
RAI14 Promotes Melanoma Progression by Regulating the FBXO32/c-MYC Pathway

 and
and 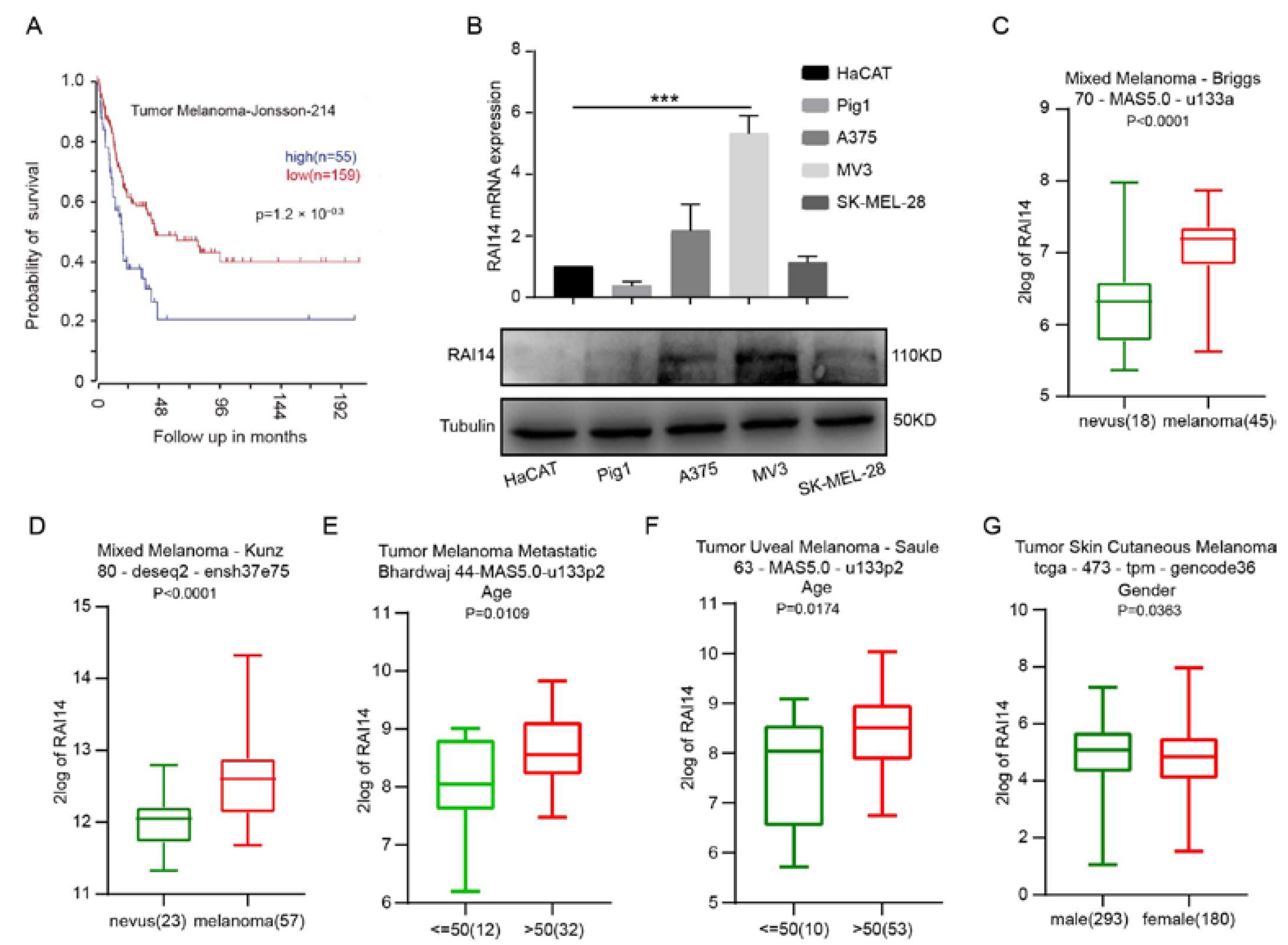{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. High Expression of RAI14 Is Associated with Poor Prognosis of Melanoma
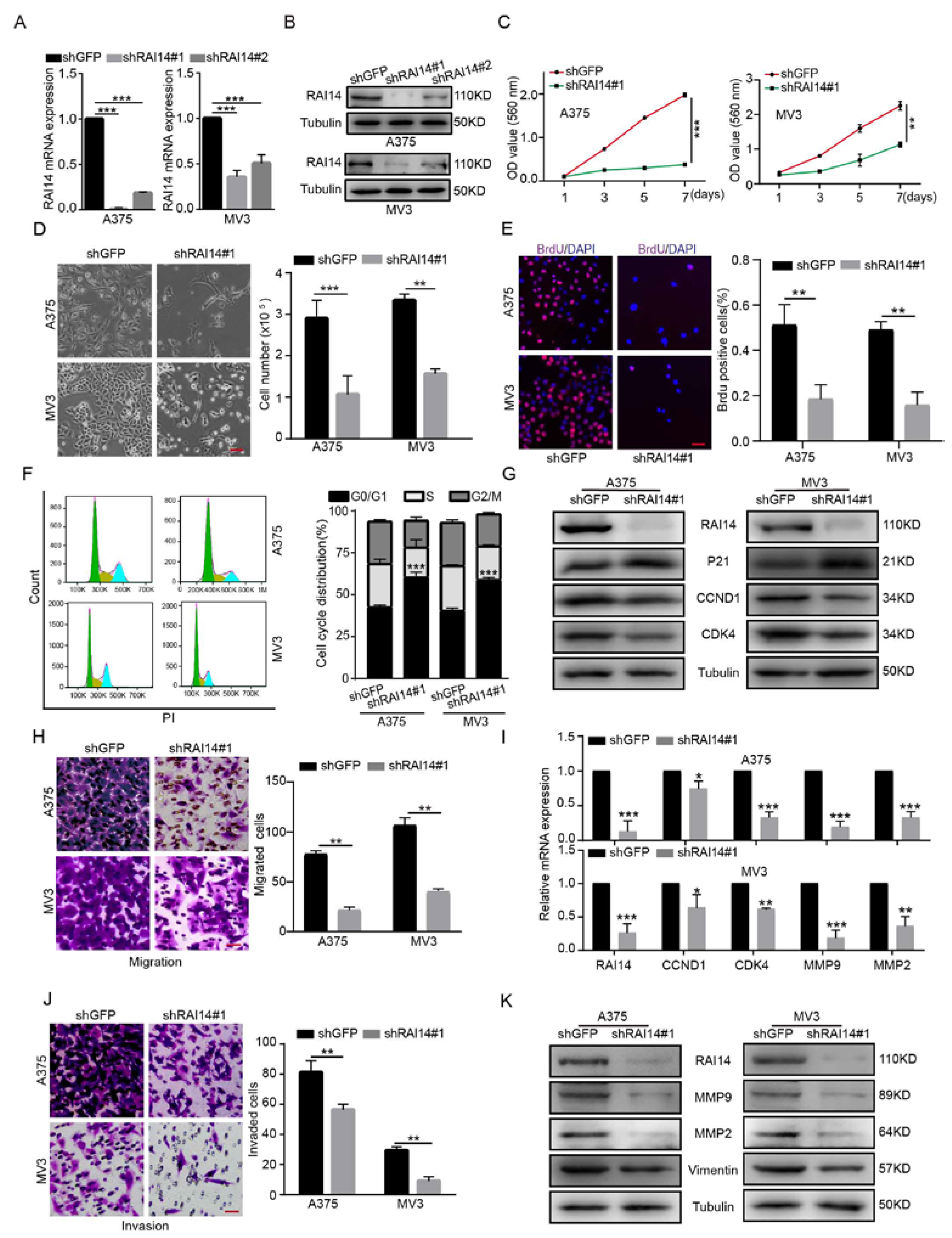
2.2. RAI14 Knockdown Inhibited the Cell Proliferation, Migration and Invasion of Melanoma Cells
2.3. RAI14 Recovery Restored the Cell Proliferation and Migration of RAI14 Knockdown Melanoma Cells
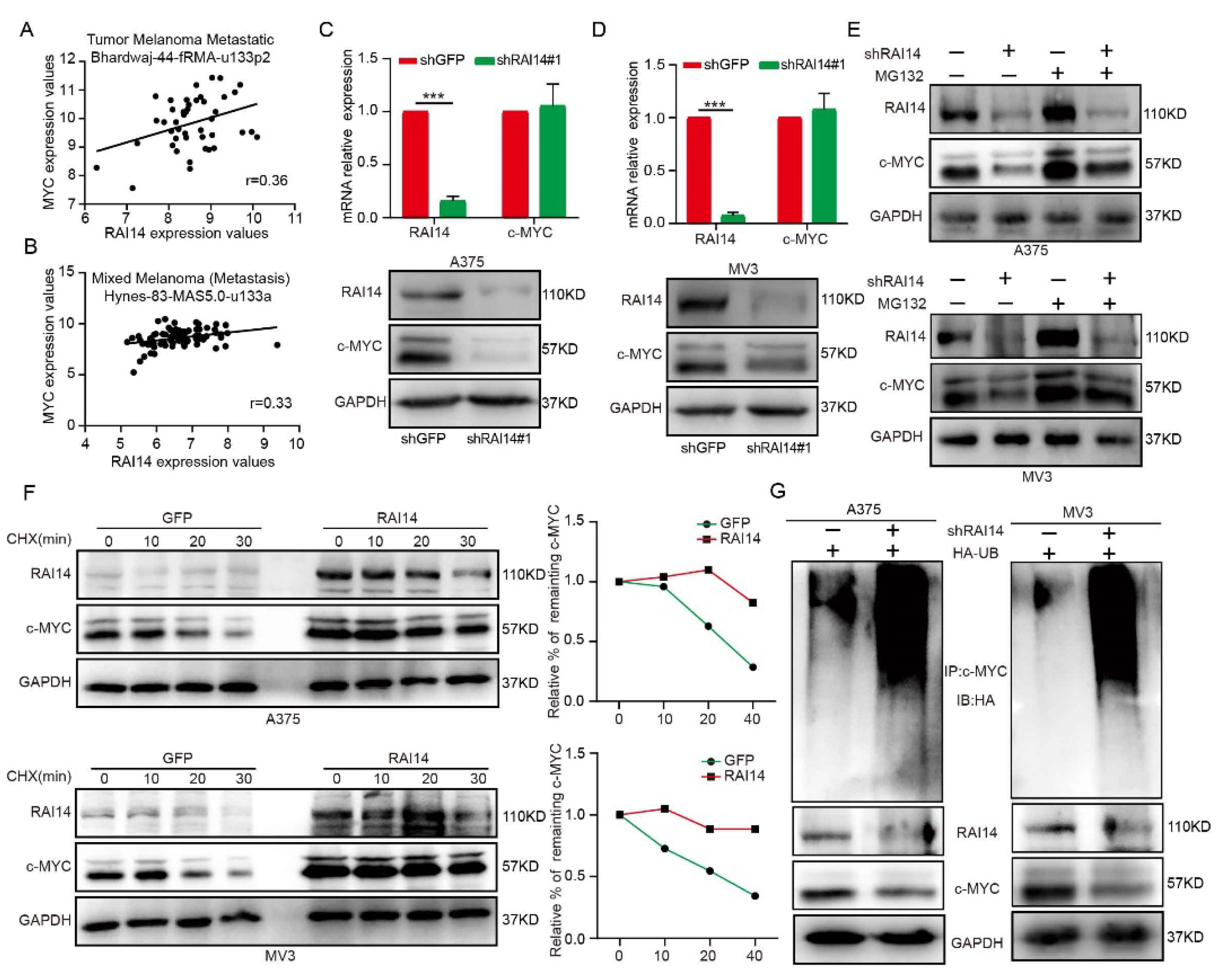
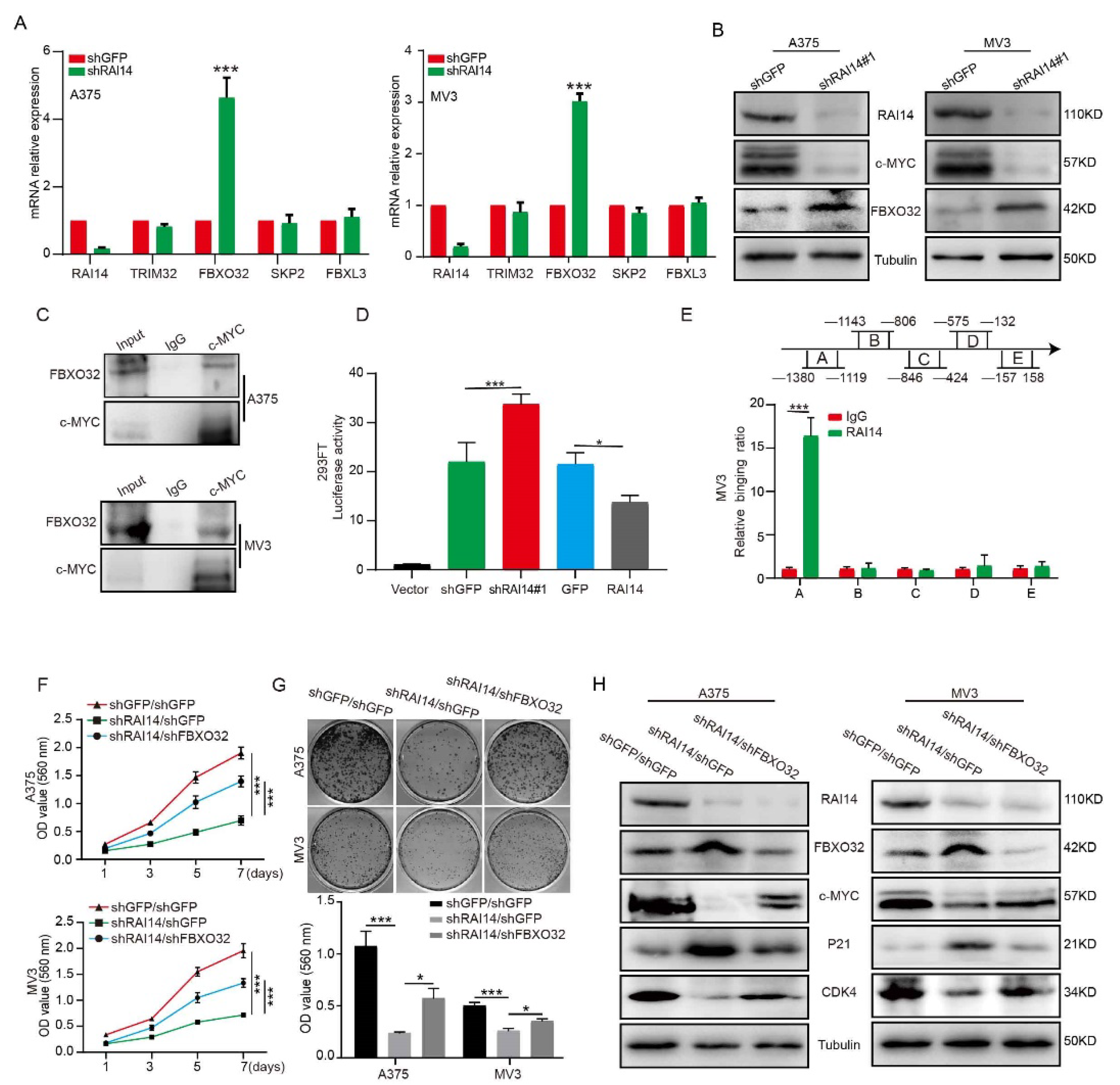
2.4. RAI14 Regulates the Stability of c-MYC by Regulating c-MYC Ubiquitination
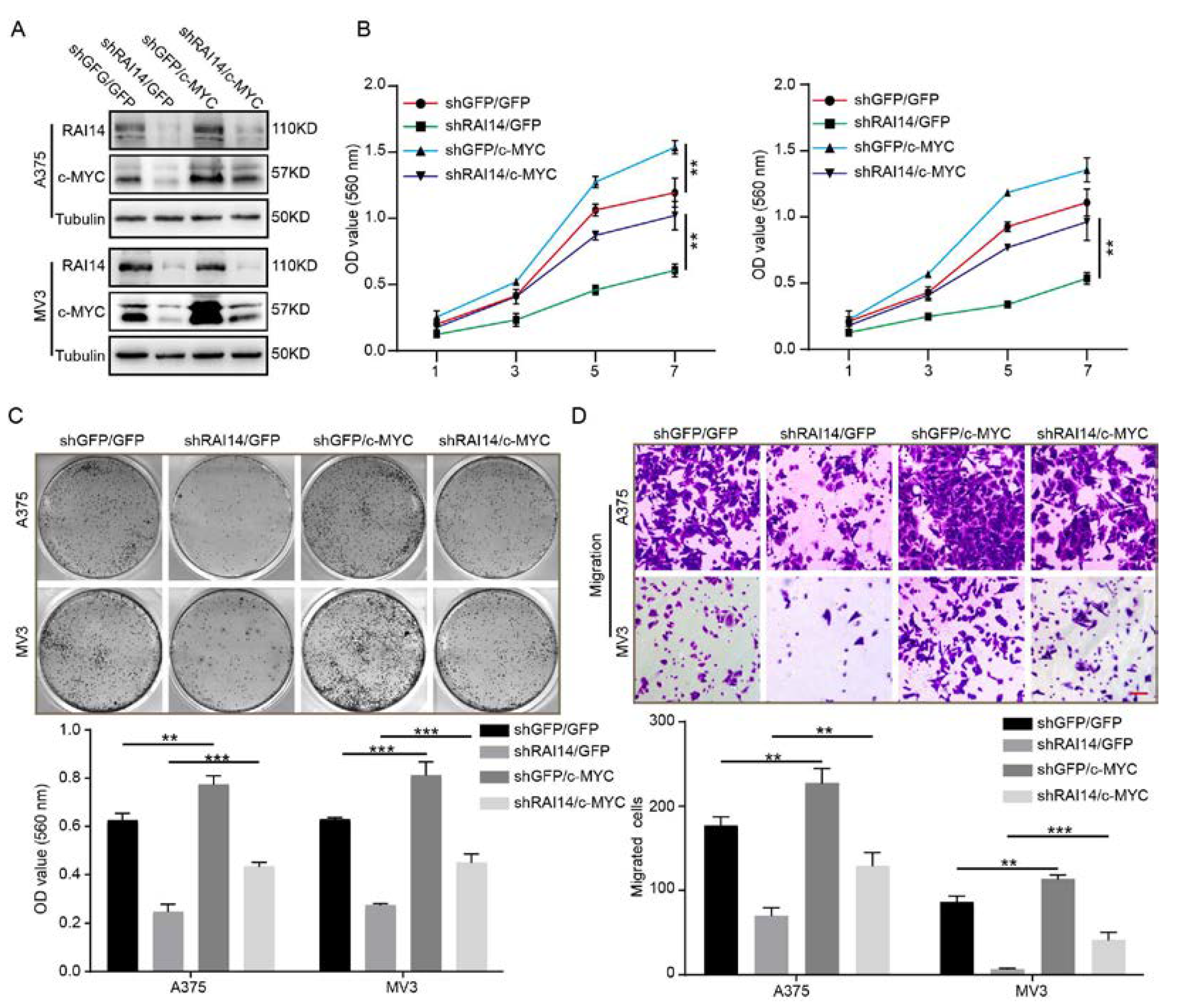
2.5. The Overexpression of c-MYC Significantly Restored Cell Proliferation and Migration of RAI14 Knockdown Melanoma Cells
2.6. RAI14 Suppressed the Transcription of FBXO32
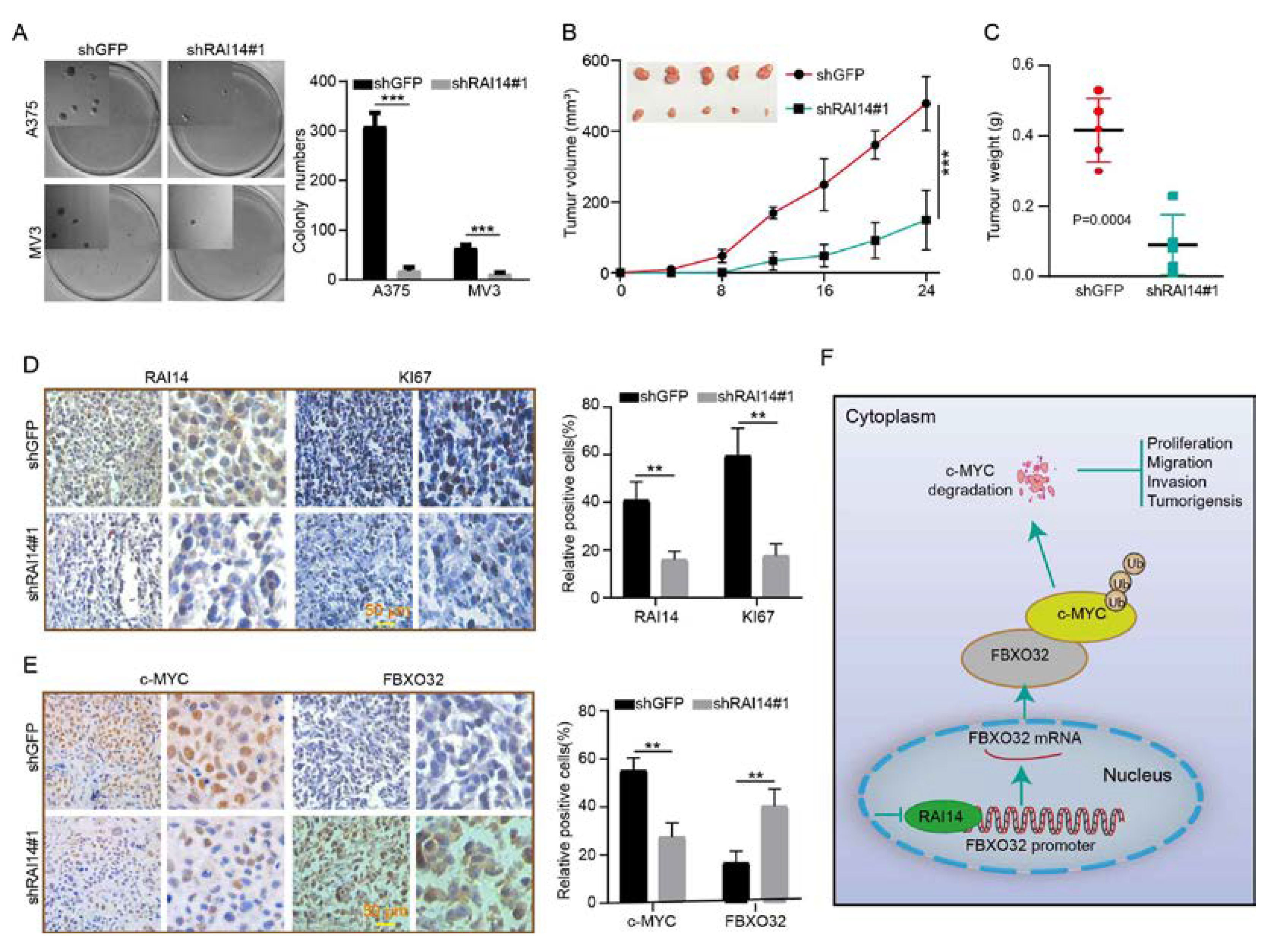
2.7. RAI14 Knockdown Inhibits Tumor Growth and Improved Prognosis in Mice
3. Discussion
4. Materials and Methods
4.1. Cell Lines, Antibodies, and Reagents
4.2. Transfection and Infection
4.3. Quantitative Real-Time RT-PCR Assay
4.4. Cell Proliferation Assay (MTT Assay)
4.5. BrdU Assay
4.6. Flow Cytometry
4.7. Plate Clone Formation and Soft Agar Assays
4.8. Transwell Assay
4.9. Western Blot Analysis
4.10. Ubiquitination and Turnover Assay
4.11. Dual-Luciferase Reporter Assay
4.12. Chromatin Immunoprecipitation
4.13. Xenograft Assay
4.14. Immunohistochemistry
4.15. Patient Data Analysis
4.16. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Rastrelli, M.; Tropea, S.; Rossi, C.R.; Alaibac, M. Melanoma: Epidemiology, risk factors, pathogenesis, diagnosis and classification. In Vivo 2014, 28, 1005–1011. [Google Scholar] [PubMed]
- Eddy, K.; Chen, S. Overcoming Immune Evasion in Melanoma. Int. J. Mol. Sci. 2020, 21, 8984. [Google Scholar] [CrossRef]
- Ahmed, B.; Qadir, M.I.; Ghafoor, S. Malignant Melanoma: Skin Cancer-Diagnosis, Prevention, and Treatment. Crit. Rev. Eukaryot. Gene Expr. 2020, 30, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Kutty, R.K.; Kutty, G.; Samuel, W.; Duncan, T.; Bridges, C.C.; El-Sherbeeny, A.; Nagineni, C.N.; Smith, S.B.; Wiggert, B. Molecular characterization and developmental expression of NORPEG, a novel gene induced by retinoic acid. J. Biol. Chem. 2001, 276, 2831–2840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, W.Y.; Sharpless, N.E. The regulation of INK4/ARF in cancer and aging. Cell 2006, 127, 265–275. [Google Scholar] [CrossRef] [Green Version]
- Kohda, A.; Yamazaki, S.; Sumimoto, H. The Nuclear Protein IκBζ Forms a Transcriptionally Active Complex with Nuclear Factor-κB (NF-κB) p50 and the Lcn2 Promoter via the N- and C-terminal Ankyrin Repeat Motifs. J. Biol. Chem. 2016, 291, 20739–20752. [Google Scholar] [CrossRef] [Green Version]
- Mosavi, L.K.; Cammett, T.J.; Desrosiers, D.C.; Peng, Z.Y. The ankyrin repeat as molecular architecture for protein recognition. Protein Sci. 2004, 13, 1435–1448. [Google Scholar] [CrossRef] [Green Version]
- Voronin, D.A.; Kiseleva, E.V. Functional role of proteins containing ankyrin repeats. Tsitologiia 2007, 49, 989–999. [Google Scholar] [CrossRef]
- Yuan, C.; Hu, H.; Kuang, M.; Chen, Z.; Tao, X.; Fang, S.; Sun, Y.; Zhang, Y.; Chen, H. Super enhancer associated RAI14 is a new potential biomarker in lung adenocarcinoma. Oncotarget 2017, 8, 105251–105261. [Google Scholar] [CrossRef]
- Gu, M.; Zheng, W.; Zhang, M.; Dong, X.; Zhao, Y.; Wang, S.; Jiang, H.; Liu, L.; Zheng, X. Downregulation of RAI14 inhibits the proliferation and invasion of breast cancer cells. J. Cancer 2019, 10, 6341–6348. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Zhang, H.; Du, G.; Meng, X.; Wu, T.; Zhou, Q.; Wang, Y.; Tan, B. RAI14 Is a Prognostic Biomarker and Correlated With Immune Cell Infiltrates in Gastric Cancer. Technol. Cancer Res. Treat. 2020, 19, 1533033820970684. [Google Scholar] [CrossRef]
- Wang, J.; Cai, Y.; Luo, J.; Sun, Z.; Yu, J.; Yan, F.; He, X. RAI14 silencing suppresses progression of esophageal cancer via the STAT3 pathway. Aging 2020, 12, 18084–18098. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.C.; Chen, H.Y.; Yuan, S.; Yu, S.L.; Lin, C.H.; Wu, G.; Yang, P.C.; Li, K.C. Genome-wide analysis of three-way interplay among gene expression, cancer cell invasion and anti-cancer compound sensitivity. BMC Med. 2013, 11, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dang, C.V. MYC on the path to cancer. Cell 2012, 149, 22–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dang, C.V.; Le, A.; Gao, P. MYC-induced cancer cell energy metabolism and therapeutic opportunities. Clin. Cancer Res. 2009, 15, 6479–6483. [Google Scholar] [CrossRef] [Green Version]
- Thompson, E.B. The many roles of c-Myc in apoptosis. Annu. Rev. Physiol. 1998, 60, 575–600. [Google Scholar] [CrossRef] [PubMed]
- Mei, Z.; Zhang, D.; Hu, B.; Wang, J.; Shen, X.; Xiao, W. FBXO32 Targets c-Myc for Proteasomal Degradation and Inhibits c-Myc Activity. J. Biol. Chem. 2015, 290, 16202–16214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, N.; Penn, L.Z. Reflecting on 25 years with MYC. Nat. Rev. Cancer 2008, 8, 976–990. [Google Scholar] [CrossRef] [PubMed]
- He, X.Y.; Zhao, J.; Chen, Z.Q.; Jin, R.; Liu, C.Y. High Expression of Retinoic Acid Induced 14 (RAI14) in Gastric Cancer and Its Prognostic Value. Med. Sci. Monit. 2018, 24, 2244–2251. [Google Scholar] [CrossRef] [PubMed]
- Meng, C.; Xia, S.; He, Y.; Tang, X.; Zhang, G.; Zhou, T. Discovery of Prognostic Signature Genes for Overall Survival Prediction in Gastric Cancer. Comput. Math. Methods Med. 2020, 2020, 5479279. [Google Scholar] [CrossRef] [PubMed]
- Mansour, M.A. Ubiquitination: Friend and foe in cancer. Int. J. Biochem. Cell Biol. 2018, 101, 80–93. [Google Scholar] [CrossRef] [PubMed]
- Faktor, J.; Pjechová, M.; Hernychová, L.; Vojtěšek, B. Protein Ubiquitination Research in Oncology. Klin. Onkol. 2019, 32 (Suppl. S3), 56–64. [Google Scholar] [CrossRef] [Green Version]
- Popovic, D.; Vucic, D.; Dikic, I. Ubiquitination in disease pathogenesis and treatment. Nat. Med. 2014, 20, 1242–1253. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Dong, Z.; Cui, H. Interplay between Epigenetics and Cellular Metabolism in Colorectal Cancer. Biomolecules 2021, 11, 1406. [Google Scholar] [CrossRef]
- Zhang, G.; Zhu, Q.; Fu, G.; Hou, J.; Hu, X.; Cao, J.; Peng, W.; Wang, X.; Chen, F.; Cui, H. TRIP13 promotes the cell proliferation, migration and invasion of glioblastoma through the FBXW7/c-MYC axis. Br. J. Cancer 2019, 121, 1069–1078. [Google Scholar] [CrossRef] [PubMed]
- Kutty, R.K.; Chen, S.; Samuel, W.; Vijayasarathy, C.; Duncan, T.; Tsai, J.Y.; Fariss, R.N.; Carper, D.; Jaworski, C.; Wiggert, B. Cell density-dependent nuclear/cytoplasmic localization of NORPEG (RAI14) protein. Biochem. Biophys. Res. Commun. 2006, 345, 1333–1341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kane, E.I.; Spratt, D.E. Structural Insights into Ankyrin Repeat-Containing Proteins and Their Influence in Ubiquitylation. Int. J. Mol. Sci. 2021, 22, 609. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Xia, Y.; Wu, Y.; Huang, S.; Teng, Y.; Liu, X.; Li, P.; Chen, J.; Zhuang, J. Ankyrin repeat domain 1: A novel gene for cardiac septal defects. J. Gene Med. 2019, 21, e3070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michel, F.; Soler-Lopez, M.; Petosa, C.; Cramer, P.; Siebenlist, U.; Müller, C.W. Crystal structure of the ankyrin repeat domain of Bcl-3: A unique member of the IkappaB protein family. EMBO J. 2001, 20, 6180–6190. [Google Scholar] [CrossRef] [PubMed]
- Kojic, S.; Radojkovic, D.; Faulkner, G. Muscle ankyrin repeat proteins: Their role in striated muscle function in health and disease. Crit. Rev. Clin. Lab. Sci. 2011, 48, 269–294. [Google Scholar] [CrossRef] [PubMed]
- Shi, P.; Xu, J.; Zhao, X.; Shen, P.; Wen, D.; Yu, Q.; Deng, Y.; Shi, D.; Lu, F. CK1 inhibitor affects in vitro maturation and developmental competence of bovine oocytes. Reprod. Domest. Anim. Zuchthyg. 2019, 54, 1104–1112. [Google Scholar] [CrossRef]
- Hou, J.; Xu, M.; Gu, H.; Pei, D.; Liu, Y.; Huang, P.; Chang, H.; Cui, H. ZC3H15 promotes glioblastoma progression through regulating EGFR stability. Cell Death Disease 2022, 13, 55. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, J.; Shi, P.; Xia, F.; Zhao, X.; Chen, J.; Geng, R.; Cui, H.; Yang, L. RAI14 Promotes Melanoma Progression by Regulating the FBXO32/c-MYC Pathway. Int. J. Mol. Sci. 2022, 23, 12036. https://doi.org/10.3390/ijms231912036
Xu J, Shi P, Xia F, Zhao X, Chen J, Geng R, Cui H, Yang L. RAI14 Promotes Melanoma Progression by Regulating the FBXO32/c-MYC Pathway. International Journal of Molecular Sciences. 2022; 23(19):12036. https://doi.org/10.3390/ijms231912036
Chicago/Turabian StyleXu, Jie, Pengfei Shi, Fanwei Xia, Xuan Zhao, Junfan Chen, Rui Geng, Hongjuan Cui, and Liqun Yang. 2022. "RAI14 Promotes Melanoma Progression by Regulating the FBXO32/c-MYC Pathway" International Journal of Molecular Sciences 23, no. 19: 12036. https://doi.org/10.3390/ijms231912036
APA StyleXu, J., Shi, P., Xia, F., Zhao, X., Chen, J., Geng, R., Cui, H., & Yang, L. (2022). RAI14 Promotes Melanoma Progression by Regulating the FBXO32/c-MYC Pathway. International Journal of Molecular Sciences, 23(19), 12036. https://doi.org/10.3390/ijms231912036