
Genome-Wide Identification, Characterization, and Expression Analysis of Tubby-like Protein (TLP) Gene Family Members in Woodland Strawberry (Fragaria vesca)

Abstract
:1. Introduction
2. Results
2.1. Identification of TLP Gene Family Members in F. vesca
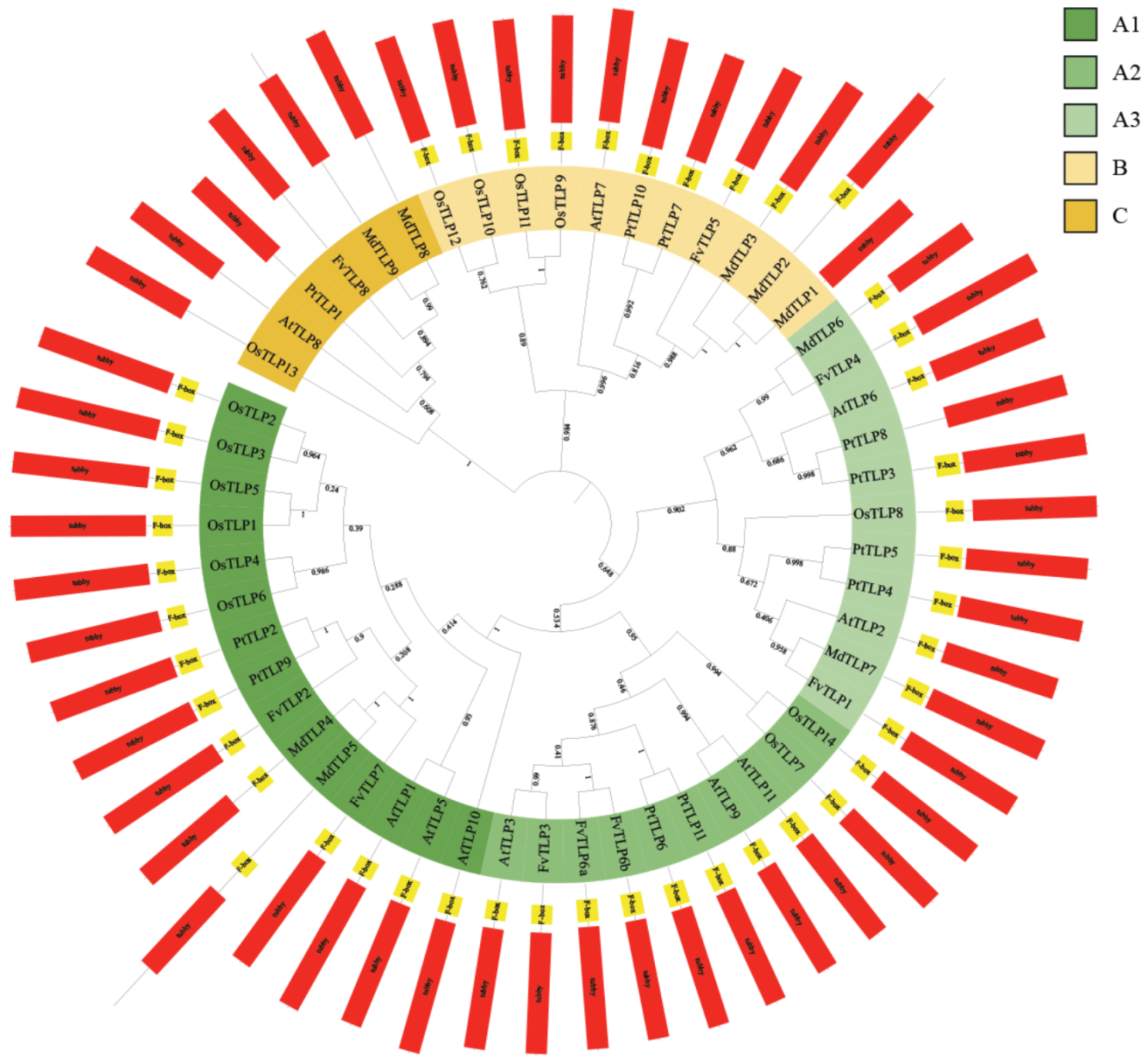
2.2. Phylogenetic and Gene Structure Analysis
2.3. Cis-Element and Promoter Analysis
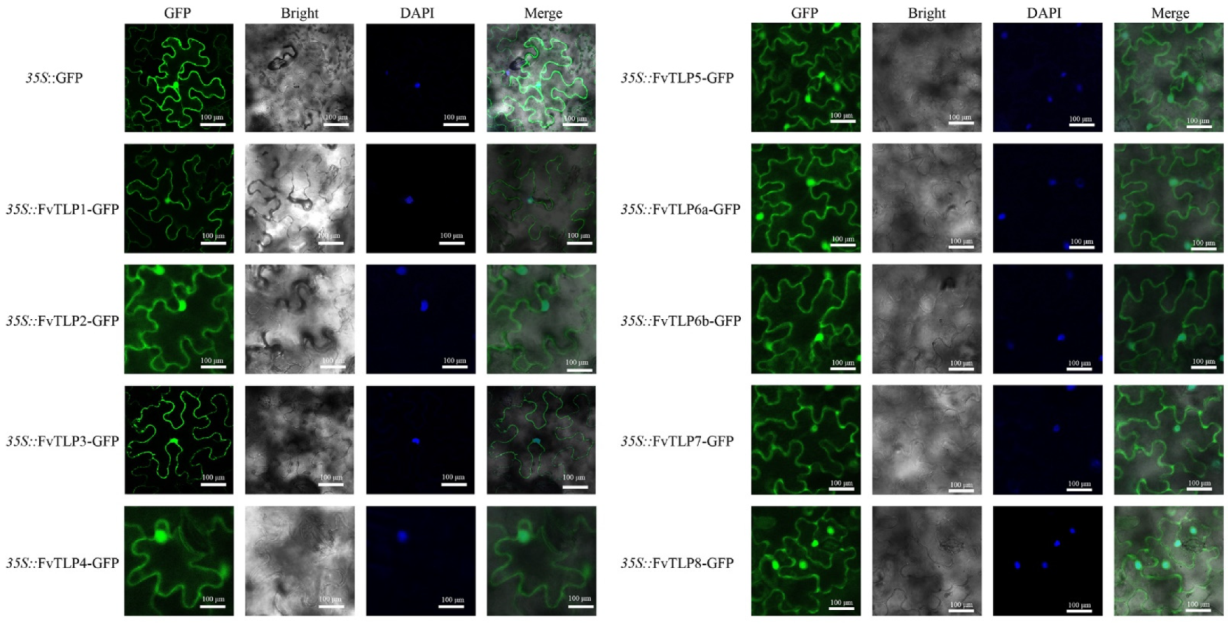
2.4. Subcellular Localization and Transcriptional Activity of FvTLPs
2.5. FvTLPs Interact with FvSKP1s
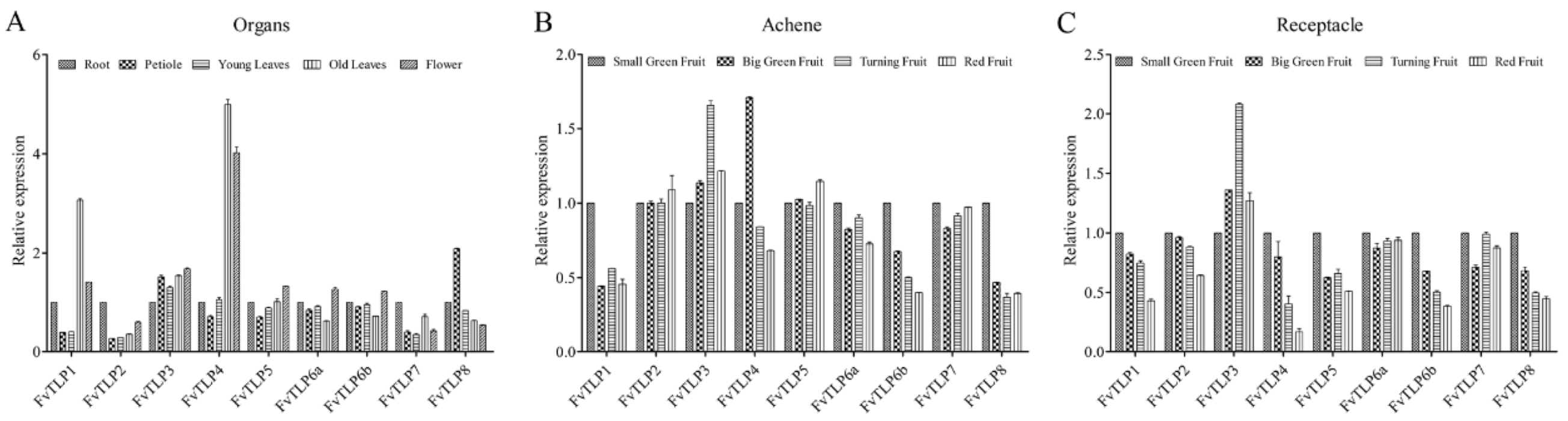
2.6. Expression Patterns of FvTLPs in Different Tissues of F. vesca
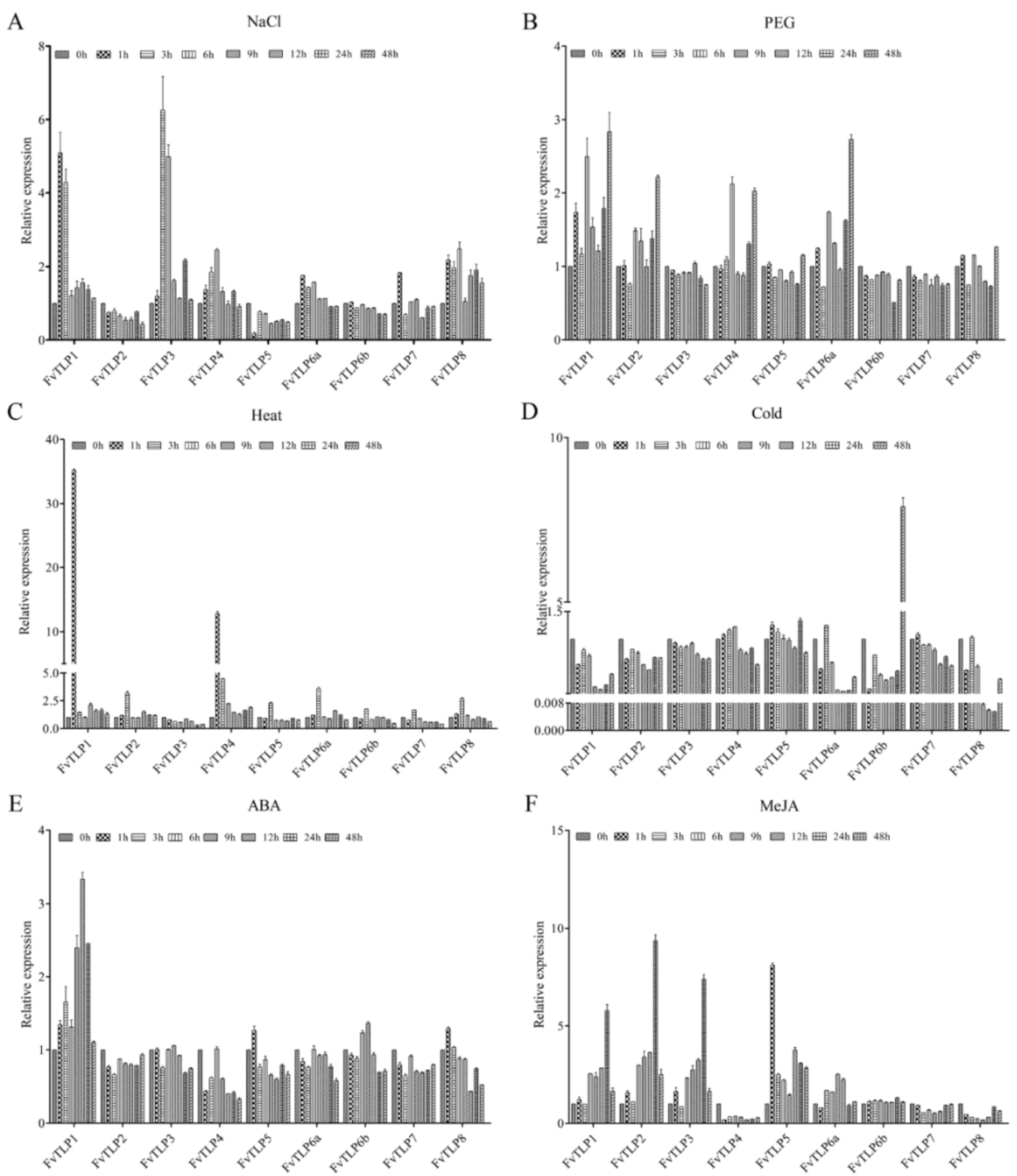
2.7. Responses of FvTLPs to Abiotic Stress and Exogenous Hormones in F. vesca Leaves
2.8. FvTLP1 Expression Responds to Abiotic Stress
3. Discussion
4. Materials and Methods
4.1. Genome-Scale Identification of FvTLPs
4.2. Gene Structure, Phylogenetic, and Cis-Element Analysis of FvTLPs
4.3. Plant Materials and Treatment
4.4. RNA Isolation and qRT-PCR Analysis
4.5. Subcellular Localization of FvTLPs
4.6. Transactivation Activity Assay for FvTLPs in Yeast
4.7. Yeast Two-Hybrid (Y2H) Assays
4.8. Plasmid Construction and Arabidopsis Transformation
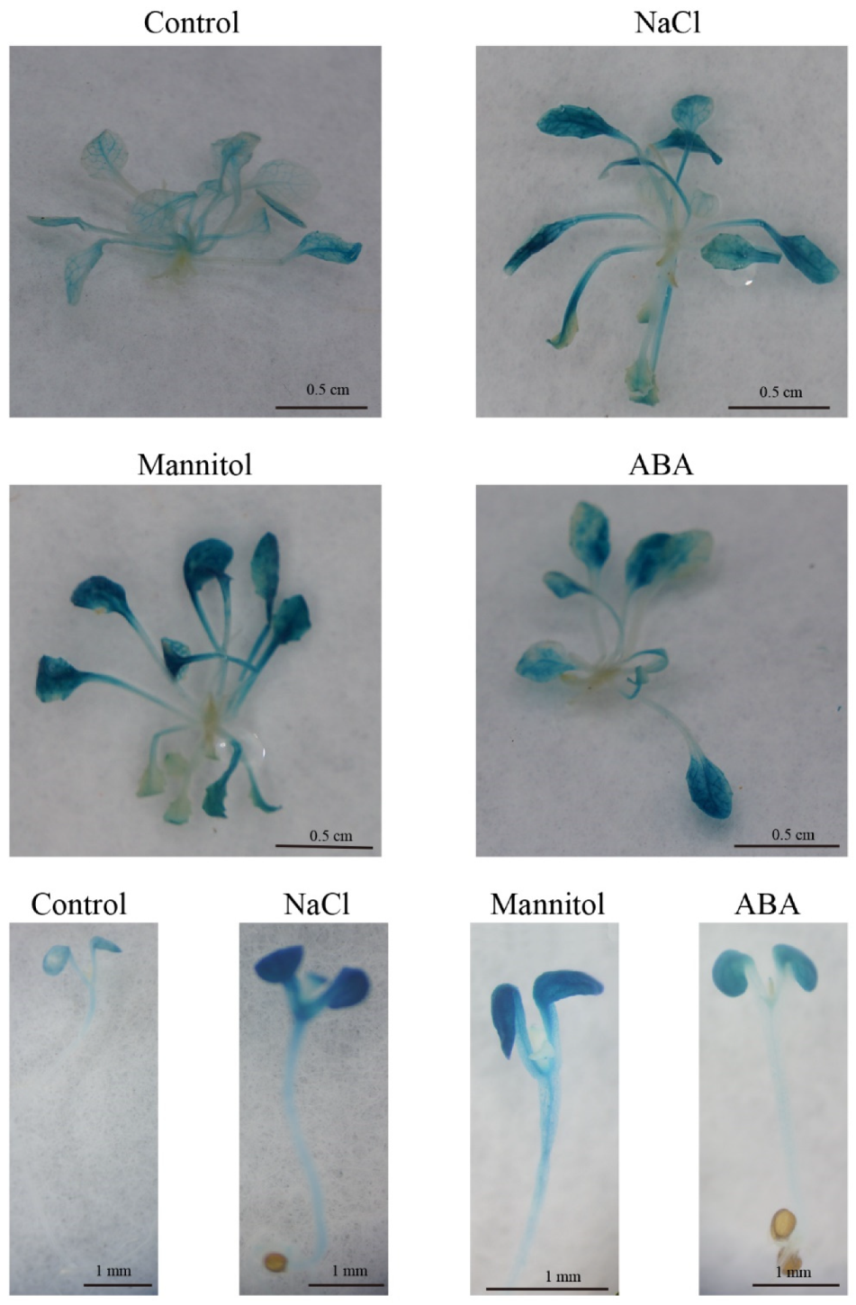
4.9. Histochemical Assay for GUS Activity
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Titus, J.B.; Wei-Song, S.; Sandro, S.; Samuel, C.M.; Lawrence, S. Implication of Tubby proteins as transcription factors by structure-based functional analysis. Science 1999, 286, 2119–2125. [Google Scholar]
- Bateman, A.; Finn, R.D.; Sims, P.J.; Wiedmer, T.; Biegert, A.; Soding, J. Phospholipid scramblases and Tubby-like proteins belong to a new superfamily of membrane tethered transcription factors. Bioinformatics 2009, 25, 159–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Min, J.H.; Dae, Y.K.; Yong, W.S. Interactions between wheat Tubby-like and SKP1-like proteins. Genes Genet. Syst. 2015, 90, 293–304. [Google Scholar]
- Kou, Y.; Qiu, D.; Wang, L.; Li, X.; Wang, S. Molecular analyses of the rice tubby-like protein gene family and their response to bacterial infection. Plant Cell Rep. 2009, 28, 113–121. [Google Scholar] [CrossRef]
- Albuquerque, E.V.S.; Petitot, A.; Da Silva, J.P.; Grossi-de-Sa, M.F.; Fernandez, D. Early responses of coffee immunity-related genes to root-knot nematode infection. Physiol. Mol. Plant Pathol. 2017, 100, 142–150. [Google Scholar] [CrossRef]
- Muthumeena, M.; Ashwin, R.; Ramesh, S.A.; Malathi, P.; Viswanathan, R. Expression profiling of transcription factors (TFs) in sugarcane X Colletotrichum falcatum interaction. J. Plant Biochem. Biotechnol. 2013, 22, 286–294. [Google Scholar]
- Zhang, Y.; Miao, L.; Yang, X.; Jiang, G. Genome-wide characterization and expression of the TLP gene family associated with Colletotrichum gloeosporioides inoculation in Fragaria × ananassa. PeerJ 2022, 10, e12979. [Google Scholar] [CrossRef]
- Cai, M.; Qiu, D.; Yuan, T.; Ding, X.; Li, H.; Duan, L.; Xu, C.; Li, X.; Wang, S. Identification of novel pathogen-responsive cis-elements and their binding proteins in the promoter of OsWRKY13, a gene regulating rice disease resistance. Plant Cell Environ. 2008, 31, 86–96. [Google Scholar] [CrossRef]
- Gharaghani, F.; Rafiei, F.; Mirakhorli, N.; Ebrahimie, E. Differential expression of TLP, ERF1, and R2R3MYB in annual Medicago species under salinity conditions. Genet. Mol. Res. 2015, 14, 10152–10164. [Google Scholar] [CrossRef]
- Janiak, A.; Kwasniewski, M.; Sowa, M.; Kuczyńska, A.; Mikołajczak, K.; Ogrodowicz, P.; Szarejko, I. Insights into barley root transcriptome under mild drought stress with an emphasis on gene expression regulatory mechanisms. Int. J. Mol. Sci. 2019, 20, 6139. [Google Scholar] [CrossRef] [Green Version]
- Yulong, C.; Wei, D.; Baoming, S.; Yang, Z.; Qing, M. Genome-wide identification and comparative analysis of the TUBBY-like protein gene family in maize. Genes Genom. 2016, 38, 25–36. [Google Scholar] [CrossRef]
- Dong, M.; Fan, X.; Pang, X.; Li, Y. Decrypting tubby-like protein gene family of multiple functions in starch root crop cassava. AoB Plants 2019, 11, plz075. [Google Scholar] [CrossRef]
- Xu, J.; Xing, S.; Zhang, Z.; Chen, X.; Wang, X. Genome-wide identification and expression analysis of the tubby-like protein family in the Malus domestica genome. Front. Plant Sci. 2016, 7, 1693. [Google Scholar] [CrossRef] [PubMed]
- Bano, N.; Fakhrah, S.; Mohanty, C.S.; Bag, S.K. Genome-wide identification and evolutionary analysis of Gossypium Tubby-Like Protein (TLP) gene family and expression analyses during salt and drought stress. Front. Plant Sci. 2021, 12, 667929. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Liu, Y.; Yu, T.; Hou, Z.; Zheng, J.; Chen, J.; Zhou, Y.; Chen, M.; Fu, J.; Ma, Y.; et al. Comprehensive profiling of Tubby-Like Proteins in soybean and roles of the GmTLP8 gene in abiotic stress responses. Front. Plant Sci. 2022, 13, 844545. [Google Scholar] [CrossRef]
- Altaf, A.; Zada, A.; Hussain, S.; Gull, S.; Ding, Y.; Tao, R.; Zhu, M.; Zhu, X. Genome-wide identification, characterization, and expression analysis of Tubby gene family in wheat (Triticum aestivum L.) under biotic and abiotic stresses. Agronomy 2022, 12, 1121. [Google Scholar] [CrossRef]
- Wang, K.; Cheng, Y.; Yi, L.; He, H.; Zhan, S.; Yang, P. Genome-wide identification of the Tubby-Like Protein (TLPs) family in medicinal model plant Salvia miltiorrhiza. PeerJ 2021, 9, e1140. [Google Scholar] [CrossRef]
- Bano, N.; Fakhrah, S.; Mohanty, C.S.; Bag, S.K. Transcriptome meta-analysis associated targeting hub genes and pathways of drought and salt stress responses in cotton (Gossypium hirsutum): A network biology approach. Front. Plant Sci. 2022, 13, 818472. [Google Scholar] [CrossRef]
- Xu, J.; Xing, S.; Sun, Q.; Zhan, C.; Liu, X.; Zhang, S.; Wang, X. The expression of a tubby-like protein from Malus domestica (MdTLP7) enhances abiotic stress tolerance in Arabidopsis. BMC Plant Biol. 2019, 19, 60–68. [Google Scholar] [CrossRef] [Green Version]
- Wardhan, V.; Pandey, A.; Chakraborty, S.; Chakraborty, N. Chickpea transcription factor CaTLP1 interacts with protein kinases, modulates ROS accumulation and promotes ABA-mediated stomatal closure. Sci. Rep. 2016, 6, 38121. [Google Scholar] [CrossRef] [Green Version]
- Vijay, W.; Kishwer, J.; Sonika, G.; Srinivasarao, C.; Asis, D.; Subhra, C.; Niranjan, C. Overexpression of CaTLP1, a putative transcription factor in chickpea (Cicer arietinum L.), promotes stress tolerance. Plant Mol. Biol. 2012, 79, 479–493. [Google Scholar]
- Li, S.; Zhang, J.; Liu, L.; Wang, Z.; Li, Y.; Guo, L.; Li, Y.; Zhang, X.; Ren, S.; Zhao, B.; et al. SlTLFP8 reduces water loss to improve water-use efficiency by modulating cell size and stomatal density via endoreduplication. Plant Cell Environ. 2020, 43, 2666–2679. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; He, X.; Su, D.; Feng, Y.; Zhao, H.; Deng, H.; Liu, M. Comprehensive profiling of Tubby-Like Protein expression uncovers ripening-related TLP genes in tomato (Solanum lycopersicum). Int. J. Mol. Sci. 2020, 21, 1000. [Google Scholar] [CrossRef] [Green Version]
- Bao, Y.; Song, W.; Jin, Y.; Jiang, C.; Yang, Y.; Li, B.; Huang, W.; Liu, H.; Zhang, H. Characterization of Arabidopsis Tubby-like proteins and redundant function of AtTLP3 and AtTLP9 in plant response to ABA and osmotic stress. Plant Mol. Biol. 2014, 86, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wang, Z.; Wang, F.; Lv, H.; Cao, M.; Zhang, N.; Li, F.; Wang, H.; Li, X.; Yuan, X.; et al. A tubby-like protein CsTLP8 acts in the ABA signaling pathway and negatively regulates osmotic stresses tolerance during seed germination. BMC Plant Biol. 2021, 21, 340. [Google Scholar] [CrossRef]
- Li, Z.; Wang, X.; Cao, X.; Chen, B.; Ma, C.; Lv, J.; Sun, Z.; Qiao, K.; Zhu, L.; Zhang, C.; et al. GhTULP34, a member of tubby-like proteins, interacts with GhSKP1A to negatively regulate plant osmotic stress. Genomics 2021, 113, 462–474. [Google Scholar] [CrossRef]
- Song, W.; Cheng, Z.; Guo, X.; Yu, C.; Wang, H.; Wang, J.; Li, B.; Zhang, H.; Bao, Y. Overexpression of NHL6 affects seed production in transgenic Arabidopsis plants. Plant Growth Regul. 2019, 88, 41–47. [Google Scholar] [CrossRef]
- Wang, M.; Xu, Z.; Ahmed, R.I.; Wang, Y.; Hu, R.; Zhou, G.; Kong, Y. Tubby-like Protein 2 regulates homogalacturonan biosynthesis in Arabidopsis seed coat mucilage. Plant Mol. Biol. 2019, 99, 421–436. [Google Scholar] [CrossRef]
- Myśków, B.; Góralska, M.; Lenarczyk, N.; Czyczyło-Mysza, I.; Stojałowski, S. Putative candidate genes responsible for leaf rolling in rye (Secale cereale L.). BMC Genet. 2018, 19, 57. [Google Scholar] [CrossRef]
- Santagata, S.; Boggon, T.J.; Baird, C.L.; Gomez, C.A.; Zhao, J.; Shan, W.S.; Myszka, D.G.; Shapiro, L. G-protein signaling through Tubby proteins. Science 2001, 292, 2041–2050. [Google Scholar] [CrossRef]
- Zea, D.J.; Monzon, A.M.; Parisi, G.; Marino-Buslje, C. How is structural divergence related to evolutionary information? Mol. Phylogenet. Evol. 2018, 127, 859–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, C.; Sun, Q.; Kong, Y. Genome-wide mining seed-specific candidate genes from peanut for promoter cloning. PLoS ONE 2019, 14, e214025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ain-Ali, Q.; Mushtaq, N.; Amir, R.; Gul, A.; Tahir, M.; Munir, F. Genome-wide promoter analysis, homology modeling and protein interaction network of Dehydration Responsive Element Binding (DREB) gene family in Solanum tuberosum. PLoS ONE 2021, 16, e261215. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhao, X.; Li, G.; Kumar, S.; Sun, Z.; Li, Y.; Guo, W.; Yang, J.; Hou, H. Genome-wide identification of the Nramp gene family in Spirodela polyrhiza and expression analysis under cadmium stress. Int. J. Mol. Sci. 2021, 22, 6414. [Google Scholar] [CrossRef]
- Yang, Z.; Zhou, Y.; Wang, X.; Gu, S.; Yu, J.; Liang, G.; Yan, C.; Xu, C. Genomewide comparative phylogenetic and molecular evolutionary analysis of tubby-like protein family in Arabidopsis, rice, and poplar. Genomics 2008, 92, 246–253. [Google Scholar] [CrossRef] [Green Version]
- Márquez, Y.; Mantica, F.; Cozzuto, L.; Burguera, D.; Hermoso-Pulido, A.; Ponomarenko, J.; Roy, S.W.; Irimia, M. ExOrthist: A tool to infer exon orthologies at any evolutionary distance. Genome Biol. 2021, 22, 239. [Google Scholar] [CrossRef]
- Koralewski, T.E.; Krutovsky, K.V. Evolution of exon-intron structure and alternative splicing. PLoS ONE 2011, 6, e18055. [Google Scholar] [CrossRef] [Green Version]
- Heidari, P.; Puresmaeli, F.; Mora-Poblete, F. Genome-wide identification and molecular evolution of the Magnesium Transporter (MGT) gene family in Citrullus lanatus and Cucumis sativus. Agronomy 2022, 12, 2253. [Google Scholar] [CrossRef]
- Al-Saharin, R.; Hellmann, H.; Mooney, S. Plant E3 ligases and their role in abiotic stress response. Cells 2022, 11, 890. [Google Scholar] [CrossRef]
- Wei, Y.Q.; Yuan, J.J.; Xiao, C.C.; Li, G.X.; Yan, J.Y.; Zheng, S.J.; Ding, Z.J. RING-box proteins regulate leaf senescence and stomatal closure via repression of ABA transporter gene ABCG40. J. Integr. Plant Biol. 2022, 64, 979–994. [Google Scholar] [CrossRef]
- Wang, Z.; Gao, M.; Li, Y.; Zhang, J.; Su, H.; Cao, M.; Liu, Z.; Zhang, X.; Zhao, B.; Guo, Y.; et al. The transcription factor SlWRKY37 positively regulates jasmonic acid- and dark-induced leaf senescence in tomato. J. Exp. Bot. 2022, erac258. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Dai, Z.; Zeng, B.; Li, J.; Ouyang, J.; Kang, L.; Wang, W.; Jia, W. Autocatalytic biosynthesis of abscisic acid and its synergistic action with auxin to regulate strawberry fruit ripening. Hortic. Res. 2022, 9, uhab076. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Bigotes, A.; Figueroa, P.M.; Figueroa, C.R. Jasmonate metabolism and its relationship with abscisic acid during strawberry fruit development and ripening. J. Plant Growth Regul. 2018, 37, 101–113. [Google Scholar] [CrossRef]
- Laloum, T.; Martín, G.; Duque, P. Alternative splicing control of abiotic stress responses. Trends Plant Sci. 2018, 23, 140–150. [Google Scholar] [CrossRef]
- Li, S.; Yu, X.; Cheng, Z.; Zeng, C.; Li, W.; Zhang, L.; Peng, M. Large-scale analysis of the cassava transcriptome reveals the impact of cold stress on alternative splicing. J. Exp. Bot. 2019, 71, 422–434. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Du, Q.; Campbell, M.; Yu, B.; Walia, H.; Zhang, C. Genome-wide discovery of natural variation in pre-mRNA splicing and prioritising causal alternative splicing to salt stress response in rice. New Phytol. 2021, 230, 1273–1287. [Google Scholar] [CrossRef] [PubMed]
- Thatcher, S.R.; Danilevskaya, O.N.; Meng, X.; Beatty, M.; Zastrow-Hayes, G.; Harris, C.; Van Allen, B.; Habben, J.; Li, B. Genome-wide analysis of alternative splicing during development and drought stress in maize. Plant Physiol. 2015, 170, 586–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, Y.; Serrano, N.; Gao, G.; Atia, M.; Mokhtar, M.; Woo, Y.H.; Bazin, J.; Veluchamy, A.; Benhamed, M.; Crespi, M.; et al. Thermopriming triggers splicing memory in Arabidopsis. J. Exp. Bot. 2018, 69, 2659–2675. [Google Scholar] [CrossRef] [Green Version]
- Okamoto, M.; Matsui, A.; Tanaka, M.; Morosawa, T.; Ishida, J.; Iida, K.; Mochizuki, Y.; Toyoda, T.; Seki, M. Sm-Like Protein-mediated RNA metabolism is required for heat stress tolerance in Arabidopsis. Front. Plant Sci. 2016, 7, 1079. [Google Scholar] [CrossRef] [Green Version]
- Chong, G.L.; Foo, M.H.; Lin, W.; Wong, M.M.; Verslues, P.E. Highly ABA-Induced 1 (HAI1)-Interacting protein HIN1 and drought acclimation-enhanced splicing efficiency at intron retention sites. Proc. Natl. Acad. Sci. USA 2019, 116, 22376–22385. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Li, X.; Guo, Q.; Liu, P.; Li, Y.; Wu, C.; Yang, G.; Huang, J.; Zhang, S.; Zheng, C.; et al. Salt responsive alternative splicing of a RING finger E3 ligase modulates the salt stress tolerance by fine-tuning the balance of COP9 signalosome subunit 5A. PLoS Genet. 2021, 17, e1009898. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Zheng, C.; Ji, G.; Jiang, P. Synergistic effects of HSE and LTR elements from HSP70 gene promoter of Ulva prolifera (Ulvophyceae, Chlorophyta) upon temperature induction1. J. Phycol. 2019, 55, 738–743. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Wang, N.; Wang, Y.; Jiang, S.; Fang, H.; Zhang, J.; Su, M.; Zuo, W.; Xu, L.; Zhang, Z.; et al. Overexpression of the transcription factor MdbHLH33 increases cold tolerance of transgenic apple callus. Plant Cell Tissue Organ Cult. 2018, 134, 131–140. [Google Scholar] [CrossRef]
- Wang, N.; Qu, C.; Jiang, S.; Chen, Z.; Xu, H.; Fang, H.; Su, M.; Zhang, J.; Wang, Y.; Liu, W.; et al. The proanthocyanidin-specific transcription factor MdMYBPA1 initiates anthocyanin synthesis under low-temperature conditions in red-fleshed apples. Plant J. 2018, 96, 39–55. [Google Scholar] [CrossRef]
- Wang, D.; Sun, Z.; Hu, X.; Xiong, J.; Hu, L.; Xu, Y.; Tang, Y.; Wu, Y. The key regulator LcERF056 enhances salt tolerance by modulating reactive oxygen species-related genes in Lotus corniculatus. BMC Plant Biol. 2021, 21, 605. [Google Scholar] [CrossRef]
- Mao, L.; Deng, M.; Jiang, S.; Zhu, H.; Yang, Z.; Yue, Y.; Zhao, K. Characterization of the DREBA4-type transcription factor (SlDREBA4), which contributes to heat tolerance in tomatoes. Front. Plant Sci. 2020, 11, 554520. [Google Scholar] [CrossRef]
- Xu, Z.; Wang, M.; Guo, Z.; Zhu, X.; Xia, Z. Identification of a 119-bp promoter of the maize sulfite oxidase gene (ZmSO) That confers high-level gene expression and ABA or drought inducibility in transgenic plants. Int. J. Mol. Sci. 2019, 20, 3326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Zhang, J.; Li, J.; Li, H.; Zhang, G. The functional and regulatory mechanisms of the Thellungiella salsuginea Ascorbate Peroxidase 6 (TsAPX6) in response to salinity and water deficit stresses. PLoS ONE 2016, 11, e154042. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Zhou, L.; Gao, H.; Wang, X.; Li, Z.; Li, Y. The transcription factor MdMYB2 influences cold tolerance and anthocyanin accumulation by activating SUMO E3 ligase MdSIZ1 in apple. Plant Physiol. 2022, 189, 2044–2060. [Google Scholar] [CrossRef]
- Freitas, F.Z.; Virgilio, S.; Cupertino, F.B.; Kowbel, D.J.; Fioramonte, M.; Gozzo, F.C.; Glass, N.L.; Bertolini, M.C. The SEB-1 transcription factor binds to the STRE motif in Neurospora crassa and Regulates a Variety of Cellular Processes Including the Stress Response and Reserve Carbohydrate Metabolism. G3-Genes Genomes Genet. 2016, 6, 1327–1343. [Google Scholar] [CrossRef] [Green Version]
- Kim, H. Disruption of YCP4 enhances freeze-thaw tolerance in Saccharomyces cerevisiae. Biotechnol. Lett. 2022, 44, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Sun, R.; Liu, H.; Chang, L.; Li, S.; Zhao, M.; Shennan, C.; Lei, J.; Dong, J.; Zhong, C.; et al. Complete chloroplast genome sequencing of ten wild Fragaria species in China provides evidence for phylogenetic evolution of Fragaria. Genomics 2021, 113, 1170–1179. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Huang, X.; Lin, Y.; Wang, X.; Yan, Z.; Wang, Q.; Ding, J.; Gu, T.; Li, Y. Identification of reference genes for transcript normalization in various tissue types and seedlings subjected to different abiotic stresses of woodland strawberry Fragaria vesca. Sci. Hortic. 2020, 261, 108840. [Google Scholar] [CrossRef]
- Liang, Y.; Jiang, C.; Liu, Y.; Gao, Y.; Lu, J.; Aiwaili, P.; Fei, Z.; Jiang, C.; Hong, B.; Ma, C.; et al. Auxin regulates sucrose transport to repress petal abscission in rose (Rosa hybrida). Plant Cell 2020, 32, 3485–3499. [Google Scholar] [CrossRef]
- Gong, Z.; Luo, Y.; Zhang, W.; Jian, W.; Zhang, L.; Gao, X.; Hu, X.; Yuan, Y.; Wu, M.; Xu, X.; et al. A SlMYB75-centred transcriptional cascade regulates trichome formation and sesquiterpene accumulation in tomato. J. Exp. Bot. 2021, 72, 3806–3820. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, L.; Chen, X.; Zhao, L.; Liu, X.; Wang, Y.; Wu, G.; Xia, C.; Zhang, L.; Kong, X. The protein phosphatase 2C clade A TaPP2CA interact with calcium-dependent protein kinases, TaCDPK5/TaCDPK9–1, that phosphorylate TabZIP60 transcription factor from wheat (Triticum aestivum L.). Plant Sci. 2022, 321, 111304. [Google Scholar] [CrossRef]
- Jameel, A.; Ketehouli, T.; Wang, Y.; Wang, F.; Li, X.; Li, H. Detection and validation of cis-regulatory motifs in osmotic stress-inducible synthetic gene switches via computational and experimental approaches. Funct. Plant Biol. 2022. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | CDS (bp) | Peptide (aa) | Predicted Isoelectric | Molecular Weight (kD) | Amino Acid Residue Localization of F-Box Domain | Amino Acid Residue Localization of Tubby Domain | |
|---|---|---|---|---|---|---|---|---|
| FvTLP1 | FvH4_1g13070 | 1254 | 417 | 9.31 | 46.9 | 54–99 | 117–412 | |
| FvTLP2 | FvH4_2g13720 | 1290 | 429 | 9.74 | 48.1 | 53–95 | 116–424 | |
| FvTLP3 | FvH4_2g24790 | 1215 | 404 | 9.46 | 44.9 | 49–94 | 114–399 | |
| FvTLP4 | FvH4_2g39640 | 1308 | 435 | 9.11 | 48.9 | 58–100 | 125–340 | |
| FvTLP5 | FvH4_3g18250 | 1077 | 358 | 9.43 | 40.1 | 41–86 | 106–353 | |
| FvTLP6a | FvH4_4g17170.t1 | 1188 | 395 | 9.78 | 44.2 | 37–90 | 102–390 | |
| FvTLP6b | FvH4_4g17170.t2 | 1173 | 390 | 9.69 | 43.6 | 37–90 | 102–385 | |
| FvTLP7 | FvH4_5g30230 | 1284 | 427 | 9.47 | 48.0 | 53–95 | 116–422 | |
| FvTLP8 | FvH4_6g28190 | 1245 | 414 | 9.39 | 46.2 | 166–408 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Wang, G.; Chang, L.; Sun, R.; Wu, R.; Zhong, C.; Gao, Y.; Zhang, H.; Wei, L.; Wei, Y.; et al. Genome-Wide Identification, Characterization, and Expression Analysis of Tubby-like Protein (TLP) Gene Family Members in Woodland Strawberry (Fragaria vesca). Int. J. Mol. Sci. 2022, 23, 11961. https://doi.org/10.3390/ijms231911961
Li S, Wang G, Chang L, Sun R, Wu R, Zhong C, Gao Y, Zhang H, Wei L, Wei Y, et al. Genome-Wide Identification, Characterization, and Expression Analysis of Tubby-like Protein (TLP) Gene Family Members in Woodland Strawberry (Fragaria vesca). International Journal of Molecular Sciences. 2022; 23(19):11961. https://doi.org/10.3390/ijms231911961
Chicago/Turabian StyleLi, Shuangtao, Guixia Wang, Linlin Chang, Rui Sun, Ruishuang Wu, Chuanfei Zhong, Yongshun Gao, Hongli Zhang, Lingzhi Wei, Yongqing Wei, and et al. 2022. "Genome-Wide Identification, Characterization, and Expression Analysis of Tubby-like Protein (TLP) Gene Family Members in Woodland Strawberry (Fragaria vesca)" International Journal of Molecular Sciences 23, no. 19: 11961. https://doi.org/10.3390/ijms231911961
APA StyleLi, S., Wang, G., Chang, L., Sun, R., Wu, R., Zhong, C., Gao, Y., Zhang, H., Wei, L., Wei, Y., Zhang, Y., Dong, J., & Sun, J. (2022). Genome-Wide Identification, Characterization, and Expression Analysis of Tubby-like Protein (TLP) Gene Family Members in Woodland Strawberry (Fragaria vesca). International Journal of Molecular Sciences, 23(19), 11961. https://doi.org/10.3390/ijms231911961