
Validation of Appropriate Reference Genes for qRT–PCR Normalization in Oat (Avena sativa L.) under UV-B and High-Light Stresses
Abstract
:1. Introduction
2. Results
2.1. Analysis of Primer Specificity and Amplification Efficiency
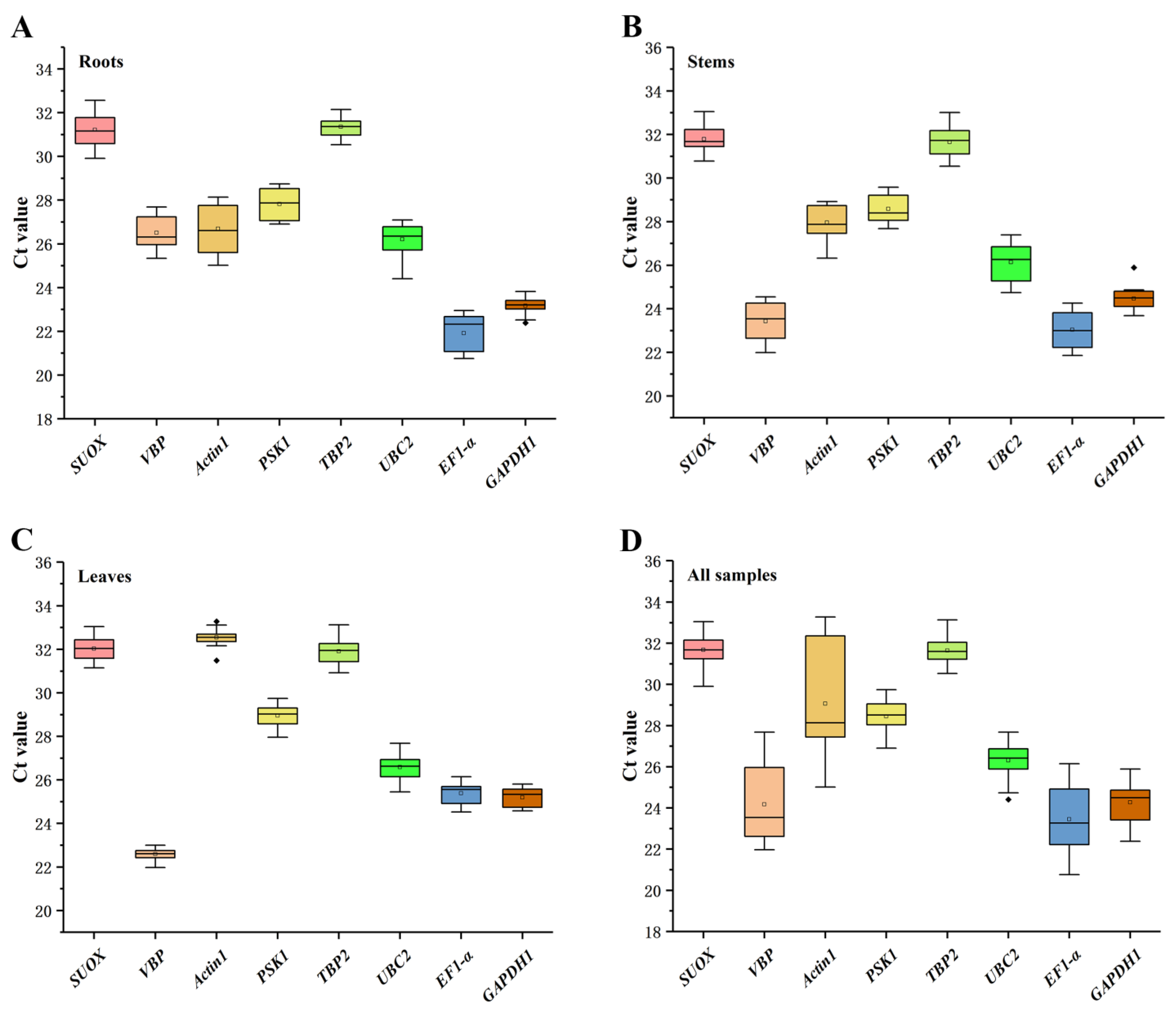
2.2. Expression Profiles and Stability of Candidate Reference Genes
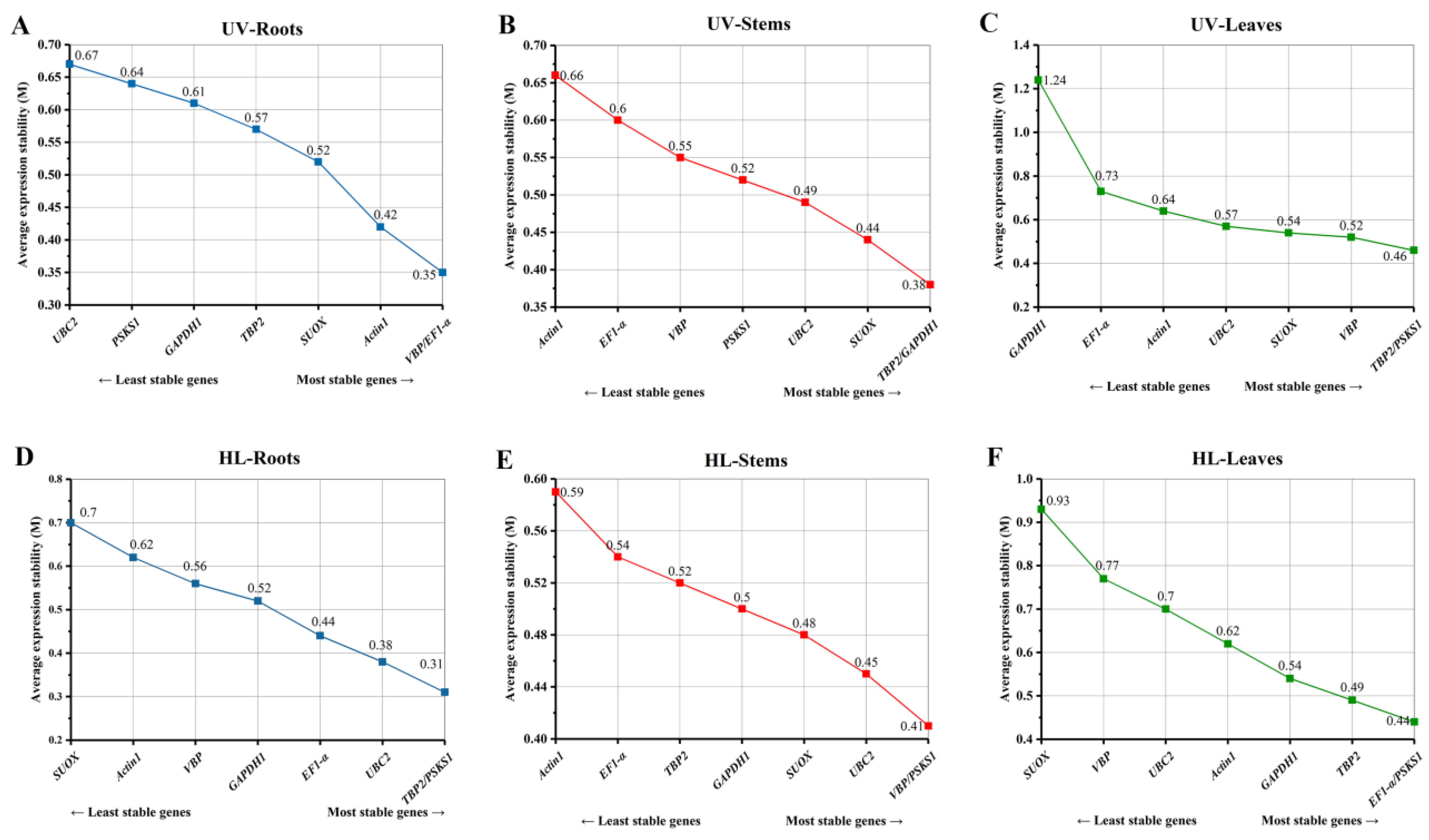
2.3. GeNorm Analysis
2.4. BestKeeper Analysis
2.5. NormFinder Analysis
2.6. RefFinder Analysis
2.7. Reference Gene Validation
3. Discussion
4. Materials and Methods
4.1. Plant and Stress Treatments
4.2. Total RNA Extraction and cDNA Synthesis
4.3. Selection and Primer Design of Candidate Reference Genes
4.4. qRT–PCR Analysis of Candidate Reference Genes
4.5. Algorithms for Evaluating the Stability of Candidate Reference Genes
4.6. Validation of Selected Candidate Reference Genes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Butt, M.S.; Tahir-Nadeem, M.; Khan, M.K.I.; Shabir, R.; Butt, M.S. Oat: Unique among the cereals. Eur. J. Nutr. 2008, 47, 68–79. [Google Scholar] [CrossRef] [PubMed]
- Grundy, M.M.L.; Fardet, A.; Tosh, S.M.; Rich, G.T.; Wilde, P.J. Processing of oat: The impact on oat’s cholesterol lowering effect. Food Funct. 2018, 9, 1328–1343. [Google Scholar] [CrossRef]
- Hu, C.; Sang, S. Triterpenoid Saponins in Oat Bran and Their Levels in Commercial Oat Products. J. Agric. Food Chem. 2020, 68, 6381–6389. [Google Scholar] [CrossRef] [PubMed]
- Winkler, L.R.; Murphy, K.M.; Hermes, J.C. Three hulless oat varieties show economic potential as organic layer feed grain. Renew. Agric. Food Syst. 2018, 33, 418–431. [Google Scholar] [CrossRef]
- Dai, J. Qinghai-Tibet Plateau Climate; China Meteorological Press: Beijing, China, 1990; pp. 22–37. [Google Scholar]
- Zhang, Z.H.; Chang, X.; Su, D.Y.; Yao, R.; Liu, X.D.; Zhu, H.; Liu, G.X.; Zhong, B.J. Comprehensive transcriptome analyses of two Oocystis algae provide insights into the adaptation to Qinghai-Tibet Plateau. J. Syst. Evol. 2021, 59, 1209–1219. [Google Scholar] [CrossRef]
- Huggett, J.; Dheda, K.; Bustin, S.; Zumla, A. Real-time RT-PCR normalisation; strategies and considerations. Genes Immun. 2005, 6, 279–284. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Nolan, T.; Pfaffl, M.W. Quantitative real-time RT-PCR—A perspective. J. Mol. Endocrinol. 2005, 34, 597–601. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper—Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef]
- Udvardi, M.K.; Czechowski, T.; Scheible, W.-R. Eleven golden rules of quantitative RT-PCR. Plant Cell 2008, 20, 1736–1737. [Google Scholar] [CrossRef] [PubMed]
- Czechowski, T.; Stitt, M.; Altmann, T.; Udvardi, M.K.; Scheible, W.R. Genome-wide identification and testing of superior reference genes for transcript normalization in Arabidopsis. Plant Physiol. 2005, 139, 5–17. [Google Scholar] [CrossRef] [Green Version]
- Remans, T.; Smeets, K.; Opdenakker, K.; Mathijsen, D.; Vangronsveld, J.; Cuypers, A. Normalisation of real-time RT-PCR gene expression measurements in Arabidopsis thaliana exposed to increased metal concentrations. Planta 2008, 227, 1343–1349. [Google Scholar] [CrossRef] [PubMed]
- Dudziak, K.; Sozoniuk, M.; Szczerba, H.; Kuzdralinski, A.; Kowalczyk, K.; Boerner, A.; Nowak, M. Identification of stable reference genes for qPCR studies in common wheat (Triticum aestivum L.) seedlings under short-term drought stress. Plant Methods 2020, 16, 58. [Google Scholar] [CrossRef]
- Skiljaica, A.; Jagic, M.; Vuk, T.; Levanic, D.L.; Bauer, N.; Markulin, L. Evaluation of reference genes for RT-qPCR gene expression analysis in Arabidopsis thaliana exposed to elevated temperatures. Plant Biol. 2022, 24, 367–379. [Google Scholar] [CrossRef]
- Verstraeten, B.; De Smet, L.; Kyndt, T.; De Meyer, T. Selection of miRNA reference genes for plant defence studies in rice (Oryza sativa). Planta 2019, 250, 2101–2110. [Google Scholar] [CrossRef]
- Chen, K.; Fessehaie, A.; Arora, R. Selection of Reference Genes for Normalizing Gene Expression During Seed Priming and Germination Using qPCR in Zea mays and Spinacia oleracea. Plant Mol. Biol. Report. 2012, 30, 478–487. [Google Scholar] [CrossRef]
- Abbas, A.; Yu, H.; Li, X.; Cui, H.; Chen, J.; Huang, P. Selection and Validation of Reference Genes for RT-qPCR Analysis in Aegilops tauschii (Coss.) under Different Abiotic Stresses. Int. J. Mol. Sci. 2021, 22, 11017. [Google Scholar] [CrossRef]
- Tajti, J.; Pal, M.; Janda, T. Validation of Reference Genes for Studying Different Abiotic Stresses in Oat (Avena sativa L.) by RT-qPCR. Plants 2021, 10, 1272. [Google Scholar] [CrossRef]
- Duan, Z.L.; Han, W.H.; Yan, L.; Wu, B. Reference gene selections for real time quantitative PCR analysis of gene expression in different oat tissues and under salt stress. Biol. Plant. 2020, 64, 838–844. [Google Scholar] [CrossRef]
- Chen, Y.; Tan, Z.; Hu, B.; Yang, Z.; Xu, B.; Zhuang, L.; Huang, B. Selection and validation of reference genes for target gene analysis with quantitative RT-PCR in leaves and roots of bermudagrass under four different abiotic stresses. Physiol. Plant. 2015, 155, 138–148. [Google Scholar] [CrossRef]
- Shen, J.; Wu, Y.; Jiang, Z.; Xu, Y.; Zheng, T.; Wang, J.; Cheng, T.; Zhang, Q.; Pan, H. Selection and validation of appropriate reference genes for gene expression studies in Forsythia. Physiol. Mol. Biol. Plants 2020, 26, 173–188. [Google Scholar] [CrossRef]
- Xu, M.; Zhang, B.; Su, X.; Zhang, S.; Huang, M. Reference gene selection for quantitative real-time polymerase chain reaction in Populus. Anal. Biochem. 2011, 408, 337–339. [Google Scholar] [CrossRef] [PubMed]
- Costa-Silva, J.; Domingues, D.; Lopes, F.M. RNA-Seq differential expression analysis: An extended review and a software tool. PLoS ONE 2017, 12, e0190152. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Gu, C.; Xuan, L.; Hua, J.; Shi, Q.; Fan, W.; Yin, Y.; Yu, F. Identification of suitable reference genes in Taxodium ’Zhongshanshan’ under abiotic stresses. Trees-Struct. Funct. 2017, 31, 1519–1530. [Google Scholar] [CrossRef]
- Li, X.; Tang, D.; Shi, Y. Selection of reference genes for quantitative real-time PCR normalization in Narcissus pseudonarcissu in different cultivars and different organs. Heliyon 2018, 4, e00686. [Google Scholar] [CrossRef]
- Rapacz, M.; Stepien, A.; Skorupa, K. Internal standards for quantitative RT-PCR studies of gene expression under drought treatment in barley (Hordeum vulgare L.): The effects of developmental stage and leaf age. Acta Physiol. Plant. 2012, 34, 1723–1733. [Google Scholar] [CrossRef]
- Wang, W.T.; Hu, S.Y.; Cao, Y.; Chen, R.; Wang, Z.Z.; Cao, X.Y. Selection and evaluation of reference genes for qRT-PCR of Scutellaria baicalensis Georgi under different experimental conditions. Mol. Biol. Rep. 2021, 48, 1115–1126. [Google Scholar] [CrossRef]
- Zhu, X.L.; Wang, B.Q.; Wang, X.; Wei, X.H. Screening of stable internal reference gene of Quinoa under hormone treatment and abiotic stress. Physiol. Mol. Biol. Plants 2021, 27, 2459–2470. [Google Scholar] [CrossRef]
- Maroufi, A.; Van Bockstaele, E.; De Loose, M. Validation of reference genes for gene expression analysis in chicory (Cichorium intybus) using quantitative real-time PCR. Bmc Mol. Biol. 2010, 11, 15. [Google Scholar] [CrossRef]
- Li, M.-Y.; Song, X.; Wang, F.; Xiong, A.-S. Suitable Reference Genes for Accurate Gene Expression Analysis in Parsley (Petroselinum crispum) for Abiotic Stresses and Hormone Stimuli. Front. Plant Sci. 2016, 7, 1481. [Google Scholar] [CrossRef]
- Rowlands, T.; Baumann, P.; Jackson, S.P. The TATA-binding protein: A general transcription factor in eukaryotes and archaebacteria. Science 1994, 264, 1326–1329. [Google Scholar] [CrossRef]
- Bendjennat, M.; Weil, P.A. The transcriptional repressor activator protein Rap1p is a direct regulator of TATA-binding protein. J. Biol. Chem. 2008, 283, 8699–8710. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Li, X.; Chen, W.; Chen, J.; Lu, W.; Chen, L.; Fu, D. Evaluation of New Reference Genes in Papaya for Accurate Transcript Normalization under Different Experimental Conditions. PLoS ONE 2012, 7, e44405. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xie, W.; Yu, X.; Zhang, Z.; Zhao, Y.; Wang, N.; Wang, Y. Selection of Suitable Reference Genes for RT-qPCR Gene Expression Analysis in Siberian Wild Rye (Elymus sibiricus) under Different Experimental Conditions. Genes 2019, 10, 451. [Google Scholar] [CrossRef] [PubMed]
- Khanlou, K.M.; Van Bockstaele, E. A critique of widely used normalization software tools and an alternative method to identify reliable reference genes in red clover (Trifolium pratense L.). Planta 2012, 236, 1381–1393. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liang, X.; Zhou, X.; Wu, Z.; Yuan, L.; Wang, Y.; Li, Y. Selection of Reference Genes for qRT-PCR Analysis in Medicinal Plant Glycyrrhiza under Abiotic Stresses and Hormonal Treatments. Plants 2020, 9, 1441. [Google Scholar] [CrossRef]
- Maroufi, A. Selection of reference genes for real-time quantitative PCR analysis of gene expression in Glycyrrhiza glabra under drought stress. Biol. Plant. 2016, 60, 645–654. [Google Scholar] [CrossRef]
- Huang, M.X.; Xu, Q.H.; Mitsui, K.; Xu, Z.J. PSK1 regulates expression of SOD1 involved in oxidative stress tolerance in yeast. Fems Microbiol. Lett. 2014, 350, 154–160. [Google Scholar] [CrossRef]
- Hao, Q.; Ren, H.X.; Zhu, J.; Wang, L.S.; Huang, S.C.; Liu, Z.A.; Gao, Z.M.; Shu, Q.Y. Overexpression of PSK1, a SKP1-like gene homologue, from Paeonia suffruticosa, confers salinity tolerance in Arabidopsis. Plant Cell Rep. 2017, 36, 151–162. [Google Scholar] [CrossRef]
- Xu, X.Y.; Huang, M.X.; Ouyang, Y.H.; Iha, H.; Xu, Z.J. PSK1 coordinates glucose metabolism and utilization and regulates energy-metabolism oscillation in Saccharomyces cerevisiae. Yeast 2020, 37, 261–268. [Google Scholar] [CrossRef]
- Sreelakshmi, Y.; Sharma, R. Differential regulation of phenylalanine ammonia lyase activity and protein level by light in tomato seedlings. Plant Physiol. Biochem. 2008, 46, 444–451. [Google Scholar] [CrossRef]
- Weisshaar, B.; Jenkins, G.I. Phenylpropanoid biosynthesis and its regulation. Curr. Opin. Plant Biol. 1998, 1, 251–257. [Google Scholar] [CrossRef]
- Olsen, K.M.; Lea, U.S.; Slimestad, R.; Verheul, M.; Lillo, C. Differential expression of four Arabidopsis PAL genes; PAL1 and PAL2 have functional specialization in abiotic environmental-triggered flavonoid synthesis. J. Plant Physiol. 2008, 165, 1491–1499. [Google Scholar] [CrossRef] [PubMed]
- Shi, R.; Shuford, C.M.; Wang, J.P.; Sun, Y.-H.; Yang, Z.; Chen, H.-C.; Tunlaya-Anukit, S.; Li, Q.; Liu, J.; Muddiman, D.C.; et al. Regulation of phenylalanine ammonia-lyase (PAL) gene family in wood forming tissue of Populus trichocarpa. Planta 2013, 238, 487–497. [Google Scholar] [CrossRef]
- Nag, S.; Kumaria, S. In silico characterization and transcriptional modulation of phenylalanine ammonia lyase (PAL) by abiotic stresses in the medicinal orchid Vanda coerulea Gruff. ex Lindl. Phytochemistry 2018, 156, 176–183. [Google Scholar] [CrossRef]
- Uarrota, V.G.; Maraschin, M.; de Bairros, A.d.F.M.; Pedreschi, R. Factors affecting the capsaicinoid profile of hot peppers and biological activity of their non-pungent analogs (Capsinoids) present in sweet peppers. Crit. Rev. Food Sci. Nutr. 2021, 61, 649–665. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.M.; Zhang, K.M.; Jin, H.H.; Zhu, L.; Li, Y.H. Isolation and expression analysis of four putative structural genes involved in anthocyanin biosynthesis in Begonia semperflorens. J. Hortic. Sci. Biotechnol. 2015, 90, 444–450. [Google Scholar] [CrossRef]
- Li, P.; Ma, F.; Cheng, L. Primary and secondary metabolism in the sun-exposed peel and the shaded peel of apple fruit. Physiol. Plant. 2013, 148, 9–24. [Google Scholar] [CrossRef]
- Levee, V.; Seguin, A. Inducible expression of the heterologous PAL2 promoter from bean in white pine (Pinus strobus) transgenic cells. Tree Physiol. 2001, 21, 665–672. [Google Scholar] [CrossRef]
- Rudus, I.; Kepczynski, J. Reference gene selection for molecular studies of dormancy in wild oat (Avena fatua L.) caryopses by RT-qPCR method. PLoS ONE 2018, 13, e0192343. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034.0031. [Google Scholar] [CrossRef] [Green Version]
- Andersen, C.L.; Jensen, J.L.; Orntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.; Xiao, P.; Chen, D.; Xu, L.; Zhang, B. miRDeepFinder: A miRNA analysis tool for deep sequencing of plant small RNAs. Plant Mol. Biol. 2012, 80, 75–84. [Google Scholar] [CrossRef]
- Yang, J.; Han, F.; Yang, L.; Wang, J.; Jin, F.; Luo, A.; Zhao, F. Identification of Reference Genes for RT-qPCR Analysis in Gleditsia microphylla under Abiotic Stress and Hormone Treatment. Genes 2022, 13, 1227. [Google Scholar] [CrossRef]
- Tu, C.; Xu, P.; Han, R.; Luo, J.; Xu, L. Defining Suitable Reference Genes for qRT-PCR in Plagiodera versicolora (Coleoptera: Chrysomelidae) under Different Biotic or Abiotic Conditions. Agronomy 2022, 12, 1192. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Gene Name | Primers (Forward/Reverse, 5′-3′) | Tm (°C) | Amplicon Length (bp) | qRT–PCR Efficiency (%) | R² |
|---|---|---|---|---|---|---|
| SUOX | sulfite oxidase | GAGCCTCATCGATCTGCGTT | 80.08 | 132 | 103.42 | 0.9935 |
| ATTCACGAGGCCACTGATGG | ||||||
| VBP | victorin binding protein | CCGTTACCTGCACAAGTTGC | 81.80 | 117 | 109.03 | 0.9971 |
| GGATCGGTGACAGGCATCAT | ||||||
| Actin1 | actin-encoding | GGTATGGTCAAGGCCGGATT | 87.07 | 111 | 101.53 | 0.9993 |
| ATCCTTCTGTCCCATCCCGA | ||||||
| PSKS1 | protein PSK SIMULATOR 1-like | TGGAGAGTGGGCAAATACAGG | 81.06 | 136 | 92.61 | 0.9998 |
| AAGCCACACAACAAGCTCAAG | ||||||
| TBP2 | TATA-binding protein 2-like | GTGGATCTGTTGAGGCACCC | 82.88 | 123 | 94.26 | 0.9930 |
| ATACTCTGCATTGCGCGCTT | ||||||
| UBC2 | ubiquitin-conjugating enzyme E2 | ACAGCGTTCCGAGAGTTGTT | 82.51 | 104 | 109.42 | 0.9629 |
| AAGAGTCAGCCCGAATGCAG | ||||||
| EF1-α | elongation factor 1-alpha | AGCCTGGTATGGTTGTGACC | 84.73 | 147 | 109.21 | 0.9947 |
| GCTTGAGATCCTTCACGGCA | ||||||
| GAPDH1 | glyceraldehyde-3-phosphate dehydrogenase 1 | TTCTTCCTGAGTTGAACGGC | 82.17 | 102 | 98.14 | 0.9970 |
| ATGCAGCCTTCTCGATTCTG |
| Rank | UV-B | High-Light | All Samples | ||||
|---|---|---|---|---|---|---|---|
| Roots | Stems | Leaves | Roots | Stems | Leaves | ||
| 1 | SUOX (0.33) | UBC2 (0.31) | SUOX (0.39) | PSKS1 (0.37) | UBC2 (0.37) | EF1-α (0.47) | UBC2 (0.67) |
| 2 | TBP2 (0.36) | PSKS1 (0.58) | UBC2 (0.51) | UBC2 (0.41) | SUOX (0.38) | PSKS1 (0.49) | PSKS1 (0.67) |
| 3 | PSKS1 (0.36) | SUOX (0.61) | TBP2 (0.58) | EF1-α (0.46) | TBP2 (0.52) | TBP2 (0.54) | TBP2 (0.74) |
| 4 | GAPDH1 (0.43) | GAPDH1 (0.63) | VBP (0.58) | Actin1 (0.50) | EF1-α (0.55) | Actin1 (0.58) | SUOX (0.96) |
| 5 | VBP (0.49) | TBP2 (0.67) | PSKS1 (0.59) | TBP2 (0.53) | GAPDH1 (0.58) | GAPDH1 (0.66) | EF1-α (1.09) |
| 6 | UBC2 (0.49) | VBP (0.86) | Actin1 (0.83) | GAPDH1 (0.60) | PSKS1 (0.62) | UBC2 (0.84) | GAPDH1 (1.16) |
| 7 | EF1-α (0.50) | EF1-α (0.93) | EF1-α (1.09) | VBP (0.74) | VBP (0.65) | VBP (0.94) | VBP (1.69) |
| 8 | Actin1 (0.64) | Actin1 (1.03) | GAPDH1 (2.02) | SUOX (0.81) | Actin1 (0.78) | SUOX (0.98) | Actin1 (2.28) |
| Rank | UV-B | High-Light | All Samples | ||||
|---|---|---|---|---|---|---|---|
| Roots | Stems | Leaves | Roots | Stems | Leaves | ||
| 1 | EF1-α (0.26) | TBP2 (0.23) | VBP (0.09) | PSKS1 (0.25) | UBC2 (0.28) | PSKS1 (0.17) | PSKS1 (0.31) |
| 2 | VBP (0.29) | GAPDH1 (0.27) | UBC2 (0.35) | TBP2 (0.27) | GAPDH1 (0.38) | EF1-α (0.38) | EF1-α (0.47) |
| 3 | SUOX (0.41) | SUOX (0.34) | PSKS1 (0.4) | EF1-α (0.32) | TBP2 (0.38) | TBP2 (0.39) | TBP2 (0.71) |
| 4 | TBP2 (0.45) | PSKS1 (0.48) | TBP2 (0.44) | UBC2 (0.42) | EF1-α (0.39) | GAPDH1 (0.43) | UBC2 (0.83) |
| 5 | GAPDH1 (0.55) | VBP (0.49) | SUOX (0.46) | GAPDH1 (0.49) | SUOX (0.40) | UBC2 (0.66) | SUOX (1.31) |
| 6 | Actin1 (0.58) | UBC2 (0.56) | Actin1 (0.78) | VBP (0.52) | VBP (0.42) | Actin1 (0.80) | GAPDH1 (1.35) |
| 7 | PSKS1 (0.61) | EF1-α (0.56) | EF1-α (0.81) | Actin1 (0.72) | PSKS1 (0.42) | VBP (0.81) | Actin1 (2.27) |
| 8 | UBC2 (0.62) | Actin1 (0.71) | GAPDH1 (2.73) | SUOX (0.85) | Actin1 (0.65) | SUOX (1.30) | VBP (2.40) |
| Rank | UV-B | High-Light | All Samples | ||||
|---|---|---|---|---|---|---|---|
| Roots | Stems | Leaves | Roots | Stems | Leaves | ||
| 1 | EF1-α (1.63) | TBP2 (1.5) | PSKS1 (1.97) | PSKS1 (1) | UBC2 (1.32) | PSKS1 (1.19) | PSKS1 (1.19) |
| 2 | VBP (2.11) | GAPDH1 (2) | VBP (2.21) | TBP2 (2.11) | GAPDH1 (3.16) | EF1-α (1.41) | UBC2 (2) |
| 3 | SUOX (2.45) | SUOX (3) | TBP2 (2.45) | UBC2 (3.13) | SUOX (3.56) | TBP2 (3) | EF1-α (2.99) |
| 4 | TBP2 (3.56) | UBC2 (3.46) | UBC2 (2.99) | EF1-α (3.22) | TBP2 (3.57) | GAPDH1 (4.23) | TBP2 (3) |
| 5 | Actin1 (5.18) | PSKS1 (3.56) | SUOX (3.16) | GAPDH1 (5.23) | VBP (3.98) | Actin1 (5.18) | SUOX (4.73) |
| 6 | GAPDH1 (5.18) | VBP (5.48) | Actin1 (6) | Actin1 (6.09) | PSKS1 (4.14) | UBC2 (5.48) | GAPDH1 (6) |
| 7 | PSKS1 (5.66) | EF1-α (7) | EF1-α (7) | VBP (6.24) | EF1-α (4.87) | VBP (7) | Actin1 (7.24) |
| 8 | UBC2 (7.44) | Actin1 (8) | GAPDH1 (8) | SUOX (8) | Actin1 (8) | SUOX (8) | VBP (7.74) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yin, H.; Yin, D.; Zhang, M.; Gao, Z.; Tuluhong, M.; Li, X.; Li, J.; Li, B.; Cui, G. Validation of Appropriate Reference Genes for qRT–PCR Normalization in Oat (Avena sativa L.) under UV-B and High-Light Stresses. Int. J. Mol. Sci. 2022, 23, 11187. https://doi.org/10.3390/ijms231911187
Yin H, Yin D, Zhang M, Gao Z, Tuluhong M, Li X, Li J, Li B, Cui G. Validation of Appropriate Reference Genes for qRT–PCR Normalization in Oat (Avena sativa L.) under UV-B and High-Light Stresses. International Journal of Molecular Sciences. 2022; 23(19):11187. https://doi.org/10.3390/ijms231911187
Chicago/Turabian StyleYin, Hang, Danni Yin, Mingzhi Zhang, Zhiqiang Gao, Muzhapaer Tuluhong, Xiaoming Li, Jikai Li, Bing Li, and Guowen Cui. 2022. "Validation of Appropriate Reference Genes for qRT–PCR Normalization in Oat (Avena sativa L.) under UV-B and High-Light Stresses" International Journal of Molecular Sciences 23, no. 19: 11187. https://doi.org/10.3390/ijms231911187
APA StyleYin, H., Yin, D., Zhang, M., Gao, Z., Tuluhong, M., Li, X., Li, J., Li, B., & Cui, G. (2022). Validation of Appropriate Reference Genes for qRT–PCR Normalization in Oat (Avena sativa L.) under UV-B and High-Light Stresses. International Journal of Molecular Sciences, 23(19), 11187. https://doi.org/10.3390/ijms231911187