
A Screening of Epigenetic Therapeutic Targets for Non-Small Cell Lung Cancer Reveals PADI4 and KDM6B as Promising Candidates

 , , , ,
, , , ,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. Evaluation of Epigenetic Targets in Non-Small Cell Lung Cancers Using Publicly Available Data
2.2. Systematic Literature Review Analysis of Potential Epigenetic Targets
2.3. In Silico Analysis of Expression of the 4 Target Genes in Lung Cancer Cells
2.4. Epigenetic Targets Gene Expression by Real-Time PCR
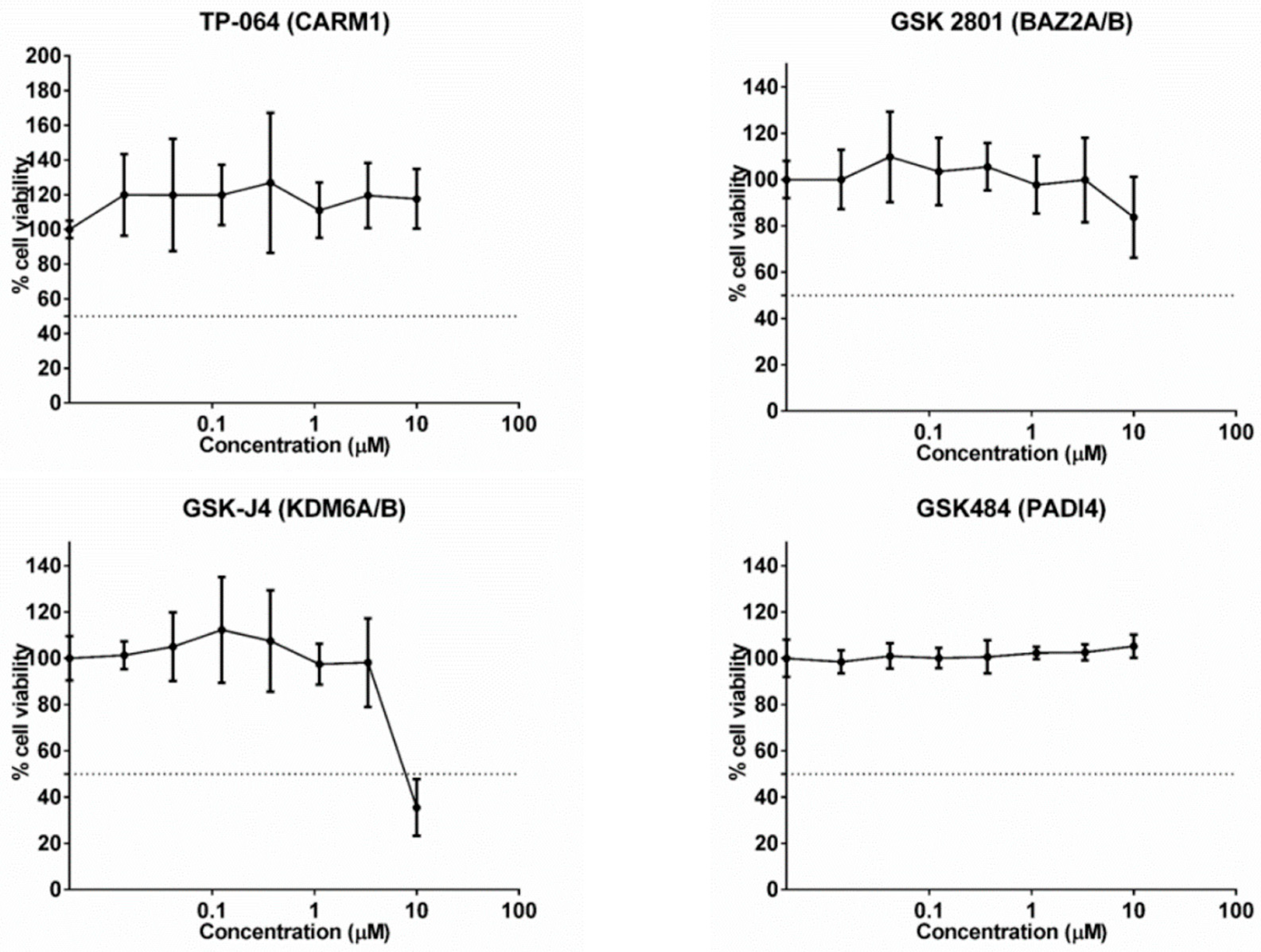
2.5. Cytotoxic Potential (IC50) and Determination of the Maximum Concentration without Cytotoxic Effect of Epigenetic Molecules
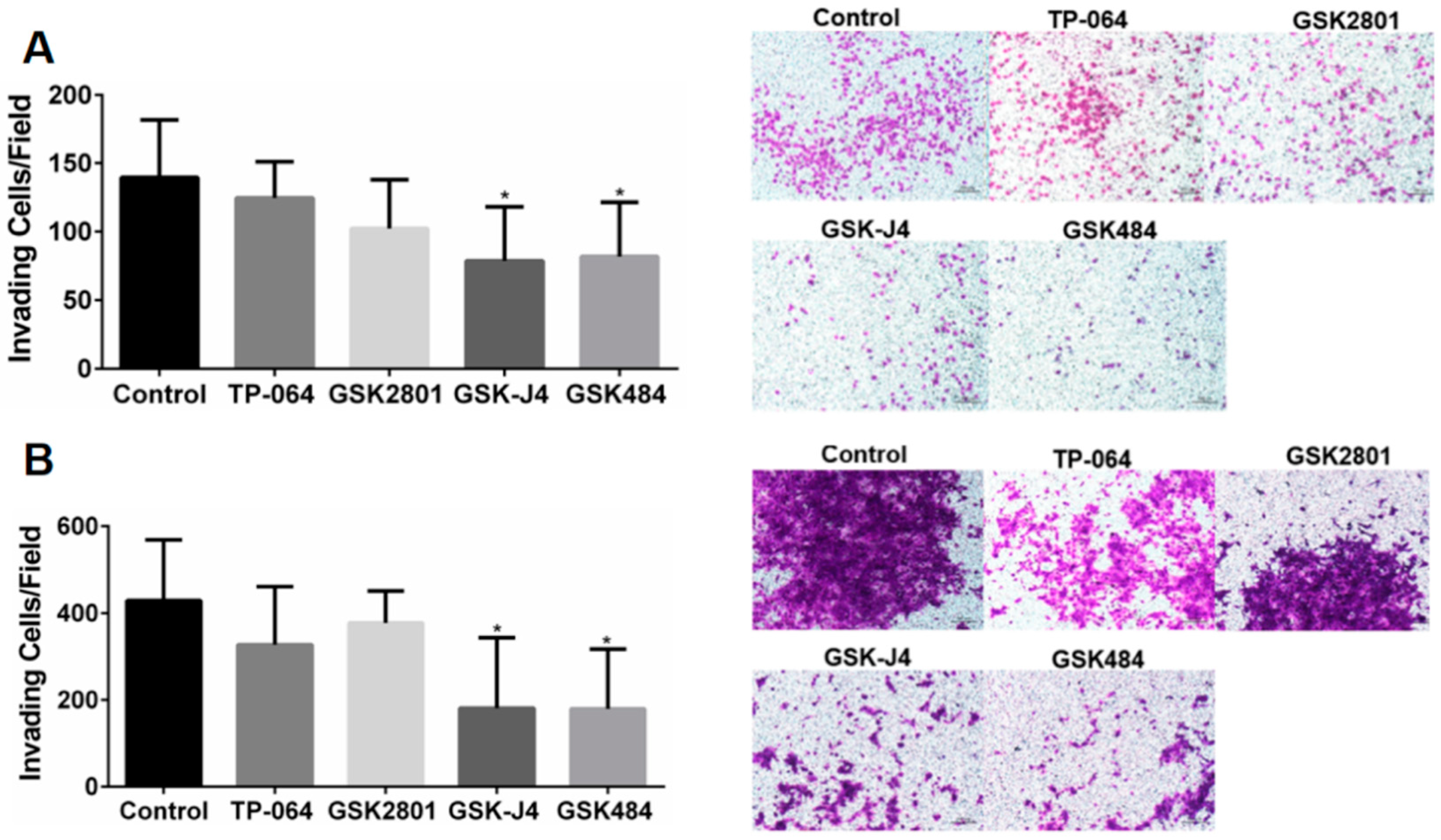
2.6. Epigenetic Inhibitor Effects on Cancer Cell Migration and Invasiveness
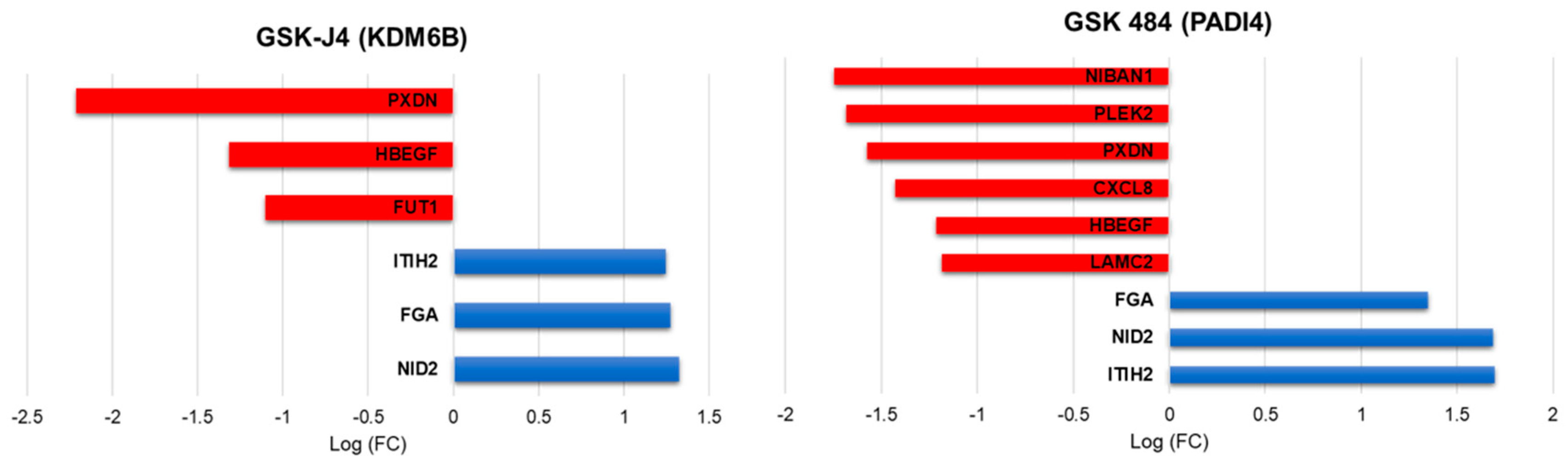
2.7. Global Gene Expression Changes upon Treatment with Chemical Probes for KDM6B and PADI4
3. Discussion
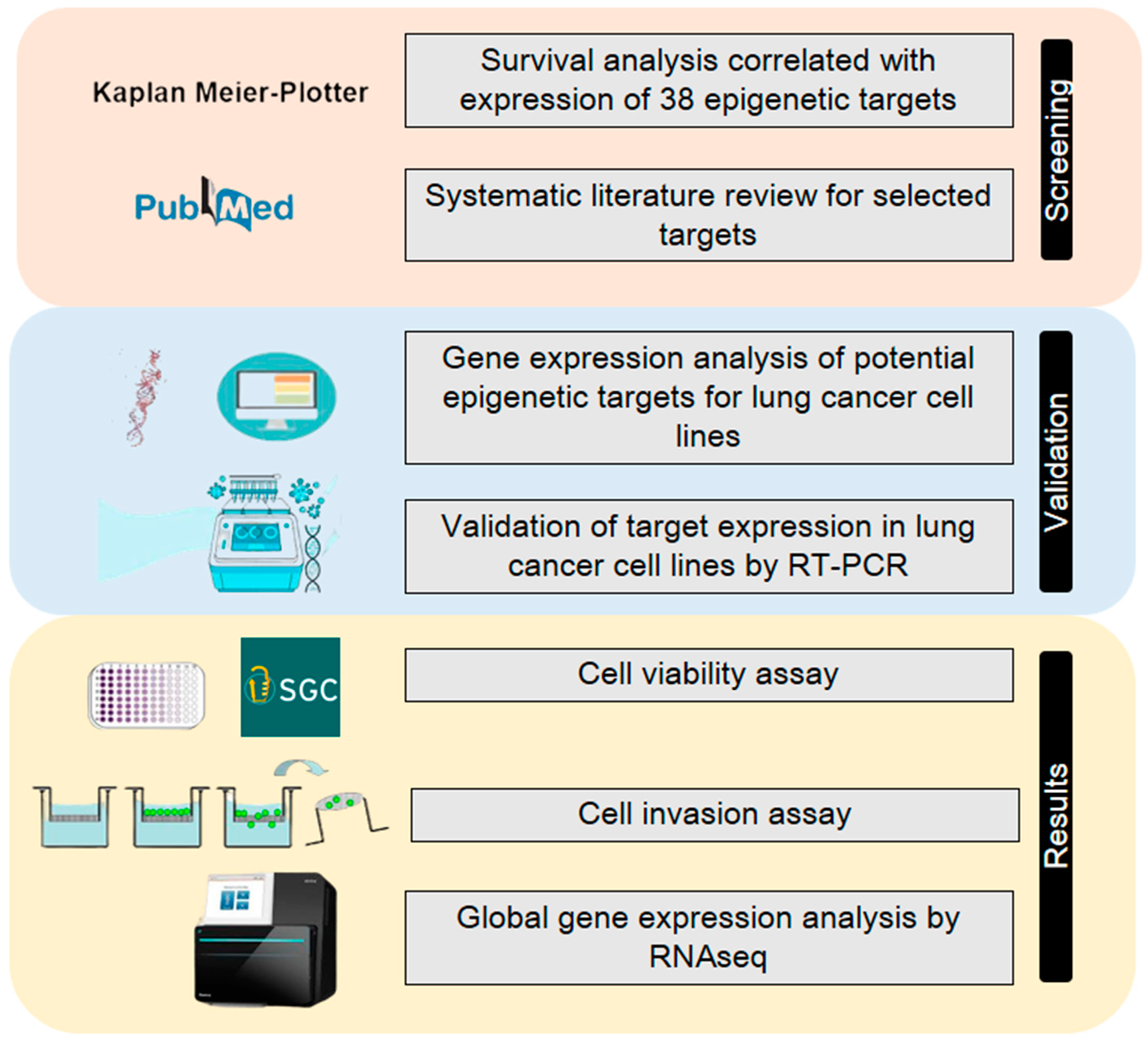
4. Materials and Methods
4.1. In Silico Analysis for Determination of Potential Epigenetic Targets in Patients with Non-Small Cell Lung Cancer
4.2. Systematic Literature Review for Selection of New Epigenetic Targets for NSCLC
4.3. In Silico Analysis of Epigenetic Target Expression
4.4. Cell Culture
4.5. RNA Isolation and Reverse Transcription-Quantitative Polymerase Chain Reaction (RT-qPCR)
4.6. Epigenetic Probes Cell Viability Assay
4.7. Invasion Assay
4.8. RNAseq Data Generation
4.9. Alignment and Differential Expression
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA. Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Devarakonda, S.; Morgensztern, D.; Govindan, R. Genomic alterations in lung adenocarcinoma. Lancet Oncol. 2015, 16, e342–e351. [Google Scholar] [CrossRef] [PubMed]
- Ansari, J.; Shackelford, R.E.; El-Ostal, H. Epigenetics in non-small cell lung cancer: From basics to therapeutics. Transl. Lung Cancer Res. 2016, 5, 155–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardtstock, F.; Myers, D.; Li, T.; Cizova, D.; Maywald, U.; Wilke, T.; Griesinger, F. Real-world treatment and survival of patients with advanced non-small cell lung Cancer: A German retrospective data analysis. BMC Cancer 2020, 20, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Niu, F.Y.; Zhou, Q.; Yang, J.J.; Zhong, W.Z.; Chen, Z.H.; Deng, W.; He, Y.Y.; Chen, H.J.; Zeng, Z.; Ke, E.E.; et al. Distribution and prognosis of uncommon metastases from non-small cell lung cancer. BMC Cancer 2016, 16, 1. [Google Scholar] [CrossRef] [Green Version]
- Hildebrandt, M.A.T.; Gu, J.; Wu, X. Pharmacogenomics of platinum-based chemotherapy in NSCLC. Expert Opin. Drug Metab. Toxicol. 2009, 5, 745–755. [Google Scholar] [CrossRef] [Green Version]
- Yuan, M.; Huang, L.L.; Chen, J.H.; Wu, J.; Xu, Q. The emerging treatment landscape of targeted therapy in non-small-cell lung cancer. Signal Transduct. Target. Ther. 2019, 4, 61. [Google Scholar] [CrossRef] [Green Version]
- Bajbouj, K.; Al-ali, A.; Ramakrishnan, R.K.; Saber-ayad, M.; Hamid, Q. Histone modification in nsclc: Molecular mechanisms and therapeutic targets. Int. J. Mol. Sci. 2021, 22, 11701. [Google Scholar] [CrossRef]
- Audia, J.E.; Campbell, R.M. Histone modifications and cancer. Cold Spring Harb. Perspect. Biol. 2016, 8, 1–31. [Google Scholar] [CrossRef]
- Yu, X.; Li, M.; Guo, C.; Wu, Y.; Zhao, L.; Shi, Q.; Song, J.; Song, B. Therapeutic Targeting of Cancer: Epigenetic Homeostasis. Front. Oncol. 2021, 11, 4471. [Google Scholar] [CrossRef]
- Müller, S.; Ackloo, S.; Arrowsmith, C.H.; Bauser, M.; Baryza, J.L.; Blagg, J.; Böttcher, J.; Bountra, C.; Brown, P.J.; Bunnage, M.E.; et al. Donated chemical probes for open science. Elife 2018, 7, e34311. [Google Scholar] [CrossRef] [PubMed]
- Ackloo, S.; Brown, P.J.; Müller, S. Chemical probes targeting epigenetic proteins: Applications beyond oncology. Epigenetics 2017, 12, 378–400. [Google Scholar] [CrossRef] [PubMed]
- Brown, P.J.; Müller, S. Open access chemical probes for epigenetic targets. Future Med. Chem. 2015, 7, 1901–1917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Q.; Heidenreich, D.; Zhou, S.; Ackloo, S.; Krämer, A.; Nakka, K.; Lima-Fernandes, E.; Deblois, G.; Duan, S.; Vellanki, R.N.; et al. A chemical toolbox for the study of bromodomains and epigenetic signaling. Nat. Commun. 2019, 10, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huston, A.; Arrowsmith, C.H.; Knapp, S.; Schapira, M. Probing the epigenome. Nat. Chem. Biol. 2015, 11, 542–545. [Google Scholar] [CrossRef]
- Wu, Q.; Schapira, M.; Arrowsmith, C.H.; Barsyte-Lovejoy, D. Protein arginine methylation: From enigmatic functions to therapeutic targeting. Nat. Rev. Drug Discov. 2021, 20, 509–530. [Google Scholar] [CrossRef]
- Nervi, C.; De Marinis, E.; Codacci-Pisanelli, G. Epigenetic treatment of solid tumours: A review of clinical trials. Clin. Epigenetics 2015, 7, 127. [Google Scholar] [CrossRef] [Green Version]
- Pachmayr, E.; Treese, C.; Stein, U. Underlying Mechanisms for Distant Metastasis—Molecular Biology. Visc. Med. 2017, 33, 11–20. [Google Scholar] [CrossRef]
- Zhai, Q.; Wang, L.; Zhao, P.; Li, T. Role of citrullination modification catalyzed by peptidylarginine deiminase 4 in gene transcriptional regulation. Acta Biochim. Biophys. Sin. 2017, 49, 567–572. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Qu, Y.; Teng, X.; Xing, Y.; Li, D.; Li, C.; Cai, L. PADI4-mediated epithelial-mesenchymal transition in lung cancer cells. Mol. Med. Rep. 2019, 19, 3087–3094. [Google Scholar] [CrossRef]
- Ramadoss, S.; Chen, X.; Wang, C.Y. Histone demethylase KDM6B promotes epithelial-mesenchymal transition. J. Biol. Chem. 2012, 287, 44508–44517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xun, J.; Gao, R.; Wang, B.; Li, Y.; Ma, Y.; Guan, J.; Zhang, Q. Histone demethylase KDM6B inhibits breast cancer metastasis by regulating Wnt/β-catenin signaling. FEBS Open Bio 2021, 11, 2273–2281. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-H.; Kim, O.; Kim, H.-J.; Hwangbo, C.; Lee, J.-H. Epigenetic regulation of TGF-β-induced EMT by JMJD3/KDM6B histone H3K27 demethylase. Oncogenesis 2021, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Li, F.; Gao, B.; Ma, M.; Chen, M.; Wu, Y.; Zhang, W.; Sun, Y.; Liu, S.; Shen, H. KDM6B-mediated histone demethylation of LDHA promotes lung metastasis of osteosarcoma. Theranostics 2021, 11, 3868–3881. [Google Scholar] [CrossRef]
- Jose, A.; Shenoy, G.G.; Rodrigues, G.S.; Kumar, N.A.N.; Munisamy, M.; Thomas, L.; Kolesar, J.; Rai, G.; Rao, P.P.N.; Rao, M. Histone demethylase KDM5B as a therapeutic target for cancer therapy. Cancers 2020, 12, 2121. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, G.; Zhang, Y.; Cui, X.; Wang, S.; Gao, S.; Wang, Y.; Liu, Y.; Bae, J.H.; Yang, W.H.; et al. Fibrinogen Alpha Chain Knockout Promotes Tumor Growth and Metastasis through Integrin-AKT Signaling Pathway in Lung Cancer. Mol. Cancer Res. 2020, 18, 943–954. [Google Scholar] [CrossRef] [Green Version]
- Chai, A.W.Y.; Cheung, A.K.L.; Dai, W.; Ko, J.M.Y.; Ip, J.C.Y.; Chan, K.W.; Kwong, D.L.W.; Ng, W.T.; Lee, A.W.M.; Ngan, R.K.C.; et al. Metastasis-suppressing NID2, an epigenetically-silenced gene, in the pathogenesis of nasopharyngeal carcinoma and esophageal squamous cell carcinoma. Oncotarget 2016, 7, 78859. [Google Scholar] [CrossRef] [Green Version]
- Hamm, A.; Veeck, J.; Bektas, N.; Wild, P.J.; Hartmann, A.; Heindrichs, U.; Kristiansen, G.; Werbowetski-Ogilvie, T.; Del Maestro, R.; Knuechel, R.; et al. Frequent expression loss of Inter-alpha-trypsin inhibitor heavy chain (ITIH) genes in multiple human solid tumors: A systematic expression analysis. BMC Cancer 2008, 8, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.Z.; Liang, L. High expression of PXDN is associated with poor prognosis and promotes proliferation, invasion as well as migration in ovarian cancer. Ann. Diagn. Pathol. 2018, 34, 161–165. [Google Scholar] [CrossRef]
- Ongusaha, P.P.; Kwak, J.C.; Zwible, A.J.; Macip, S.; Higashiyama, S.; Taniguchi, N.; Fang, L.; Lee, S.W. HB-EGF is a potent inducer of tumor growth and angiogenesis. Cancer Res. 2004, 64, 5283–5290. [Google Scholar] [CrossRef]
- Moon, Y.W.; Rao, G.; Kim, J.J.; Shim, H.S.; Park, K.S.; An, S.S.; Kim, B.; Steeg, P.S.; Sarfaraz, S.; Changwoo Lee, L.; et al. LAMC2 enhances the metastatic potential of lung adenocarcinoma. Cell Death Differ. 2015, 22, 1341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, X.; Liao, X.; Qiu, S.; Xu, H.; Zhang, S.; Wang, S.; Ai, J.; Yang, L. CXCL8 in Tumor Biology and Its Implications for Clinical Translation. Front. Mol. Biosci. 2022, 9, 235. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Zhou, X.-M.; Yang, F.-F.; Zhang, Q.; Miao, Y.; Hou, G. FAM129A promotes invasion and proliferation by activating FAK signaling pathway in non-small cell lung cancer. Int. J. Clin. Exp. Pathol. 2019, 12, 893. [Google Scholar] [PubMed]
- Cai, T.; Yao, W.; Qiu, L.; Zhu, A.R.; Shi, Z.; Du, Y. PLEK2 promotes the proliferation and migration of non-small cell lung cancer cells in a BRD4-dependent manner. Mol. Biol. Rep. 2022, 1–12, 3693–3704. [Google Scholar] [CrossRef]
- Park, S.; Lim, J.M.; Chun, J.N.; Lee, S.; Kim, T.M.; Kim, D.W.; Kim, S.Y.; Bae, D.J.; Bae, S.M.; So, I.; et al. Altered expression of fucosylation pathway genes is associated with poor prognosis and tumor metastasis in non-small cell lung cancer. Int. J. Oncol. 2020, 56, 559. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Chan, Y.T.; Tan, H.Y.; Li, S.; Wang, N.; Feng, Y. Epigenetic regulation in human cancer: The potential role of epi-drug in cancer therapy. Mol. Cancer 2020, 19, 1–16. [Google Scholar] [CrossRef]
- De Marchi, P.; Berardinelli, G.N.; Cavagna, R.D.O.; Pinto, I.A.; Da Silva, F.A.F.; Duval Da Silva, V.; Santana, I.V.V.; Da Silva, E.C.A.; Ferro Leal, L.; Reis, R.M. Microsatellite Instability Is Rare in the Admixed Brazilian Population of Non-Small Cell Lung Cancer: A Cohort of 526 Cases. Pathobiology 2022, 89, 101–106. [Google Scholar] [CrossRef]
- De Marchi, P.; Leal, L.F.; Duval Da Silva, V.; Da Silva, E.C.A.; Cordeiro De Lima, V.C.; Reis, R.M. PD-L1 expression by Tumor Proportion Score (TPS) and Combined Positive Score (CPS) are similar in non-small cell lung cancer (NSCLC). J. Clin. Pathol. 2021, 74, 735–740. [Google Scholar] [CrossRef]
- Bodor, J.N.; Boumber, Y.; Borghaei, H. Biomarkers for immune checkpoint inhibition in non-small cell lung cancer (NSCLC). Cancer 2020, 126, 260–270. [Google Scholar] [CrossRef]
- Rohrbach, A.S.; Slade, D.J.; Thompson, P.R.; Mowen, K.A. Activation of PAD4 in NET formation. Front. Immunol. 2012, 3, 360. [Google Scholar] [CrossRef]
- Shi, L.; Yao, H.; Liu, Z.; Xu, M.; Tsung, A.; Wang, Y. Endogenous PAD4 in Breast Cancer Cells Mediates Cancer Extracellular Chromatin Network Formation and Promotes Lung Metastasis. Mol. Cancer Res. 2020, 18, 735–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, T.; Schutte, A.; Jimenez, L.; Gonçalves, A.N.A.; Keller, A.; Pipkin, M.E.; Nakaya, H.I.; Pereira, R.M.; Martinez, G.J. Kdm6b Regulates the Generation of Effector CD8 + T Cells by Inducing Chromatin Accessibility in Effector-Associated Genes. J. Immunol. 2021, 206, 2170–2183. [Google Scholar] [CrossRef] [PubMed]
- Gyorffy, B.; Surowiak, P.; Budczies, J.; Lánczky, A. Online survival analysis software to assess the prognostic value of biomarkers using transcriptomic data in non-small-cell lung cancer. PLoS ONE 2013, 8, e82241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.F.; Lee, C.Y.; Lai, L.C.; Tsai, M.H.; Lu, T.P.; Chuang, E.Y. CellExpress: A comprehensive microarray-based cancer cell line and clinical sample gene expression analysis online system. Database J. Biol. Databases Curation 2018, 2018, bax101. [Google Scholar] [CrossRef] [PubMed]
- Fukumasu, H.; Rochetti, A.L.; Pires, P.R.L.; Silva, E.R.; Mesquita, L.G.; Strefezzi, R.F.; De Carvalho, D.D.; Dagli, M.L. Constitutive androstane receptor ligands modulate the anti-tumor efficacy of paclitaxel in non-small cell lung cancer cells. PLoS ONE 2014, 9, e99484. [Google Scholar] [CrossRef] [Green Version]
- Bourne, R. ImageJ. Fundam. Digit. Imaging Med. 2010, 9, 185–188. [Google Scholar] [CrossRef]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinf. 2012, 13, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Vallone, P.M.; Butler, J.M. AutoDimer: A screening tool for primer-dimer and hairpin structures. Biotechniques 2004, 37, 226–231. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Alcalde, F.; Okonechnikov, K.; Carbonell, J.; Cruz, L.M.; Götz, S.; Tarazona, S.; Dopazo, J.; Meyer, T.F.; Conesa, A. Qualimap: Evaluating next-generation sequencing alignment data. Bioinformatics 2012, 28, 2678–2679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, L.J.; Kuhn, M.; Stark, M.; Chaffron, S.; Creevey, C.; Muller, J.; Doerks, T.; Julien, P.; Roth, A.; Simonovic, M.; et al. STRING 8--a global view on proteins and their functional interactions in 630 organisms. Nucleic Acids Res. 2009, 37, D412–D416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Epigenetic Targets | Enzyme Class | p Value | Hazard Ratio (HR) | Low Expression Survival (Median in Months) | High Expression Survival (Median in Months) |
|---|---|---|---|---|---|
| PRMT1 | Methyltransferase | 5.8 × 10−9 | (HR = 3.22; IC = 95%; 2.12–4.88) | 75 | 21 |
| KDM6B | Demethylase | 6.3 × 10−9 | (HR = 2.81; IC = 95%; 1.95–4.03) | 150 | 34 |
| CARM1 | Methyltransferase | 9.2 × 10−8 | (HR = 2.73; IC = 95%; 1.86–4.00) | 48 | 18 |
| BAZ2A | Bromodomain | 7.8 × 10−6 | (HR = 2.23; IC = 95%; 1.56–3.20) | 175 | 52 |
| BRD4 | Bromodomain | 0.0025 | (HR = 1.81; IC = 95%; 1.23–2.68) | 117 | 69 |
| EZH2 | Methyltransferase | 0.024 | (HR = 1.50; IC = 95%; 1.05–2.13) | 126 | 70 |
| PADI4 | Deiminase | 0.025 | (HR = 1.47; IC = 95%; 1.05–2.06) | 107 | 80 |
| BRD9 | Bromodomain | 0.033 | (HR = 1. 45; IC = 95%; 1.03–2.05) | 103 | 52 |
| Clinical Data | n | Freq. (%) |
|---|---|---|
| Histology | ||
| Adenocarcinoma | 590 | −100% |
| Stage | ||
| 1 | 277 | (46.95%) |
| 2 | 115 | (19.49 %) |
| 3 | 16 | (2.71 %) |
| 4 | 4 | (0.68%) |
| Staging | ||
| T1 | 123 | (20.85%) |
| T2 | 103 | (17.46%) |
| T3 | 4 | (0.68%) |
| T4 | 0 | 0% |
| N0 | 184 | (31.19%) |
| N1 | 42 | (7.12%) |
| N2 | 3 | (0.51%) |
| M0 | 229 | (38.81%) |
| M1 | 1 | (0.17%) |
| Gender | ||
| Women | 247 | (41.86%) |
| Men | 289 | (48.98%) |
| Smoking history | ||
| Exclude those never smoked | 180 | (30.51%) |
| Only those never smoked | 92 | (15.59%) |
| Surgery success | ||
| Only surgical margins negative | 127 | (21.53%) |
| Chemotherapy | ||
| Yes | 14 | (2.37%) |
| No | 8 | (1.36%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lesbon, J.C.C.; Garnica, T.K.; Xavier, P.L.P.; Rochetti, A.L.; Reis, R.M.; Müller, S.; Fukumasu, H. A Screening of Epigenetic Therapeutic Targets for Non-Small Cell Lung Cancer Reveals PADI4 and KDM6B as Promising Candidates. Int. J. Mol. Sci. 2022, 23, 11911. https://doi.org/10.3390/ijms231911911
Lesbon JCC, Garnica TK, Xavier PLP, Rochetti AL, Reis RM, Müller S, Fukumasu H. A Screening of Epigenetic Therapeutic Targets for Non-Small Cell Lung Cancer Reveals PADI4 and KDM6B as Promising Candidates. International Journal of Molecular Sciences. 2022; 23(19):11911. https://doi.org/10.3390/ijms231911911
Chicago/Turabian StyleLesbon, Jéssika Cristina Chagas, Taismara Kustro Garnica, Pedro Luiz Porfírio Xavier, Arina Lázaro Rochetti, Rui Manuel Reis, Susanne Müller, and Heidge Fukumasu. 2022. "A Screening of Epigenetic Therapeutic Targets for Non-Small Cell Lung Cancer Reveals PADI4 and KDM6B as Promising Candidates" International Journal of Molecular Sciences 23, no. 19: 11911. https://doi.org/10.3390/ijms231911911
APA StyleLesbon, J. C. C., Garnica, T. K., Xavier, P. L. P., Rochetti, A. L., Reis, R. M., Müller, S., & Fukumasu, H. (2022). A Screening of Epigenetic Therapeutic Targets for Non-Small Cell Lung Cancer Reveals PADI4 and KDM6B as Promising Candidates. International Journal of Molecular Sciences, 23(19), 11911. https://doi.org/10.3390/ijms231911911