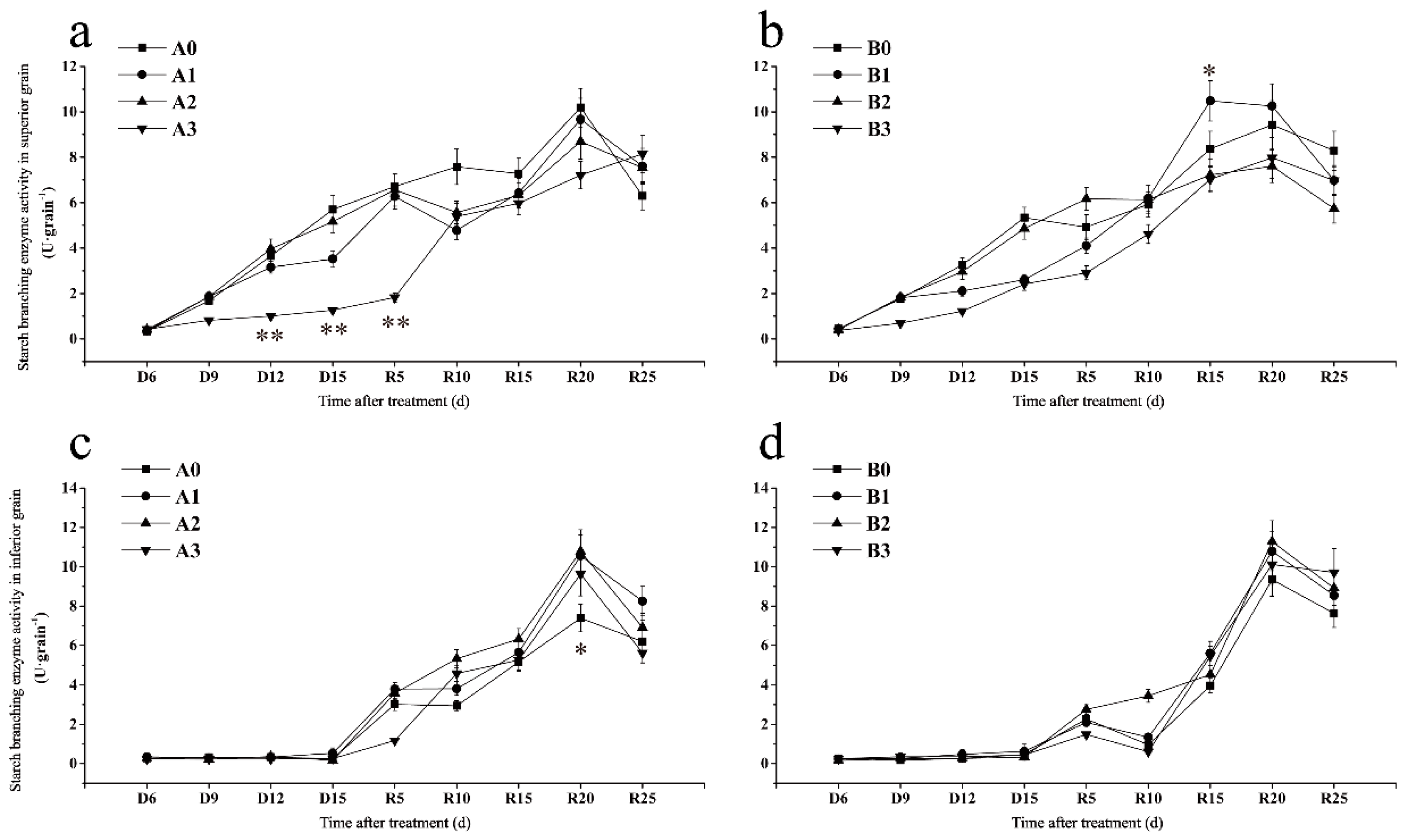
2.7.3. Starch Branching Enzyme
2.8. Correlation between Dry Matter Accumulation, Transportation and Grain-Filling
3. Discussion
3.1. Different Degrees of Drought Stress May Change the Rule of Dry Matter Accumulation
3.2. Difference in Grain-Filling Strategies between Superior and Inferior Grains under Drought Stress
3.3. Effects of Dry Matter Transportation on Yield Formation under Drought Stress
3.4. Effects of Drought Stress on Starch Synthesis of Superior and Inferior Grains
4. Material and Methods
4.1. Plant Material and Growth Conditions
4.2. Experimental Design
4.3. Determination of Indexes
4.3.1. Sample Collection
4.3.2. Amylose and Amylopectin
4.3.3. Soluble Starch Synthase and Granule Bound-Starch Synthase (GBSS)
4.3.4. Starch Branching Enzyme (SBE)
4.3.5. Dry Matter Accumulation and Grain Filling Dynamics
4.4. Yield and Yield Components
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- National Bureau of Statistics of China. China Statistical Yearbook; National Bureau of Statistics of China: Beijing, China, 2017.
- Todaka, D.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Recent advances in the dissection of drought-stress regulatory networks and strategies for development of drought-tolerant transgenic rice plants. Front. Plant Sci. 2015, 6, 84. [Google Scholar] [CrossRef] [PubMed]
- Greenham, K.; Guadagno, C.R.; Gehan, M.A.; Mockler, T.C.; Weinig, C.; Ewers, B.E.; McClung, C.R. Temporal network analysis identifies early physiological and transcriptomic indicators of mild drought in Brassica rapa. eLife 2017, 6, e29655. [Google Scholar] [CrossRef] [PubMed]
- National Bureau of Statistics of China. China Statistical Yearbook; National Bureau of Statistics of China: Beijing, China, 2020.
- Singh, A.K.; Choudhury, B.U.; Bouman, B.A.M. The international workshop on water-wise rice production. In Water-Wise Rice Production; Bouman, B.A.M., Hengsdijk, H., Hardy, B., Bindraban, P.S., Tuong, T.P., Ladha, J.K., Eds.; International Rice Research Institute: LosBaños, Philippines, 2002; pp. 237–248. [Google Scholar]
- Okamura, M.; Arai Sanoh, Y.; Yoshida, H.E.A. Characterization of high-yielding rice cultivars with different grain-filling properties to clarify limiting factors for improving grain yield. Field Crops Res. 2018, 219, 139–147. [Google Scholar] [CrossRef]
- Mcdowell, N.G. Mechanisms Linking Drought, Hydraulics, Carbon Metabolism, and Vegetation Mortality. Plant Physiol. 2011, 155, 1051–1059. [Google Scholar] [CrossRef] [PubMed]
- Luxmoore, R.; Oren, R.; Sheriff, D.; Thomas, R. Resource Physiology of Conifers; Academic Press: Cambridge, MA, USA, 1995. [Google Scholar]
- Estiarte, M.; Uelas, J.P. Excess carbon: The relationship with phenotypical plasticity in storage and defense functions of plants. Orsis Org. I Sist. 1999, 14, 159–203. [Google Scholar]
- Kozlowski, T.T.; Pallardy, S.G. Acclimation and adaptive responses of woody plants to environmental stresses. Bot. Rev. 2002, 68, 270–334. [Google Scholar] [CrossRef]
- Hummel, I.; Pantin, F.; Sulpice, R.; Piques, M.; Rolland, G.; Dauzat, M.; Christophe, A.; Pervent, M.; Bouteille, M.; Stitt, M. Arabidopsis Plants Acclimate to Water Deficit at Low Cost through Changes of Carbon Usage: An Integrated Perspective Using Growth, Metabolite, Enzyme, and Gene Expression Analysis. Plant Physiol. 2010, 154, 357–372. [Google Scholar] [CrossRef]
- Pinheiro, C.; Chaves, M.M. Photosynthesis and drought: Can we make metabolic connections from available data? J. Exp. Bot. 2011, 62, 869–882. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, J.; Wang, Z.; Zhu, Q.; Wang, W. Remobilization of carbon reserves in response to water deficit during grain filling of rice. Field Crops Res. 2001, 71, 47–55. [Google Scholar] [CrossRef]
- Xu, Y.; Xu, Y.; Li, Y.; Qian, X.; Wang, Z.; Yang, J. Effect of Alternate Wetting and Drying Irrigation on Post-anthesis Remobilization of Assimilates and Grain Filling of Rice. Acta Agron. Sin. 2018, 44, 554–568. [Google Scholar] [CrossRef]
- Xu, Z.; Yu, Z.; Zhang, Y. The Effects of Soil Moisture on Grain Starch Synthesis and Accumulation of Winter Wheat. Acta Agron. Sin. 2003, 29, 595–600. [Google Scholar]
- Zhang, W.Y.; Sheng, J.Y.; Xu, Y.J.; Xiong, F.; Wu, Y.F.; Wang, W.L.; Wang, Z.Q.; Yang, J.C.; Zhang, J.H. Role of brassinosteroids in rice spikelet differentiation and degeneration under soil-drying during panicle development. BMC Plant Biol. 2019, 19, 16. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Fang, C.; Qian, Z.; Guo, B.; Huo, Z. Differences in starch structure, physicochemical properties and texture characteristics in superior and inferior grains of rice varieties with different amylose contents. Food Hydrocoll. 2021, 110, 106170. [Google Scholar] [CrossRef]
- You, C.; Zhu, H.; Xu, B.; Huang, W.; Wang, S.; Ding, Y.; Liu, Z.; Li, G.; Chen, L.; Ding, C.; et al. Effect of Removing Superior Spikelets on Grain Filling of Inferior Spikelets in Rice. Front. Plant Sci. 2016, 7, 1161. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Y.Y.; Song, N.Y.; Chen, Q.L.; Sun, H.Z.; Peng, T.; Huang, S.; Zhao, Q.Z. Response of grain-filling rate and grain quality of mid-season indica rice to nitrogen application. J. Integr. Agric. 2021, 20, 1465–1473. [Google Scholar] [CrossRef]
- Wang, Z.Q.; Ye, Y.; Yang, J.; Yuan, L.M.; Wang, X.; Zhu, Q. Changes and Regulations of Sucrose Synthase Activity in Rice Grains during Grain Filling. Acta Agron. Sin. 2004, 30, 634–643. [Google Scholar]
- Ishimaru, T.; Matsuda, T.; Ohsugi, R.; Yamagishi, T. Morphological development of rice caryopses located at the different positions in a panicle from early to middle stage of grain filling. Funct. Plant Biol. 2003, 30, 1139–1149. [Google Scholar] [CrossRef]
- Ishimaru, T.; Hirose, T.; Matsuda, T.; Goto, A.; Takahashi, K.; Sasaki, H.; Terao, T.; Ishii, R.; Ohsugi, R.; Yamagishi, T. Expression patterns of genes encoding carbohydrate-metabolizing enzymes and their relationship to grain filling in rice (Oryza sativa L.): Comparison of caryopses located at different positions in a panicle. Plant Cell Physiol. 2005, 46, 620–628. [Google Scholar] [CrossRef]
- Fu, J.; Huang, Z.; Wang, Z.; Yang, J.; Zhang, J. Pre-anthesis non-structural carbohydrate reserve in the stem enhances the sink strength of inferior spikelets during grain filling of rice. Field Crops Res. 2011, 123, 170–182. [Google Scholar] [CrossRef]
- Zhang, H.; Li, H.W.; Yuan, L.M.; Wang, Z.Q.; Yang, J.C.; Zhang, J.H. Post-anthesis alternate wetting and moderate soil drying enhances activities of key enzymes in sucrose-to-starch conversion in inferior spikelets of rice. J. Exp. Bot. 2012, 63, 215–227. [Google Scholar] [CrossRef]
- Wang, G.Q.; Li, H.X.; Feng, L.; Chen, M.X.; Meng, S.; Ye, N.H.; Zhang, J.H. Transcriptomic analysis of grain filling in rice inferior grains under moderate soil drying. J. Exp. Bot. 2019, 70, 1597–1611. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Deng, Y.; Zhu, H.; Hu, Y.; Jiang, Z.; Tang, S.; Wang, S.; Ding, Y. The Initiation of Inferior Grain Filling is Affected by Sugar Translocation Efficiency in Large Panicle Rice. Rice 2019, 12, 75. [Google Scholar] [CrossRef] [PubMed]
- Shao, X.; Zhang, R.; Qi, C.; Tong, S.; Yang, M. Effects of Water Stress on Growth and Yield of Rice in Jointing-Booting Stage. J. Jilin Agric. Univ. 2004, 26, 237–241. [Google Scholar]
- Yang, J.; Zhu, Q.; Wang, Z. Effects of Soil Moisture on the Yield and Physiological Chaaracteristics in Rice. Acta Agron. Sin. 1995, 21, 110–114. [Google Scholar]
- Wang, C.; Wang, B.; Zhang, W.; Zhao, L.; Zhao, X.; Gao, L.; Hou, W. Effects of Drought Stress at Different Growth Stages on G rain Yield and Milling Quality of Rice. Chin. J. Rice Sci. 2007, 21, 643–649. [Google Scholar]
- Yang, J.; Zhang, J.; Liu, K.; Wang, Z.; Liu, L. Abscisic Acid and Ethylene Interact in Rice Spikelets in Response to Water Stress During Meiosis. J. Plant Growth Regul. 2007, 26, 318–328. [Google Scholar] [CrossRef]
- Zhang, W.; Chen, Y.; Wang, Z.; Yang, J. Polyamines and ethylene in rice young panicles in response to soil drought during panicle differentiation. Plant Growth Regul. 2017, 82, 491–503. [Google Scholar] [CrossRef]
- Yang, J.C.; Zhang, J.H.; Huang, Z.L. Remobilization of carbon reserves is improved by controlled soil-drying during grain filling of wheat. Crop Sci. 2000, 40, 1645–1655. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, J.; Yang, J.; Zhu, Q. Effect of Water Stress on Metabolism of Stored Carbohydrate of Stem and Yield in Rice Grown under Unfavorable-delayed Senescence. Acta Agron. Sin. 2004, 30, 196–204. [Google Scholar]
- Wang, X.; Liu, H.; Zhang, D.; Zou, D.; Wang, J.; Zheng, H.; Jia, Y.; Qu, Z.; Sun, B.; Zhao, H. Photosynthetic Carbon Fixation and Sucrose Metabolism Supplemented by Weighted Gene Co-expression Network Analysis in Response to Water Stress in Rice with Overlapping Growth Stages. Front. Plant Sci. 2022, 13, 864605. [Google Scholar] [CrossRef]
- Yang, J. Mechanism and Regulation in the Filling of Inferior Spikelets of Rice. Acta Agron. Sin. 2010, 36, 2011–2019. [Google Scholar]
- Zhong, X.; Zhang, P.; Zeng, X.; Xu, Y. The difference in chalkiness between superior and inferior grains and the relation between chalkiness and grain weight. Acta Agric. Univ. Jiangxiensis 1996, 18, 154–159. [Google Scholar]
- Yang, J.; Zhu, Q.; Wang, Z.; Lang, Y. Effects of Moisture on Grain Weight-Increasing Process of Rice. J. Jiangsu Agric. Coll. 1994, 15, 9–14. [Google Scholar]
- Zhu, H.; Li, G.; Xia, Q.; Long, R.; Deng, A.; Huang, J.; Xiang, H.; Yang, C. Effects of Drought Stress on Yield and Growth Characteristics of Rice in Different Periods. China Rice 2017, 23, 135–138. [Google Scholar]
- Yang, J.; Zhang, J.; Wang, Z.; Zhu, Q.; Liu, L. Activities of enzymes involved in sucrose-to-starch metabolism in rice grains subjected to water stress during filling. Field Crops Res. 2003, 81, 69–81. [Google Scholar] [CrossRef]
- Chen, T.; Xu, G.; Qian, X.; Wang, Z.; Zhang, H.; Yang, J. Post-Anthesis Alternate Wetting and Moderate Soil Drying Irrigation Enhance Gene Expressions of Enzymes Involved in Starch Synthesis in Rice Grains. Sci. Agric. Sin. 2015, 48, 1288–1299. [Google Scholar]
- Cai, Y.; Zhu, Q.; Xu, W.; Wang, W.; Yang, J.; Zhang, Z.; Lang, Y. Effects of Water Stress on the Main Characters of Superior and Inferior Grains Quality and the Properties of RVA Profile during Grain-filling Stage. Acta Agron. Sin. 2004, 30, 241–247. [Google Scholar]
- Jiang, H.; Dian, W.; Wu, P. Effect of high temperature on fine structure of amylopectin in rice endosperm by reducing the activity of the starch branching enzyme. Phytochemistry 2003, 63, 53–59. [Google Scholar] [CrossRef]
- Zhu, Q.; Cao, X.; Luo, Y. Growth Analysis on the Process of Grain Filling in Rice. Acta Agron. Sin. 1988, 14, 182–193. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Treatment | DMT | DMTE | DMTCRV | |||
|---|---|---|---|---|---|---|---|
| StSh | Leaf | StSh | Leaf | StSh | Leaf | ||
| 2016 | A0 | 118.741 a | 53.127 a | 19.526% b | 18.406% b | 16.911% b | 7.566% b |
| A1 | 114.643 a | 46.795 b | 20.671% a | 18.684% b | 24.321% a | 9.927% a | |
| A2 | 116.126 a | 46.954 b | 21.901% a | 20.361% a | 25.852% a | 10.453% a | |
| A3 | 20.965 b | 28.836 c | 3.973% c | 11.497% c | 6.037% c | 8.304% b | |
| B0 | 120.110 a,b | 45.391 b | 19.460% b | 15.568% c | 19.610% b | 7.411% b | |
| B1 | 128.089 a | 50.888 a | 21.127% a | 19.734% a | 25.001% a | 9.932% a | |
| B2 | 116.788 b | 42.980 b | 21.834% a | 17.176% b | 28.607% a | 10.528% a | |
| B3 | 19.324 c | 23.503 c | 3.605% c | 9.059% d | 5.526% c | 6.721% b | |
| 2017 | A0 | 203.680 a | 67.108 a,b | 29.672% b | 29.962% c | 23.468% b | 7.732% b |
| A1 | 200.401 a | 75.450 a | 33.538% a | 35.796% a | 30.591% a | 11.518% a | |
| A2 | 180.365 a | 61.554 b | 33.366% a | 31.692% b | 29.486% a | 10.063% a | |
| A3 | −36.175 b | 37.105 c | −7.592% c | 18.688% d | −8.702% c | 8.926% b | |
| B0 | 203.026 a | 73.076 a | 30.726% b | 29.497% a | 24.522% b | 8.826% a | |
| B1 | 148.879 b | 62.720 b | 25.897% c | 30.176% a | 22.379% b | 9.428% a | |
| B2 | 206.152 a | 41.404 c | 34.856% a | 23.988% b | 34.881% a | 7.006% b | |
| B3 | −72.853 c | 4.395 d | −16.347% d | 2.571% c | −15.847% c | 0.956% c | |
| Year | Treatment | GRmean | GRmax | Tmax (d) | D (d) | ||||
|---|---|---|---|---|---|---|---|---|---|
| SG | IG | SG | IG | SG | IG | SG | IG | ||
| 2016 | A0 | 0.875 a | 0.573 a | 1.363 a | 0.938 a | 12.937 c | 20.460 b | 26.193 b | 30.652 c |
| A1 | 0.696 c | 0.524 b | 1.114 b | 0.837 b | 14.910 b | 25.077 a | 31.229 a | 34.103 b | |
| A2 | 0.729 b | 0.498 c | 1.095 b | 0.766 b | 11.167 d | 20.416 b | 31.045 a | 39.951 a | |
| A3 | 0.786 b | 0.418 d | 1.394 a | 0.664 c | 22.175 a | 25.073 a | 23.378 c | 41.784 a | |
| B0 | 0.949 a | 0.661 a | 1.472 a | 1.067 a | 16.707 b | 25.003 b | 26.878 c | 31.389 c | |
| B1 | 0.816 c | 0.633 a | 1.245 b | 1.031 a | 14.560 d | 26.290 a | 30.445 a | 32.074 c | |
| B2 | 0.799 d | 0.526 b | 1.240 b | 0.814 b | 15.301 c | 23.041 d | 28.695 b | 41.620 b | |
| B3 | 0.844 b | 0.461 c | 1.396 a | 0.698 c | 22.387 a | 24.337 c | 25.163 d | 47.791 a | |
| 2017 | A0 | 0.882 a | 0.570 a | 1.392 a | 0.933 a | 16.334 c | 23.602 b | 25.754 c | 30.946 b |
| A1 | 0.691 c | 0.533 a,b | 1.116 c | 0.861 a,b | 16.256 c | 26.103 a | 31.370 a | 32.840 b | |
| A2 | 0.801 b | 0.502 b | 1.237 b | 0.777 b | 16.641 b | 24.551 a,b | 27.931 b | 38.864 a | |
| A3 | 0.790 b | 0.439 c | 1.386 a | 0.701 b | 20.908 a | 23.893 a,b | 23.301 d | 39.300 a | |
| B0 | 0.929 a | 0.676 a | 1.439 a | 1.100 a | 18.425 c | 27.221 a | 26.902 c | 30.539 d | |
| B1 | 0.815 c | 0.621 b | 1.251 b | 1.007 b | 15.856 d | 27.129 a,b | 30.374 a | 32.639 c | |
| B2 | 0.801 d | 0.522 c | 1.253 b | 0.812 c | 19.616 b | 27.015 b | 28.456 b | 41.206 b | |
| B3 | 0.846 b | 0.458 d | 1.393 a | 0.700 d | 20.271 a | 23.281 c | 25.187 d | 46.443 a | |
| Variety (V) | Treatment (T) | Panicle Length (cm) | PBN | GNPB | SBN | GNSB | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 2016 | 2017 | 2016 | 2017 | 2016 | 2017 | 2016 | 2017 | 2016 | 2017 | ||
| SJ6 | A0 | 14.862 a | 15.054 a | 9.122 a | 9.936 a | 48.020 a | 53.562 a | 12.622 a | 12.787 a | 34.127 a | 38.139 a |
| A1 | 13.561 a | 14.199 a,b | 8.499 a,b | 8.744 b | 44.129 a | 47.345 a | 9.836 b | 9.614 b | 25.350 b | 31.887 b | |
| A2 | 11.511 b | 13.425 b | 8.414 a,b | 9.111 b | 43.321 a | 49.065 a | 8.095 c | 8.019 c | 20.592 c | 26.870 c | |
| A3 | 9.147 c | 11.954 c | 7.532 b | 8.236 b | 38.742 a | 45.774 a | 5.496 d | 6.412 d | 14.334 d | 18.440 d | |
| DN425 | B0 | 16.583 a | 17.679 a | 8.847 a | 8.263 a | 42.195 a | 43.877 a | 11.559 a | 13.297 a | 30.573 a | 36.204 a |
| B1 | 15.095 b | 16.206 a,b | 7.513 b | 7.706 a | 37.462 a,b | 42.236 a | 8.795 b | 10.458 b | 22.846 b | 32.499 a | |
| B2 | 13.971 b | 15.015 b,c | 7.537 b | 7.841 a | 36.216 a,b | 44.101 a | 7.723 b | 8.043 c | 19.716 b | 23.354 b | |
| B3 | 12.309 c | 14.089 c | 6.795 b | 8.296 a | 31.327 b | 44.271 a | 5.665 c | 6.858 c | 12.193 c | 17.358 c | |
| Source | df | Mean square | |||||||||
| V | 1 | 55.691 ** | 8.655 * | 436.912 ** | 0.044 ns | 42.164 ** | |||||
| T | 3 | 38.172 ** | 3.720 ** | 96.045 ** | 90.301 ** | 796.655 ** | |||||
| Y | 1 | 20.998 ** | 2.816 ** | 446.874 ** | 6.086 ** | 380.025 ** | |||||
| V × T | 3 | 0.409 ns | 0.353 ns | 5.463 ns | 0.196 ns | 1.791 ns | |||||
| V × Y | 1 | 0.051 ns | 0.206 ns | 6.200 ns | 3.197 * | 1.861 ns | |||||
| T × Y | 3 | 1.630 ns | 0.589 ns | 26.243 ns | 0.523 ns | 8.172 ns | |||||
| V × T × Y | 3 | 0.696 ns | 0.618 ns | 12.142 ns | 0.498 ns | 4.486 ns | |||||
| Error | 32 | 0.603 | 0.256 | 9.976 | 0.628 | 5.459 | |||||
| Variety (V) | Treatment (T) | EPN (Panicel·m−2) | SPP | Seed Setting Rate | TGW (g) | Theoretical Yield (kg·hm−2) | Actual Yield (kg·hm−2) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2016 | 2017 | 2016 | 2017 | 2016 | 2017 | 2016 | 2017 | 2016 | 2017 | 2016 | 2017 | ||
| SJ6 | A0 | 478.5 a | 473.0 a | 82.147 a | 91.700 a | 96.03% a,b | 93.25% a | 24.246 a | 24.670 a | 9143.151 a | 9951.101 a | 7021.7 a | 8678.9 a |
| A1 | 401.5 a | 451.0 a | 69.478 b | 79.233 b | 97.29% a | 92.78% a | 23.840 a,b | 23.620 b | 6499.060 b | 7825.292 b | 4713.8 b | 6550.9 b | |
| A2 | 423.5 a | 462.0 a | 63.912 b | 75.935 b | 96.04% a,b | 94.61% a | 23.349 a,b | 23.580 b | 6046.034 b | 7815.037 b | 4491.9 b | 6117.1 c | |
| A3 | 412.5 a | 440.0 a | 53.076 c | 64.214 c | 94.77% b | 93.56% a | 22.728 b | 23.080 b | 4706.740 c | 6083.565 c | 3472.5 c | 4157.1 d | |
| DN425 | B0 | 473.0 a | 495.0 a | 72.768 a | 80.081 a | 96.42% a | 94.58% a | 26.911 a | 26.360 a | 8925.619 a | 9949.520 a | 6125.1 a | 8279.4 a |
| B1 | 440.0 a | 473.0 a | 60.308 b | 74.735 a,b | 95.07% a | 90.86% a | 25.385 b | 25.370 b | 6410.777 b | 8107.957 b | 5123.4 b | 6652.7 b | |
| B2 | 434.5 a | 440.0 a | 55.933 b | 67.455 b,c | 95.85% a | 92.02% a | 25.028 b,c | 25.160 b,c | 5828.906 c | 6851.691 c | 4682.5 c | 5910.1 c | |
| B3 | 429.0 a | 429.0 a | 43.520 c | 61.628 c | 97.32% a | 92.34% a | 24.264 c | 24.460 c | 4402.667 d | 5963.277 d | 3496.7 d | 4597.2 d | |
| Source | df | Mean square | |||||||||||
| V | 1 | 958.547 ns | 750.532 ** | 0.000 ns | 35.837 ** | 4.98 × 105 ns | |||||||
| T | 3 | 6146.422 ns | 1411.595 ** | 0.000 ns | 7.596 ** | 3.68 × 107 ** | |||||||
| Y | 1 | 5450.672 ns | 1651.093 ** | 0.120 ** | 0.570 ns | 2.10 × 107 ** | |||||||
| V × T | 3 | 701.422 ns | 11.346 ns | 0.001 * | 0.291 ns | 2.49 × 105 ns | |||||||
| V × Y | 1 | 459.422 ns | 14.854 ns | 0.000 ns | 0.197 ns | 112.218 ns | |||||||
| T × Y | 3 | 625.797 ns | 19.369 ns | 0.000 ns | 0.107 ns | 2.28 × 105 ns | |||||||
| V × T × Y | 3 | 565.297 ns | 13.967 ns | 0.000 ns | 0.191 ns | 1.94 × 105 ns | |||||||
| Error | 32 | 3.80 × 103 | 15.858 | 0.000 | 0.234 | 1.33 × 106 | |||||||
| Yield | DMT | EPN | SPP | TGW | SGRmean | IGRmean | SGD | IGD | |
|---|---|---|---|---|---|---|---|---|---|
| Yield | — | ||||||||
| DMT | 0.806 ** | — | |||||||
| EPN | 0.866 ** | 0.682 ** | — | ||||||
| SPP | 0.906 ** | 0.691 ** | 0.782 ** | — | |||||
| TGW | 0.529 * | 0.403 | 0.514 * | 0.179 | — | ||||
| SGRmean | 0.487 | 0.134 | 0.643 ** | 0.303 | 0.692 ** | — | |||
| IGRmean | 0.815 ** | 0.659 ** | 0.733 ** | 0.574 * | 0.785 ** | 0.522 * | — | ||
| SGD | 0.279 | 0.462 | −0.049 | 0.182 | 0.174 | −0.434 | 0.421 | — | |
| IGD | −0.859 ** | −0.638 ** | −0.798 ** | −0.815 ** | −0.438 | −0.446 | −0.847 ** | −0.235 | — |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Fu, J.; Min, Z.; Zou, D.; Liu, H.; Wang, J.; Zheng, H.; Jia, Y.; Yang, L.; Xin, W.; et al. Response of Rice with Overlapping Growth Stages to Water Stress by Assimilates Accumulation and Transport and Starch Synthesis of Superior and Inferior Grains. Int. J. Mol. Sci. 2022, 23, 11157. https://doi.org/10.3390/ijms231911157
Wang X, Fu J, Min Z, Zou D, Liu H, Wang J, Zheng H, Jia Y, Yang L, Xin W, et al. Response of Rice with Overlapping Growth Stages to Water Stress by Assimilates Accumulation and Transport and Starch Synthesis of Superior and Inferior Grains. International Journal of Molecular Sciences. 2022; 23(19):11157. https://doi.org/10.3390/ijms231911157
Chicago/Turabian StyleWang, Xinpeng, Jinxu Fu, Zhaosen Min, Detang Zou, Hualong Liu, Jingguo Wang, Hongliang Zheng, Yan Jia, Luomiao Yang, Wei Xin, and et al. 2022. "Response of Rice with Overlapping Growth Stages to Water Stress by Assimilates Accumulation and Transport and Starch Synthesis of Superior and Inferior Grains" International Journal of Molecular Sciences 23, no. 19: 11157. https://doi.org/10.3390/ijms231911157
APA StyleWang, X., Fu, J., Min, Z., Zou, D., Liu, H., Wang, J., Zheng, H., Jia, Y., Yang, L., Xin, W., Sun, B., & Zhao, H. (2022). Response of Rice with Overlapping Growth Stages to Water Stress by Assimilates Accumulation and Transport and Starch Synthesis of Superior and Inferior Grains. International Journal of Molecular Sciences, 23(19), 11157. https://doi.org/10.3390/ijms231911157