
The miR156-Targeted SQUAMOSA PROMOTER BINDING PROTEIN (PmSBP) Transcription Factor Regulates the Flowering Time by Binding to the Promoter of SUPPRESSOR OF OVEREXPRESSION OF CO1 (PmSOC1) in Prunus mume

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Chromosomal Location, Phylogenetic Analysis and Sequence Analysis of miR156 from P. mume
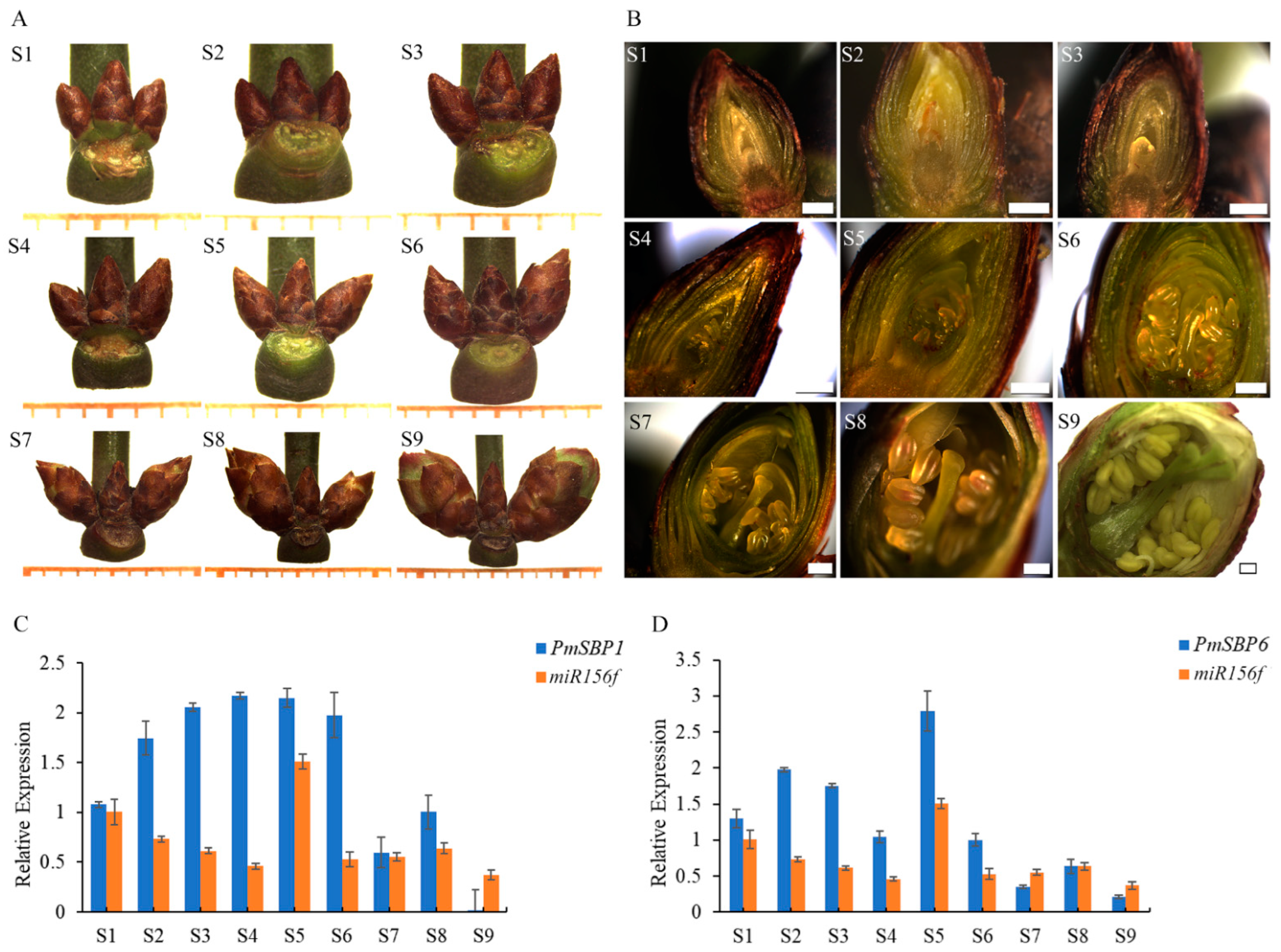
2.2. Expression Patterns of PmSBP1/6 and miR156f during the Flower Development Period
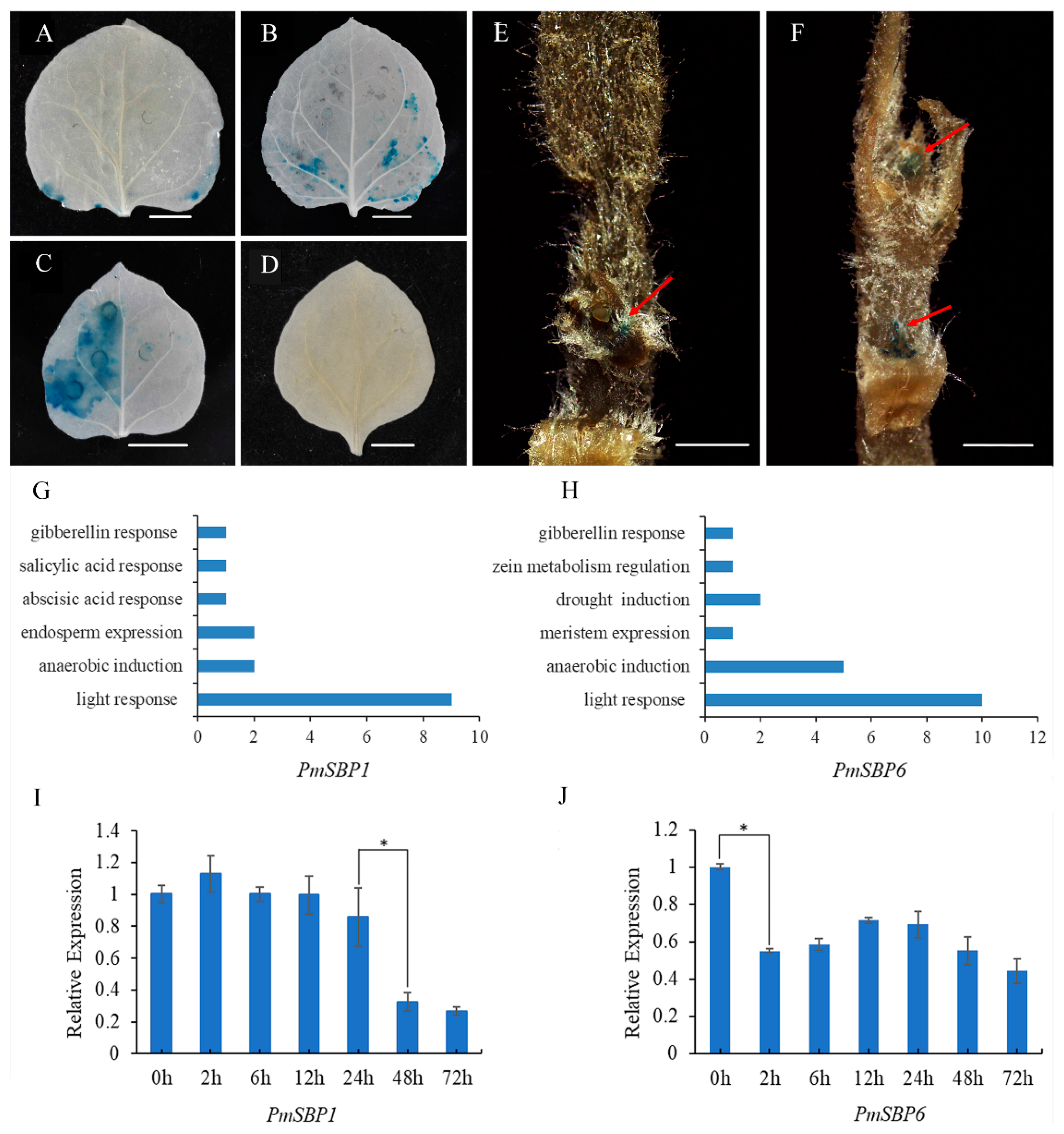
2.3. The Activity Analysis of PmSBP1/6 Promoters
2.4. The Promoter Cis-Elements Composition of PmSBP1/6 and Their Response to GA3
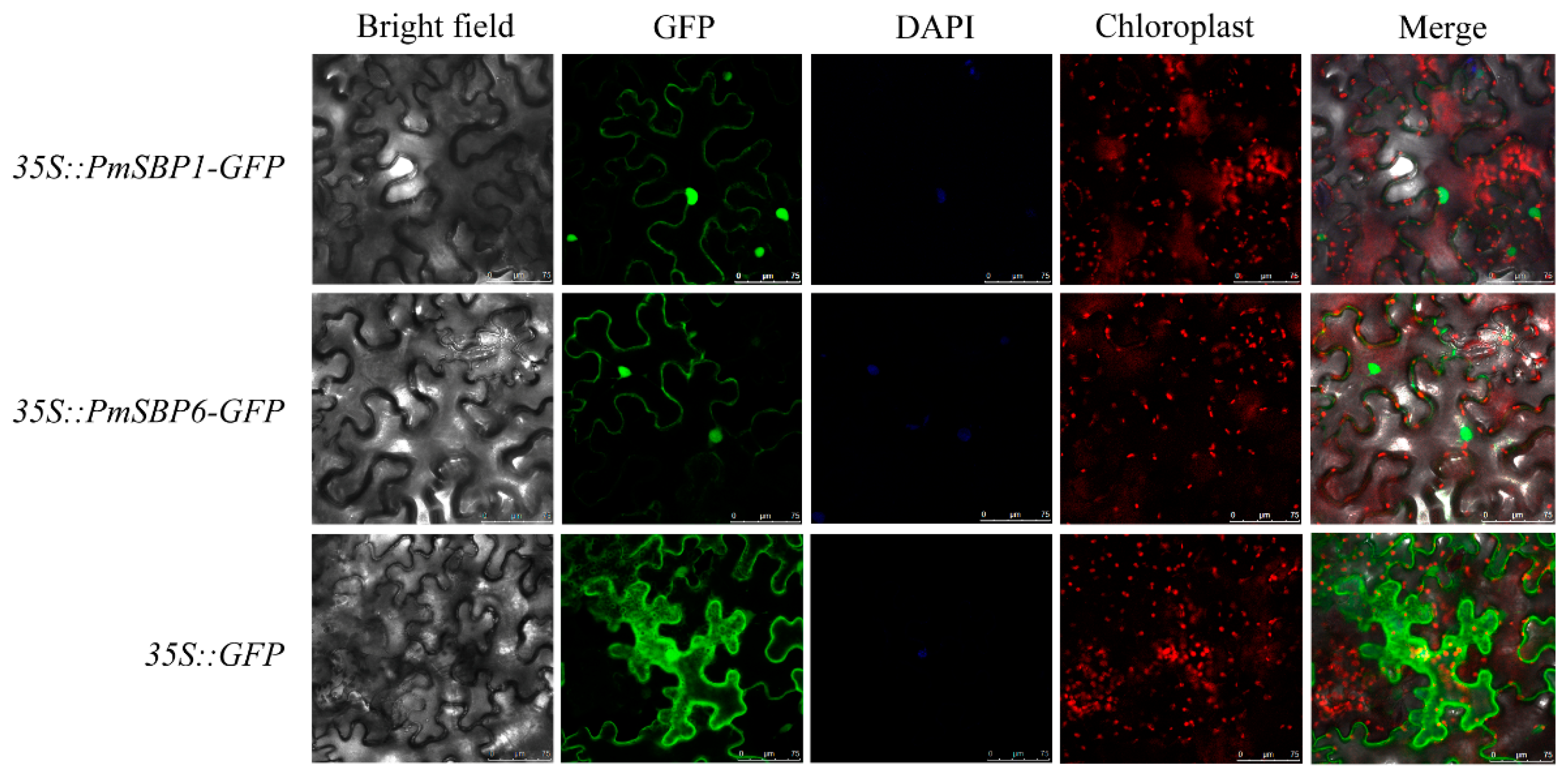
2.5. Subcellular Localization of PmSBP1/6 Proteins
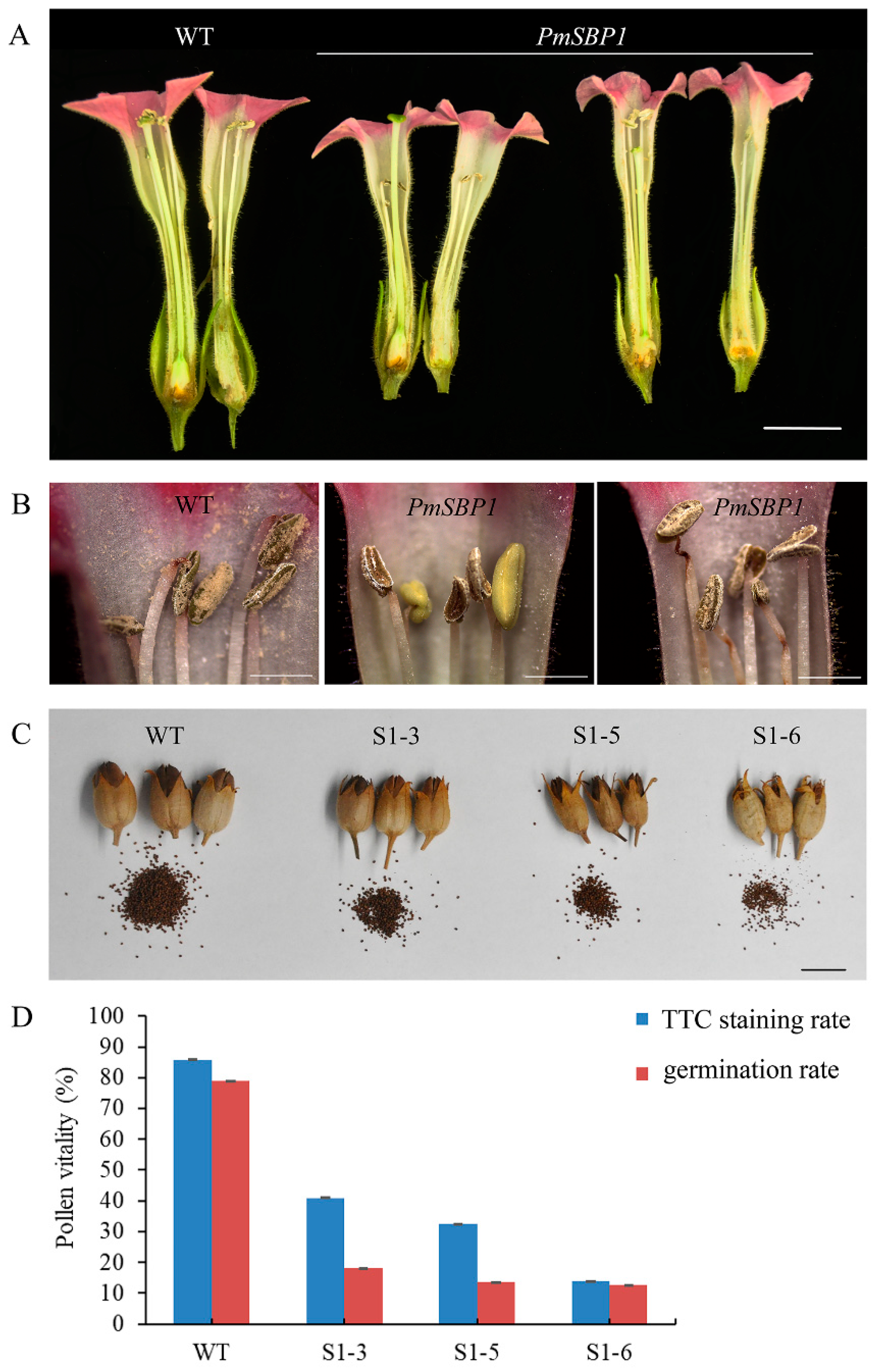
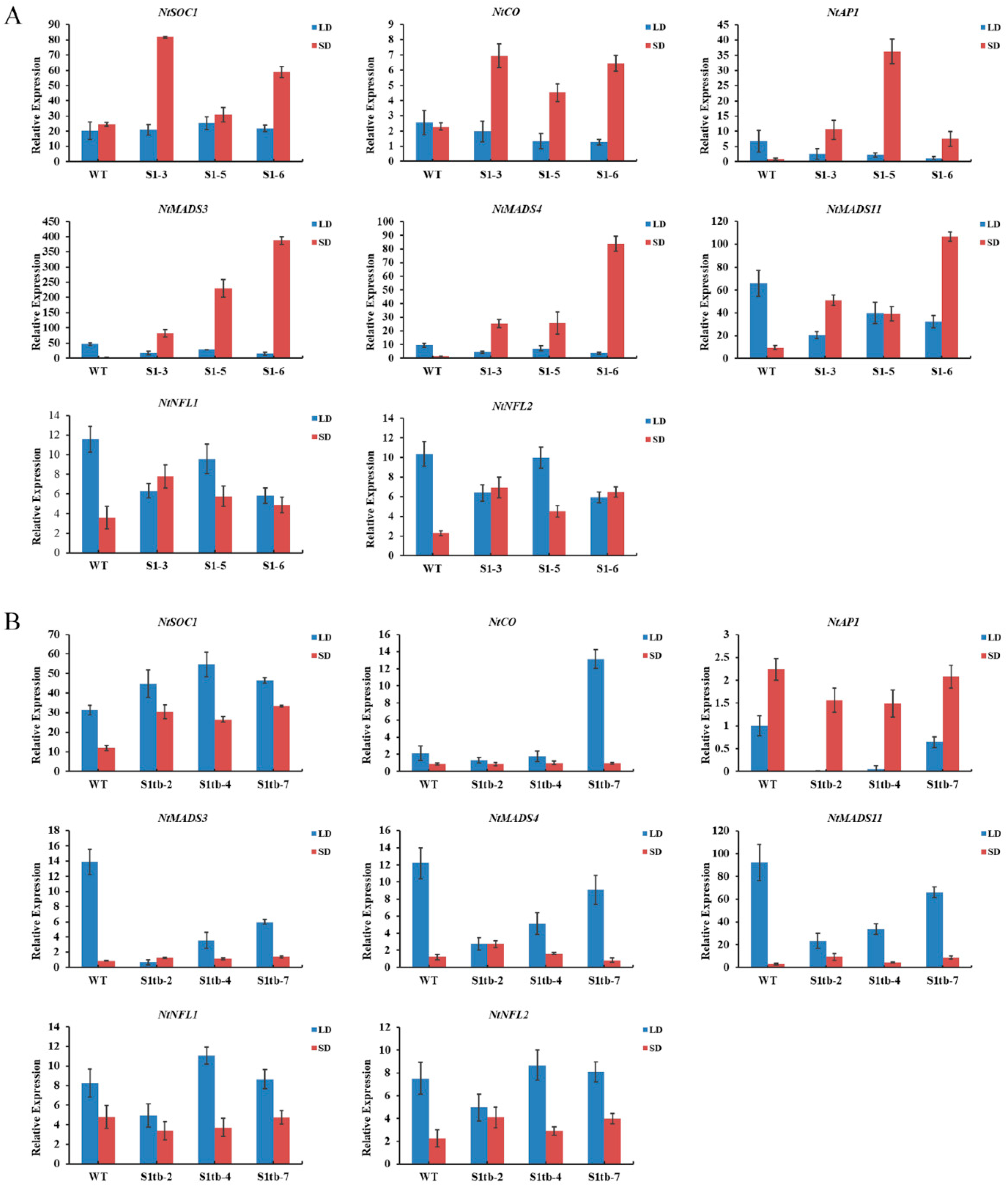
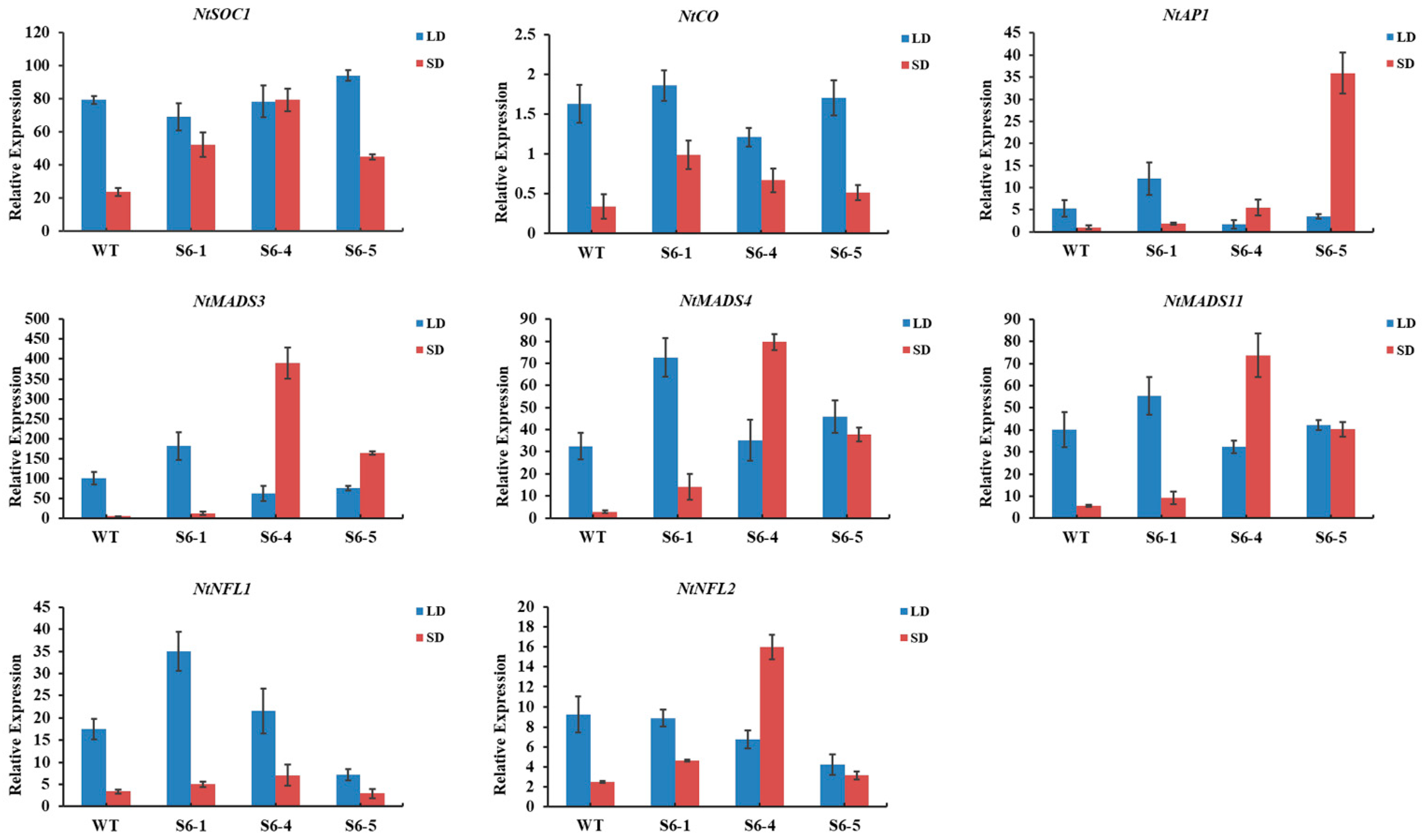
2.6. Overexpression of PmSBP1/1tb and PmSBP6 in Tobacco Promotes Flowering and Affects Fertility
2.7. Expression Pattern of Endogenous Flowering-Related Genes in PmSBP1/1tb and PmSBP6 Transgenic Tobacco
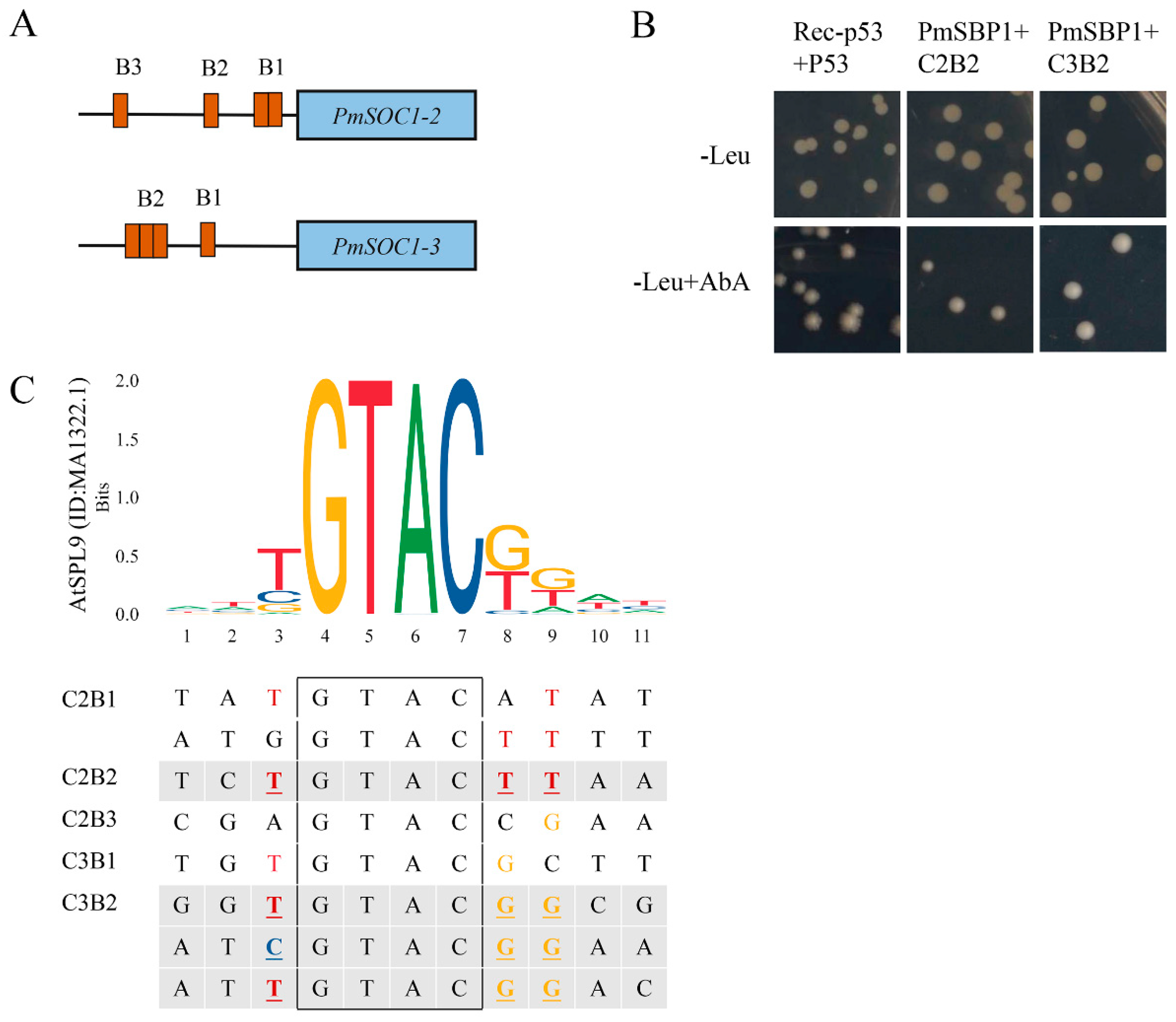
2.8. PmSBP1 Binding to the PmSOC1s Promoters
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Bioinformatics Analysis of pmu-miR156
4.3. Cloning and Analysis of PmSBP Gene and Promoter
4.4. Expression Pattern Analysis
4.5. Subcellular Localization
4.6. Promoter Activity Analysis
4.7. Overexpression of PmSBP1/6 and Their Promoters in Tobacco
4.8. Phenotypic Statistical Analysis
4.9. Yeast One-Hybrid Assay
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Moon, J.; Lee, H.; Kim, M.; Lee, I. Analysis of Flowering Pathway Integrators in Arabidopsis. Plant Cell Physiol. 2005, 46, 292–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.-W.; Czech, B.; Weigel, D. miR156-Regulated SPL Transcription Factors Define an Endogenous Flowering Pathway in Arabidopsis thaliana. Cell 2009, 138, 738–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.; Park, M.Y.; Conway, S.R.; Wang, J.-W.; Weigel, D.; Poethig, R.S. The Sequential Action of miR156 and miR172 Regulates Developmental Timing in Arabidopsis. Cell 2009, 138, 750–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.; Poethig, R.S. Temporal regulation of shoot development in Arabidopsis thaliana by miR156 and its target SPL3. Development 2006, 133, 3539–3547. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.-W.; Park, M.Y.; Wang, L.-J.; Koo, Y.; Chen, X.-Y.; Weigel, D.; Poethig, R.S. MiRNA Control of Vegetative Phase Change in Trees. PLoS Genet. 2011, 7, e1002012. [Google Scholar] [CrossRef] [Green Version]
- Xie, K.; Shen, J.; Hou, X.; Yao, J.; Li, X.; Xiao, J.; Xiong, L. Gradual Increase of miR156 Regulates Temporal Expression Changes of Numerous Genes during Leaf Development in Rice. Plant Physiol. 2012, 158, 1382–1394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergonzi, S.; Albani, M.C.; van Themaat, E.V.L.; Nordström, K.J.V.; Wang, R.; Schneeberger, K.; Moerland, P.D.; Coupland, G. Mechanisms of Age-Dependent Response to Winter Temperature in Perennial Flowering of Arabis alpina. Science 2013, 340, 1094–1097. [Google Scholar] [CrossRef]
- Zhou, C.-M.; Zhang, T.-Q.; Wang, X.; Yu, S.; Lian, H.; Tang, H.; Feng, Z.-Y.; Zozomova-Lihová, J.; Wang, J.-W. Molecular Basis of Age-Dependent Vernalization in Cardamine flexuosa. Science 2013, 340, 1097–1100. [Google Scholar] [CrossRef]
- Wei, S.; Gruber, M.Y.; Yu, B.; Gao, M.-J.; Khachatourians, G.G.; Hegedus, D.D.; AP Parkin, I.; Hannoufa, A. Arabidopsis mutant sk156 reveals complex regulation of SPL15 in a miR156-controlled gene network. BMC Plant Biol. 2012, 12, 169. [Google Scholar] [CrossRef] [Green Version]
- Shikata, M.; Koyama, T.; Mitsuda, N.; Ohme-Takagi, M. Arabidopsis SBP-Box Genes SPL10, SPL11 and SPL2 Control Morphological Change in Association with Shoot Maturation in the Reproductive Phase. Plant Cell Physiol. 2009, 50, 2133–2145. [Google Scholar] [CrossRef]
- Xu, M.; Hu, T.; Zhao, J.; Park, M.-Y.; Earley, K.W.; Wu, G.; Yang, L.; Poethig, R.S. Developmental Functions of miR156-Regulated SQUAMOSA PROMOTER BINDING PROTEIN-LIKE (SPL) Genes in Arabidopsis thaliana. PLoS Genet. 2016, 12, e1006263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyun, Y.; Richter, R.; Vincent, C.; Martinez-Gallegos, R.; Porri, A.; Coupland, G. Multi-layered Regulation of SPL15 and Cooperation with SOC1 Integrate Endogenous Flowering Pathways at the Arabidopsis Shoot Meristem. Dev. Cell 2016, 37, 254–266. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.-H.; Ju, Y.; Seo, P.J.; Lee, J.-H.; Park, C.-M. The SOC1-SPL module integrates photoperiod and gibberellic acid signals to control flowering time in Arabidopsis. Plant J. 2012, 69, 577–588. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.H.; Seo, Y.H.; Seo, P.J.; Reyes, J.L.; Yun, J.; Chua, N.H.; Park, C.M. The GIGANTEA-regulated microRNA172 mediates photoperiodic flowering independent of CONSTANS in Arabidopsis. Plant Cell 2007, 19, 2736–2748. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.-H.; Lee, H.-J.; Ryu, J.Y.; Park, C.-M. SPL3/4/5 Integrate Developmental Aging and Photoperiodic Signals into the FT-FD Module in Arabidopsis Flowering. Mol. Plant 2016, 9, 1647–1659. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, A.; Wu, M.-F.; Yang, L.; Wu, G.; Poethig, R.S.; Wagner, D. The MicroRNA-Regulated SBP-Box Transcription Factor SPL3 Is a Direct Upstream Activator of LEAFY, FRUITFULL, and APETALA1. Dev. Cell 2009, 17, 268–278. [Google Scholar] [CrossRef] [Green Version]
- Unte, U.S.; Sorensen, A.-M.; Pesaresi, P.; Gandikota, M.; Leister, D.; Saedler, H.; Huijser, P. SPL8, an SBP-Box Gene That Affects Pollen Sac Development in Arabidopsis. Plant Cell 2003, 15, 1009–1019. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Xu, Z.; Yang, W.; Cheng, T.; Wang, J.; Zhang, Q. Isolation and Functional Characterization of SOC1-like Genes in Prunus mume. J. Am. Soc. Hortic. Sci. 2016, 141, 315–326. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhou, Y.; Yang, W.; Cheng, T.; Wang, J.; Zhang, Q. Isolation and functional characterization of SVP-like genes in Prunus mume. Sci. Hortic. 2017, 215, 91–101. [Google Scholar] [CrossRef]
- Ahmad, S.; Li, Y.; Yang, Y.; Zhou, Y.; Zhao, K.; Zhang, Q. Isolation, functional characterization and evolutionary study of LFY1 gene in Prunus mume. Plant Cell Tissue Organ Cult. (PCTOC) 2018, 136, 523–536. [Google Scholar] [CrossRef]
- Xu, Z.; Sun, L.; Zhou, Y.; Yang, W.; Cheng, T.; Wang, J.; Zhang, Q. Identification and expression analysis of the SQUAMOSA promoter-binding protein (SBP)-box gene family in Prunus mume. Mol. Genet. Genom. 2015, 290, 1701–1715. [Google Scholar] [CrossRef] [PubMed]
- Gramzow, L.; Theißen, G. Plant miRNA Conservation and Evolution. Methods Mol. Biol. 2019, 1932, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Ye, M.; Sang, M.; Wu, R. A Regulatory Network for miR156-SPL Module in Arabidopsis thaliana. Int. J. Mol. Sci. 2019, 20, 6166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Y.; Guo, Z.; Li, L. Evolutionary conservation of microRNA regulatory programs in plant flower development. Dev. Biol. 2013, 380, 133–144. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Pan, H.; Wang, J.; Yang, W.; Cheng, T.; Zhang, Q. Identification and profiling of novel and conserved microRNAs during the flower opening process in Prunus mume via deep sequencing. Mol. Genet. Genom. 2014, 289, 169–183. [Google Scholar] [CrossRef]
- Xing, S.; Salinas, M.; Höhmann, S.; Berndtgen, R.; Huijser, P. miR156-Targeted and Nontargeted SBP-Box Transcription Factors Act in Concert to Secure Male Fertility in Arabidopsis. Plant Cell 2010, 22, 3935–3950. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Peng, J.; Wang, M.; Su, W.; Gan, X.; Jing, Y.; Yang, X.; Lin, S.; Gao, Y. The Role of EjSPL3, EjSPL4, EjSPL5, and EjSPL9 in Regulating Flowering in Loquat (Eriobotrya japonica Lindl.). Int. J. Mol. Sci. 2020, 21, 248. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Li, J.; Liu, Y.; Zhang, Q.; Gao, Y.; Fang, K.; Cao, Q.; Qin, L.; Xing, Y. Roles of the GA-mediated SPL Gene Family and miR156 in the Floral Development of Chinese Chestnut (Castanea mollissima). Int. J. Mol. Sci. 2019, 20, 1577. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Quan, S.; Ma, L.; Xu, H.; Yang, J.; Niu, J. Molecular characterization of SBP-box gene family during floral induction in walnut (Juglans regia L.). Tree Genet. Genomes 2020, 16, 12. [Google Scholar] [CrossRef]
- Li, X.-Y.; Lin, E.-P.; Huang, H.-H.; Niu, M.-Y.; Tong, Z.-K.; Zhang, J.-H. Molecular Characterization of SQUAMOSA PROMOTER BINDING PROTEIN-LIKE (SPL) Gene Family in Betula luminifera. Front. Plant Sci. 2018, 9, 608. [Google Scholar] [CrossRef]
- Xing, S.; Quodt, V.; Chandler, J.; Höhmann, S.; Berndtgen, R.; Huijser, P. SPL8 Acts Together with the Brassinosteroid-Signaling Component BIM1 in Controlling Arabidopsis thaliana Male Fertility. Plants 2013, 2, 416–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Schwarz, S.; Saedler, H.; Huijser, P. SPL8, a local regulator in a subset of gibberellin-mediated developmental processes in Arabidopsis. Plant Mol. Biol. 2007, 63, 429–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, S.; Salinas, M.; Garcia-Molina, A.; Höhmann, S.; Berndtgen, R.; Huijser, P. SPL8and miR156-targetedSPLgenes redundantly regulate Arabidopsis gynoecium differential patterning. Plant J. 2013, 75, 566–577. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, S.; Grande, A.V.; Bujdoso, N.; Saedler, H.; Huijser, P. The microRNA regulated SBP-box genes SPL9 and SPL15 control shoot maturation in Arabidopsis. Plant Mol. Biol. 2008, 67, 183–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, J.-H.; Seo, P.J.; Kang, S.K.; Park, C.-M. miR172 signals are incorporated into the miR156 signaling pathway at the SPL3/4/5 genes in Arabidopsis developmental transitions. Plant Mol. Biol. 2011, 76, 35–45. [Google Scholar] [CrossRef]
- Feng, S.; Xu, Y.; Guo, C.; Zheng, J.; Zhou, B.; Zhang, Y.; Ding, Y.; Zhang, L.; Zhu, Z.; Wang, H.; et al. Modulation of miR156 to identify traits associated with vegetative phase change in tobacco (Nicotiana tabacum). J. Exp. Bot. 2016, 67, 1493–1504. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Geng, S.; Wang, D.; Feng, N.; Zhang, D.; Wu, L.; Hao, C.; Zhang, X.; Li, A.; Mao, L. Characterization of Squamosa Promoter Binding Protein-Like genes in wheat. J. Plant Biol. 2015, 58, 220–229. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, Y.; Kohalmi, S.E.; Amyot, L.; Hannoufa, A. SQUAMOSA PROMOTER BINDING PROTEIN-LIKE 2 controls floral organ development and plant fertility by activating ASYMMETRIC LEAVES 2 in Arabidopsis thaliana. Plant Mol. Biol. 2016, 92, 661–674. [Google Scholar] [CrossRef]
- Frazier, T.P.; Xie, F.; Freistaedter, A.; Burklew, C.E.; Zhang, B. Identification and characterization of microRNAs and their target genes in tobacco (Nicotiana tabacum). Planta 2010, 232, 1289–1308. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Wang, T.; Hao, R.; Pan, H.; Cheng, T.; Zhang, Q. Selection of Suitable Reference Genes for Quantitative Real-time Polymerase Chain Reaction in Prunus mume during Flowering Stages and under Different Abiotic Stress Conditions. J. Am. Soc. Hortic. Sci. 2014, 139, 113–122. [Google Scholar] [CrossRef] [Green Version]
- Eli, C.; Ezhang, Y.; Ezhang, K.; Eguo, D.; Ecui, B.; Ewang, X.; Ehuang, X. Promoting flowering, lateral shoot outgrowth, leaf development, and flower abscission in tobacco plants overexpressing cotton FLOWERING LOCUS T (FT)-like gene GhFT1. Front. Plant Sci. 2015, 6, 454. [Google Scholar] [CrossRef] [Green Version]
- Liao, W.-Y.; Lin, L.-F.; Lin, M.-D.; Hsieh, S.-C.; Li, A.Y.-S.; Tsay, Y.-S.; Chou, M.-L. Overexpression of Lilium formosanum MADS-box (LFMADS) Causing Floral Defects While Promoting Flowering in Arabidopsis thaliana, Whereas Only Affecting Floral Transition Time in Nicotiana tabacum. Int. J. Mol. Sci. 2018, 19, 2217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, T.; Li, S.; Zang, L.; Dai, L.; Yang, C.; Qu, G.-Z. Overexpression of Two PsnAP1 Genes from Populus simonii × P. nigra Causes Early Flowering in Transgenic Tobacco and Arabidopsis. PLoS ONE 2014, 9, e111725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Maleki, S.S.; Mohammadi, K.; Ji, K. Influencing factors of genetic transformation efficiency in tobacco (Nicotiana tabacum ‘NC89′) using Agrobacterium tumefacience. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2018, 42, 27–34. [Google Scholar]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yong, X.; Zheng, T.; Han, Y.; Cong, T.; Li, P.; Liu, W.; Ding, A.; Cheng, T.; Wang, J.; Zhang, Q. The miR156-Targeted SQUAMOSA PROMOTER BINDING PROTEIN (PmSBP) Transcription Factor Regulates the Flowering Time by Binding to the Promoter of SUPPRESSOR OF OVEREXPRESSION OF CO1 (PmSOC1) in Prunus mume. Int. J. Mol. Sci. 2022, 23, 11976. https://doi.org/10.3390/ijms231911976
Yong X, Zheng T, Han Y, Cong T, Li P, Liu W, Ding A, Cheng T, Wang J, Zhang Q. The miR156-Targeted SQUAMOSA PROMOTER BINDING PROTEIN (PmSBP) Transcription Factor Regulates the Flowering Time by Binding to the Promoter of SUPPRESSOR OF OVEREXPRESSION OF CO1 (PmSOC1) in Prunus mume. International Journal of Molecular Sciences. 2022; 23(19):11976. https://doi.org/10.3390/ijms231911976
Chicago/Turabian StyleYong, Xue, Tangchun Zheng, Yu Han, Tianci Cong, Ping Li, Weichao Liu, Aiqing Ding, Tangren Cheng, Jia Wang, and Qixiang Zhang. 2022. "The miR156-Targeted SQUAMOSA PROMOTER BINDING PROTEIN (PmSBP) Transcription Factor Regulates the Flowering Time by Binding to the Promoter of SUPPRESSOR OF OVEREXPRESSION OF CO1 (PmSOC1) in Prunus mume" International Journal of Molecular Sciences 23, no. 19: 11976. https://doi.org/10.3390/ijms231911976
APA StyleYong, X., Zheng, T., Han, Y., Cong, T., Li, P., Liu, W., Ding, A., Cheng, T., Wang, J., & Zhang, Q. (2022). The miR156-Targeted SQUAMOSA PROMOTER BINDING PROTEIN (PmSBP) Transcription Factor Regulates the Flowering Time by Binding to the Promoter of SUPPRESSOR OF OVEREXPRESSION OF CO1 (PmSOC1) in Prunus mume. International Journal of Molecular Sciences, 23(19), 11976. https://doi.org/10.3390/ijms231911976