
Diflubenzuron Induces Cardiotoxicity in Zebrafish Embryos

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
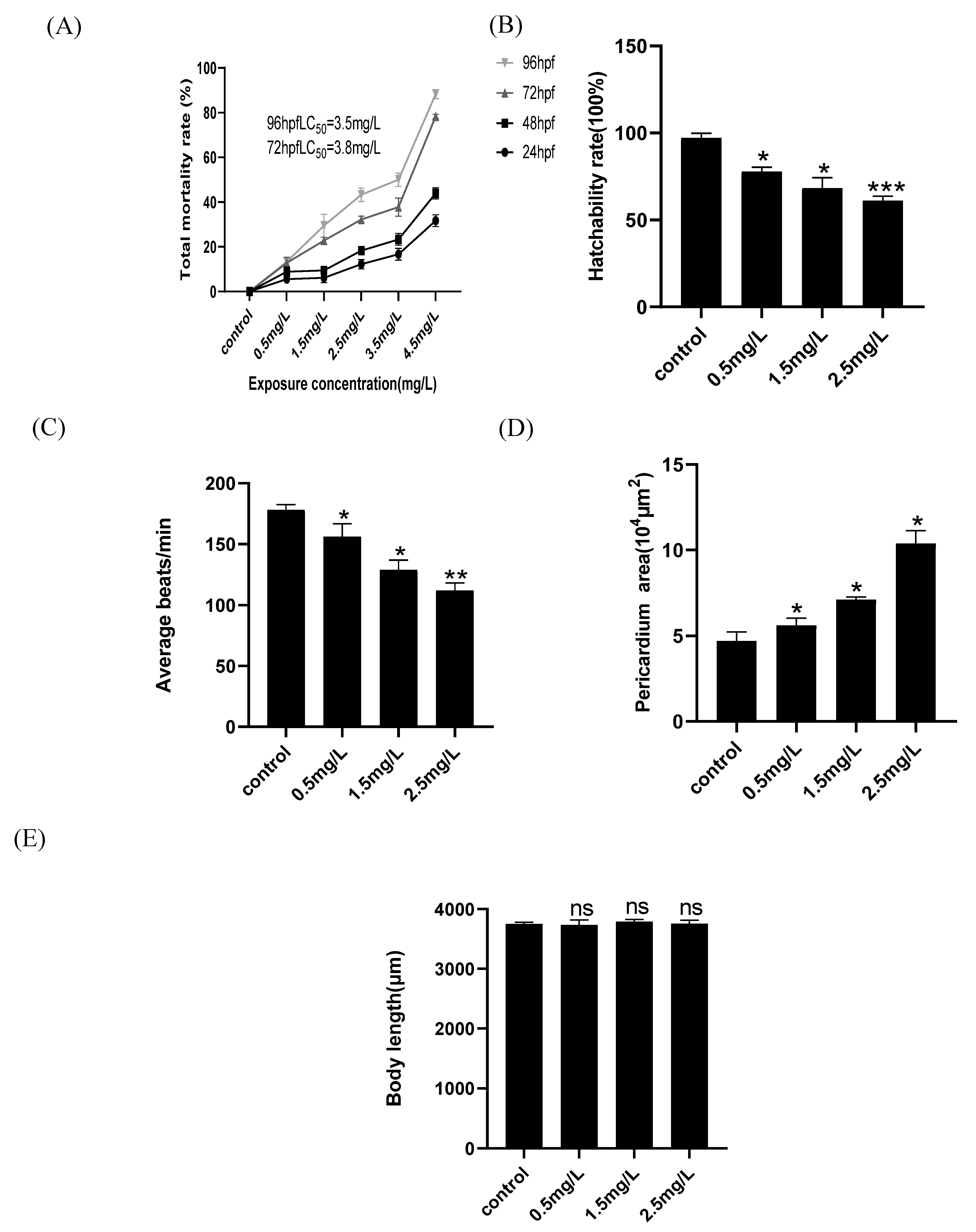
2.1. Developmental Toxicity of Zebrafish Embryos Is Induced by Diflubenzuron
2.2. Diflubenzuron Exposure Causes the Cardiovascular Injure in Zebrafish Embryos
2.3. Diflubenzuron Exposure Inhibits the Expression of Cardiac Development Genes
2.4. Diflubenzuron Exposure Induces Oxidative Stress in Zebrafish Embryos
2.5. Diflubenzuron Exposure Leads to Heart Apoptosis
3. Discussion
4. Materials and Methods
4.1. Zebrafish Husbandry and Experimental Reagents
4.2. Diflubenzuron Exposure and Morphological Changes of Zebrafish Embryos
4.3. Real-Time Quantitative PCR
4.4. Hemotoxin and Eosin (HE) Staining
4.5. Oxidative Stress Analysis
4.6. Acridine Orange (AO) Staining
4.7. Immunofluorescence
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Eisler, R. Diflubenzuron Hazards to Fish, Wildlife and Invertebrates: A Synoptic Review; Biological Report; U.S. Fish and Wildlife Service: Washington, DC, USA, 1992; Volume 4, pp. 1–36.
- Jonsson, C.M.; Silva, M.S.G.M.; Macedo, V.S.; De Dantzger, D.D.; Vallim, J.H.; Marigo, A.L.S.; Aoyama, H. Prediction of a low-risk concentration of diflubenzuron to aquatic organisms and evaluation of clay and gravel in reducing the toxicity. Panam. J. Aquat. Sci. 2015, 10, 259–272. [Google Scholar]
- Branson, E.J.; Ronsberg, S.S.; Ritchie, G. Efficacy of teflubenzuron (Calicide®) for the treatment of sea lice, Lepeophtheirus salmonis (Kroyer 1838), infestations of farmed Atlantic salmon (Salmo salar L.). Aquac. Res. 2000, 31, 861–867. [Google Scholar] [CrossRef]
- Li, M.; Liu, X.; Feng, X. Cardiovascular toxicity and anxiety-like behavior induced by deltamethrin in zebrafish (Danio rerio) larvae. Chemosphere 2019, 219, 155–164. [Google Scholar] [CrossRef]
- Ma, J.; Huang, Y.; Jiang, P.; Liu, Z.; Luo, Q.; Zhong, K.Y.; Yuan, W.; Meng, Y.L.; Lu, H.Q. Pyridaben induced cardiotoxicity during the looping stages of zebrafish (Danio rerio) embryos. Aquat. Toxicol. 2021, 237, 105870. [Google Scholar] [CrossRef]
- Samuelsen, O.B.; Lunestad, B.T.; Hannisdal, R.; Bannister, R.; Olsen, S.; Tjensvoll, T.; Farestveit, E.; Ervik, A. Distribution and persistence of the anti sea-lice drug teflubenzuron in wild fauna and sediments around a salmon farm, following a standard treatment. Sci. Total Environ. 2015, 508, 115–121. [Google Scholar] [CrossRef] [PubMed]
- United States Environmental Protection Agency, Office of Pesticide Programs, Special Review and Reregistration Division. Reregistration Eligibility Decision (RED) Diflubenzuron; 19 August 1997; EPA-738-F-97-008. Available online: https://nepis.epa.gov/Adobe/PDF/20000RXG.pdf (accessed on 17 September 2022).
- Fischer, S.A.; Hall, L.W. Environmental concentrations and aquatic toxicity data ondiflubenzuron (Dimilin®). Crit. Rev. Toxicol. 1992, 22, 45–79. [Google Scholar] [CrossRef] [PubMed]
- Hormazábal, V.; Yndestad, M. High-Performance Liquid Chromatographic Determination of Teflubenzuron and Diflubenzuron in Fish Tissues of Farmed Fish for Residue Studies. J. Liq. Chromatogr. Relat. Technol. 1996, 19, 2603–2614. [Google Scholar] [CrossRef]
- Thisse, C.; Zon, L. Organogenesis–Heart and blood formation from the zebrafish point of view. Science 2002, 295, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Bakkers, J. Zebrafish as a model to study cardiac development and human cardiac disease. Cardiovasc. Res. 2011, 91, 279–288. [Google Scholar] [CrossRef] [Green Version]
- Alzualde, A.; Behl, M.; Sipes, N.S.; Hsieh, J.H.; Alday, A.; Tice, R.R.; Paules, R.S.; Muriana, A.; Quevedo, C. Toxicity profiling of flame retardants in zebrafish embryos using a battery of assays for developmental toxicity, neurotoxicity, cardiotoxicity and hepatotoxicity toward human relevance. Neurotoxicol. Teratol. 2018, 70, 40–50. [Google Scholar] [CrossRef]
- Reimers, M.J.; Hahn, M.E.; Tanguay, R.L. Two zebrafish alcohol dehydrogenases share common ancestry with mammalian class I, II, IV, and V alcohol dehydrogenase genes but have distinct functional characteristics. J. Biol. Chem. 2004, 279, 38303–38312. [Google Scholar] [CrossRef] [PubMed]
- Zetterberg, H.; Campbell, W.A.; Hong, W.Y.; Xia, W. The cytosolic loop of the secretase component presenilin enhancer 2 protects zebrafish embryos from apop-tosis. J. Biol. Chem. 2006, 281, 11933–11939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lieschke, G.J.; Currie, P.D. Animal models of human disease: Zebrafish swim into view. Nat. Rev. Genet. 2007, 8, 353–367. [Google Scholar] [CrossRef] [PubMed]
- Zon, L.I.; Peterson, R.T. In vivo drug discovery in the zebrafish. Nat. Rev. Drug Discov. 2005, 4, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Huang, Y.; Xiao, J.; Cao, H.; Peng, Y.; Chen, Z.; Liu, F.; Wang, H.; Liao, X.; Lu, H. Exposure to diclofop-methyl induces cardiac developmental toxicity in zebrafish embryos. Environ. Pollut. 2020, 259, 113926. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Ma, J.; Meng, Y.; Wei, Y.; Xie, S.; Jiang, P.; Wang, Z.; Chen, X.; Liu, Z.; Zhong, K.; et al. Exposure to Oxadiazon-Butachlor causes cardiac toxicity in zebrafish embryos. Environ. Pollut. 2020, 265, 114775. [Google Scholar] [CrossRef]
- Shu, T.; Chi, N.C. Zebrafish models in cardiac development and congenital heart birth defects. Differentiation 2012, 84, 4–16. [Google Scholar]
- Ma, J.Z.; Huang, Y.; Peng, Y.Y.; Xu, Z.P.; Wang, Z.Q.; Chen, X.B.; Xie, S.L.; Jiang, P.; Zhong, K.Y.; Lu, H.Q. Bifenazate exposure induces cardiotoxicity in zebrafish embryos. Environ. Pollut. 2021, 274, 116539. [Google Scholar] [CrossRef] [PubMed]
- Jia, K.; Cheng, B.; Huang, L.; Xiao, J.H.; Bai, Z.H.; Liao, X.J.; Cao, Z.G.; Shen, T.Z.; Zhang, C.P.; Hu, C.Y.; et al. Thiophanate-methyl induces severe hepatotoxicity in zebrafish. Chemosphere 2020, 248, 125941. [Google Scholar] [CrossRef]
- Zhong, K.; Meng, Y.; Wu, J.; Wei, Y.; Huang, Y.; Ma, J.; Lu, H. Effect of flupyradifurone on zebrafish embryonic development. Environ. Pollut. 2021, 285, 117323. [Google Scholar] [CrossRef] [PubMed]
- Weigt, S.; Huebler, N.; Braunbeck, T. Zebrafish teratogenicity test with metabolic activation (mDarT): Effects of phase I activation of acetaminophen on zebrafish Danio rerio embryos. Toxicology 2010, 275, 36–49. [Google Scholar] [CrossRef]
- Hajdú, J.; Csapó, Z.; Rigó, J., Jr.; Marton, T.; Pete, B.; Papp, Z. A ductus venosus veleszuletett hianyanak sulyos szivfejlodesi rendellenesseggel tarsult formaja - esetismertetes es az irodalom attekintese. Orv. Hetil. 2006, 147, 1901–1903. [Google Scholar] [PubMed]
- Lawson, N.D.; Weinstein, B.M. In vivo imaging of embryonic vascular development using transgenic zebrafish. Dev. Biol. 2002, 248, 307–318. [Google Scholar] [CrossRef] [Green Version]
- Alenzi, F.Q.; El-Nashar, E.M.; Al-Ghandi, S.S.; Abbas, M.Y.; Hamad, A.M.; El-Saeed, O.M.; Wyse, R.K.H.; Lotfy, M. Investigation of Bcl-2 and PCNA in hepatocellular carcinoma: Relation to chronic HCVJ. Egypt. Nat. Cancer Inst. 2010, 22, 87–94. [Google Scholar]
- Granados-Riveron, J.T.; Ghosh, T.K.; Pope, M.; Bu’Lock, F.; Thornborough, C.; Eason, J.; Kirk, E.P.; Fatkin, D.; Feneley, M.P.; Harvey, R.P.; et al. α-Cardiac myosin heavy chain (MYH6) mutations affecting myofibril formation are associated with congenital heart defects. Hum. Mol. Genet. 2010, 19, 4007–4016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.; Zhang, J.; Qian, J.; Hu, C. Cardiotoxicity evaluation of anthracyclines in zebrafish (Danio rerio). J. Appl. Toxicol. 2015, 35, 241–252. [Google Scholar] [CrossRef] [PubMed]
- Grassini, D.; Lagendijk, A.K.; Angelis, J.; Da Silva, J.; Jeanes, A.; Zettler, N.; Bower, N.; Hogan, B.; Smith, K. Nppa and Nppb act redundantly during zebrafish cardiac development to confine AVC marker expression and reduce cardiac jelly volume. Development 2018, 145, 160739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zennaro, C.; Mariotti, M.; Carraro, S.; Pasqualetti, A.; Corbelli, S.; Armelloni, M.; Li, M.; Ikehata, M.; Clai, M.; Artero, P.; et al. Podocyte developmental defects caused by adriamycin in zebrafish embryos and larvae: A novel model of glomerular damage. PLoS ONE 2014, 9, e98131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houweling, A.C.; van Borren, M.M.; Moorman, A.F.M.; Christoffels, V.M. Expression and regulation of the atrial natriuretic factor encoding gene Nppa during development and disease. Cardiovasc. Res. 2005, 67, 583–593. [Google Scholar] [CrossRef] [Green Version]
- Guo, S.; Zhang, Y.; Zhou, T.; Wang, D.; Weng, Y.; Chen, Q.; Ma, J.; Li, Y.P.; Wang, L. GATA4 as a novel regulator involved in the development of the neural crest and craniofacial skeleton via Barx1. Cell Death Differ. 2018, 25, 1996–2009. [Google Scholar] [CrossRef] [PubMed]
- Steed, E.; Faggianelli, N.; Roth, S.; Ramspacher, C.; Concordet, J.P.; Vermot, J. Klf2a couples mechanotransduction and zebrafish valve morphogenesis through fibronectin synthesis. Nat. Commun. 2016, 7, 11646. [Google Scholar] [CrossRef] [PubMed]
- Chi, N.C.; Shaw, R.M.; De Val, S.; Kang, G.; Jan, L.Y.; Black, B.L.; Stainier, D.Y.R. Foxn4 directly regulatestbx2b expression and atrioventricular canal formation. Genes Dev. 2008, 22, 734–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alok, S.; Pachori, L.; Custer, D.H.; Shannon, C.; Erica, K.; John, K. Bone morphogenetic protein 4 mediates myocardial ischemic injury through JNK-dependent signaling pathway. J. Mol. Cell. Cardiol. 2010, 48, 1255–1265. [Google Scholar]
- Nostro, M.C.; Cheng, X.; Keller, G.M.; Gadue, P. Wnt, activin, and BMP signaling regulate distinct stages in the developmental pathway from embryonic stem cells to blood. Cell Stem Cell 2008, 2, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Rivkees, S.A. Programmed cell death in the developing heart: Regulation by BMP4 and FGF2. Dev. Dyn. 2000, 217, 388–400. [Google Scholar] [CrossRef]
- Liu, H.; Chu, T.; Chen, L.; Gui, W.; Zhu, G. The cardiovascular toxicity of triadimefon in early life stage of zebrafish and potential implications to human health. Environ. Pollut. 2017, 231, 1093–1103. [Google Scholar] [CrossRef]
- Hu, W.; Xin, Y.; Hu, J.; Sun, Y.; Zhao, Y. Inhibitor of DNA binding in heart development and cardiovascular diseases. Cell Commun. Signal. 2019, 24, 17–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.H.; Li, X.F.; Yang, G.L.; Weng, H.B.; Wang, X.Q.; Wang, Q. Changes of enzyme activity and gene expression in embryonic zebrafish co-exposed to beta-cypermethrin and thiacloprid. Environ. Pollut. 2020, 256, 113437. [Google Scholar] [CrossRef]
- Li, Y.; Zhu, H.; Lei, X.Q.; Zhang, H.J.; Guan, C.W.; Chen, Z.R.; Zheng, W.; Xu, H.; Tian, Y.; Yu, Z.M.; et al. The first evidence of deinoxanthin from Deinococcus sp. Y35 with strong algicidal effect on the toxic dinoflagellate Alexandrium tamarense. J. Hazard. Mater. 2015, 290, 87–95. [Google Scholar]
- Faggio, C.; Pagano, M.; Alampi, R.; Vazzana, I.; Felice, M.R. Cytotoxicity, hae-molymphatic parameters, and oxidative stress following exposure to sub-lethal concentrations of quaternium-15 in Mytilus galloprovincialis. Aquat. Toxicol. 2016, 180, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Jiang, C.; Wu, Z.Q.; Gong, Y.X.; Wang, G.X. Toxic effects of three stro-bilurins (trifloxystrobin, azoxystrobin and kresoxim-methyl) on mRNA expression and antioxidant enzymes in grass carp (Ctenopharyngodon idella) juveniles. Ecotoxicol. Environ. Saf. 2013, 98, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Gobi, N.; Vaseeharan, B.; Rekha, R.; Vijayakumar, S.; Faggio, C. Bio-accumulation, cytotoxicity and oxidative stress of the acute exposure selenium inOreochromis mossambicus. Ecotoxicol. Environ. Saf. 2018, 162, 147–159. [Google Scholar] [CrossRef] [PubMed]
- Freitas, R.; Silvestro, S.; Coppola, F.; Meucci, V.; Battaglia, F.; Intorre, L.; Soares, A.M.V.M.; Pretti, C.; Faggio, C. Biochemical and physiological re-sponses induced in Mytilus galloprovincialis after a chronic exposure to salicylic acid. Aquat. Toxicol. 2019, 214, 105258. [Google Scholar] [CrossRef] [PubMed]
- Ge, W.; Yan, S.; Wang, J.; Zhu, L.; Chen, A.; Wang, J. Oxidative stress and DNA damage induced by imidacloprid in zebrafish (Danio rerio). Agric. Food Chem. 2015, 63, 1856–1862. [Google Scholar] [CrossRef] [PubMed]
- Burgos-Aceves, M.A.; Cohen, A.; Paolella, G.; Lepretti, M.; Smith, Y.; Faggio, C.; Lionetti, L. Modulation of mitochondrial functions by xenobiotic-induced microRNA: From environmental sentinel organisms to mammals. Sci. Total Environ. 2018, 645, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Tian, S.N.; Teng, M.M.; Meng, Z.Y.; Yan, S.; Jia, M.; Li, R.S.; Liu, L.; Yan, J.; Zhou, Z.Q.; Zhu, W.T. Toxicity effects in zebrafish embryos (Danio rerio) induced by prothioconazole. Environ. Pollut. 2019, 255, 113269. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Dong, F.; Liu, X.; Xu, J.; Wu, X.; Liu, W.; Zheng, Y. Crosstalk of oxidative damage, apoptosis, and autophagy under endoplasmic reticulum (ER) stress involved in thifluzamide-induced liver damage in zebrafish (Danio rerio). Environ. Pollut. 2018, 243, 1904–1911. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Li, X.; Liu, X.; Yang, G.; An, X.; Wang, Q.; Wang, Y. Joint toxic effects of triazophos and imidacloprid on zebrafish (Danio rerio). Environ. Pollut. 2018, 235, 470–481. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, X.; Xu, X.; Yu, T.; Li, M.; Liu, Y.; Lai, J.; Mao, H.; Hu, C.; Wang, S. Diflubenzuron Induces Cardiotoxicity in Zebrafish Embryos. Int. J. Mol. Sci. 2022, 23, 11932. https://doi.org/10.3390/ijms231911932
Han X, Xu X, Yu T, Li M, Liu Y, Lai J, Mao H, Hu C, Wang S. Diflubenzuron Induces Cardiotoxicity in Zebrafish Embryos. International Journal of Molecular Sciences. 2022; 23(19):11932. https://doi.org/10.3390/ijms231911932
Chicago/Turabian StyleHan, Xue, Xiaowen Xu, Tingting Yu, Meifeng Li, Yulong Liu, Jingli Lai, Huiling Mao, Chengyu Hu, and Shanghong Wang. 2022. "Diflubenzuron Induces Cardiotoxicity in Zebrafish Embryos" International Journal of Molecular Sciences 23, no. 19: 11932. https://doi.org/10.3390/ijms231911932
APA StyleHan, X., Xu, X., Yu, T., Li, M., Liu, Y., Lai, J., Mao, H., Hu, C., & Wang, S. (2022). Diflubenzuron Induces Cardiotoxicity in Zebrafish Embryos. International Journal of Molecular Sciences, 23(19), 11932. https://doi.org/10.3390/ijms231911932