
Disruption of Circadian Rhythm Genes in Obstructive Sleep Apnea Patients—Possible Mechanisms Involved and Clinical Implication

 , ,
, ,  , and
, and
Abstract
:1. Master Circadian Clock and Influence of Light–Night Cycle
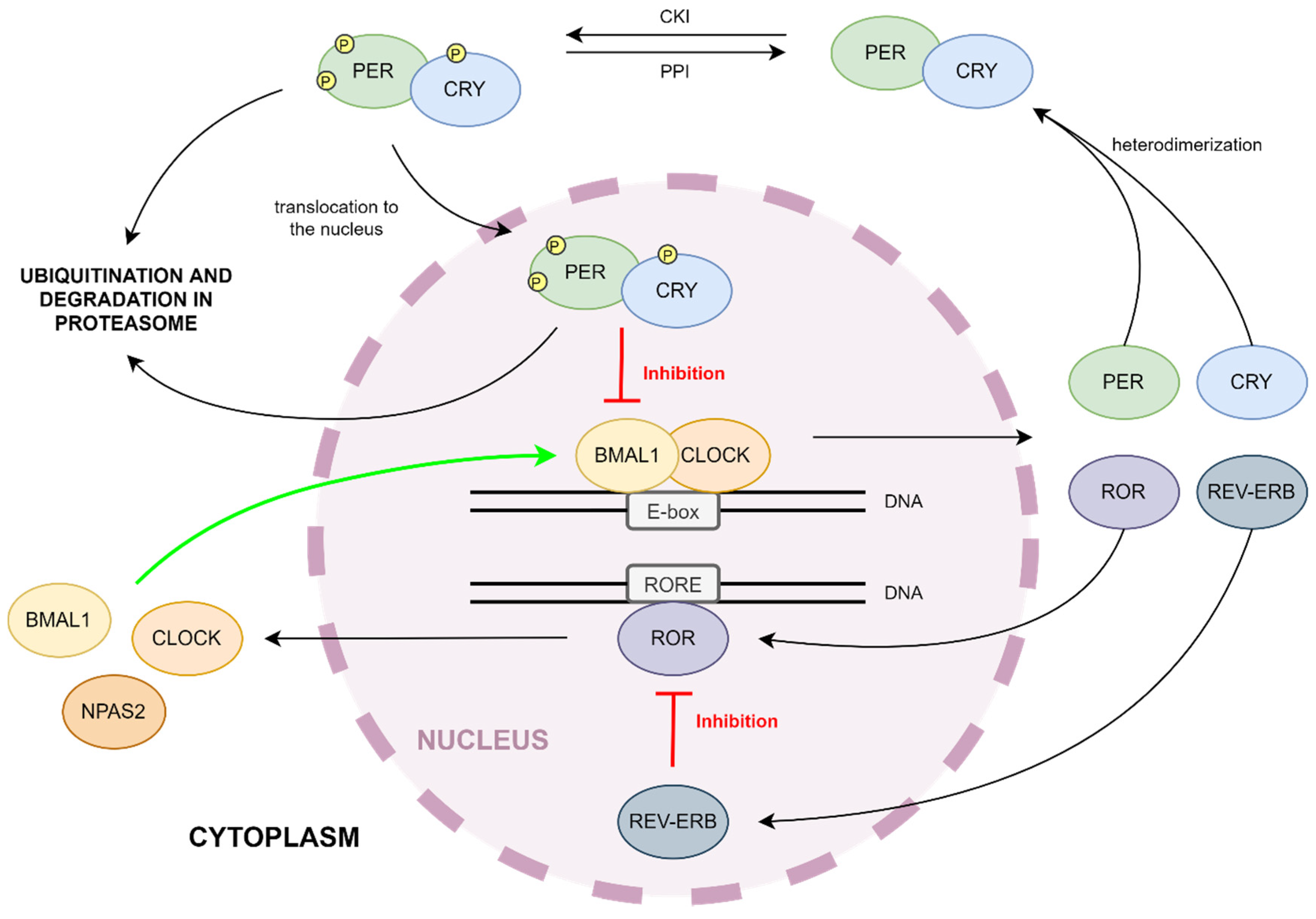
2. Molecular Mechanism of the Circadian Clock
3. Impact of Kinases and Phosphatases on the Circadian Clock
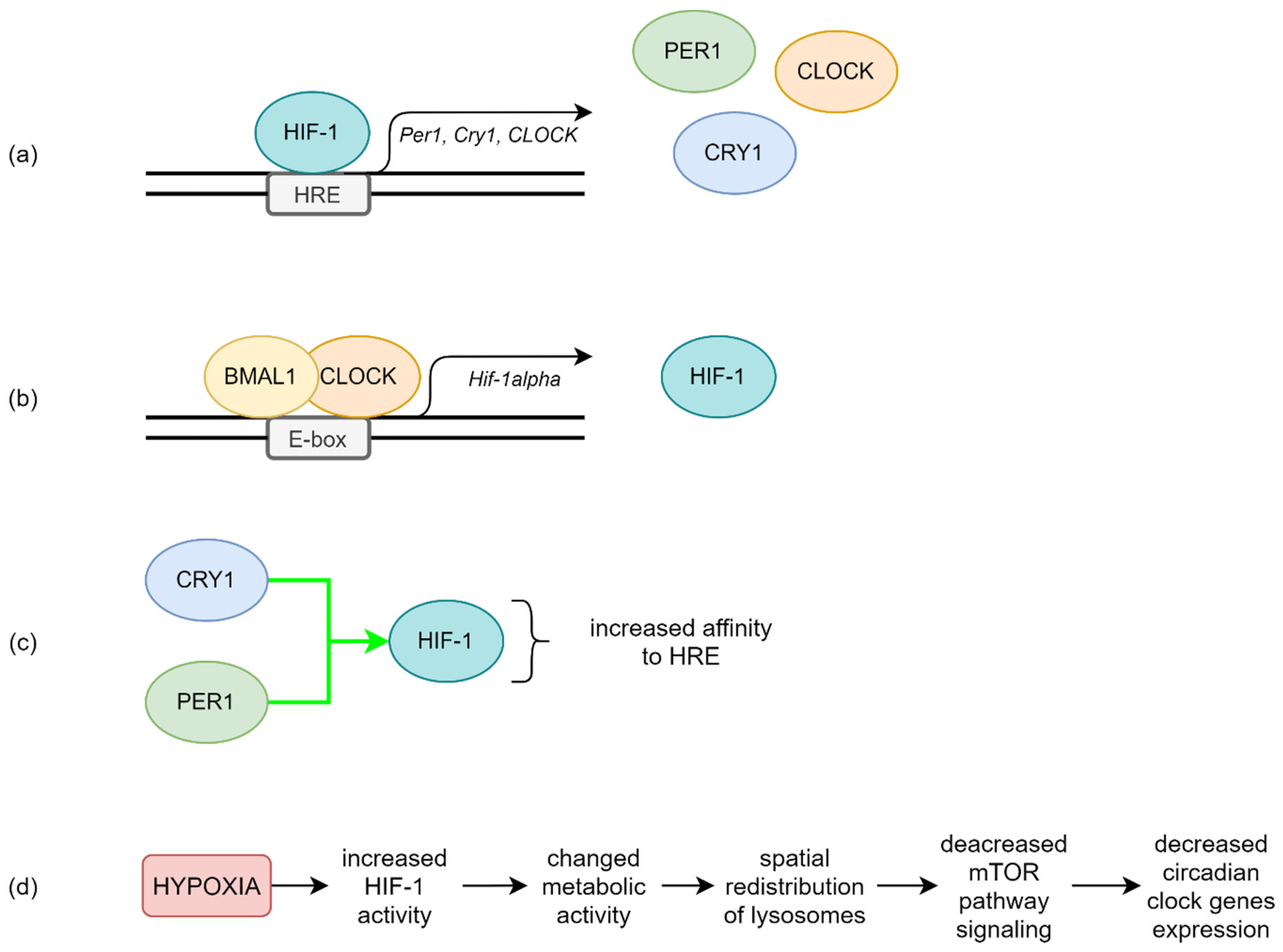
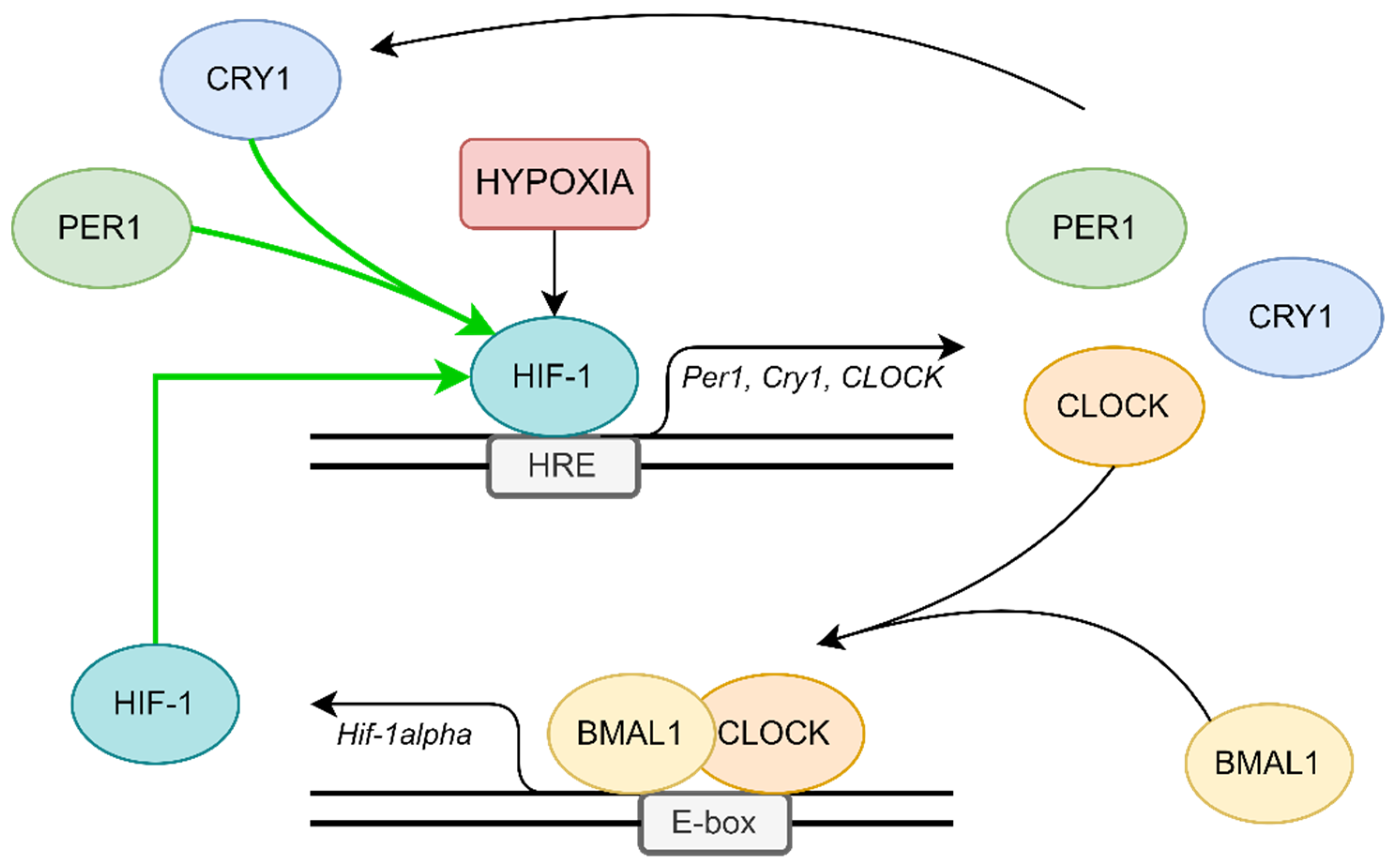
4. Possible Molecular Mechanisms in OSA
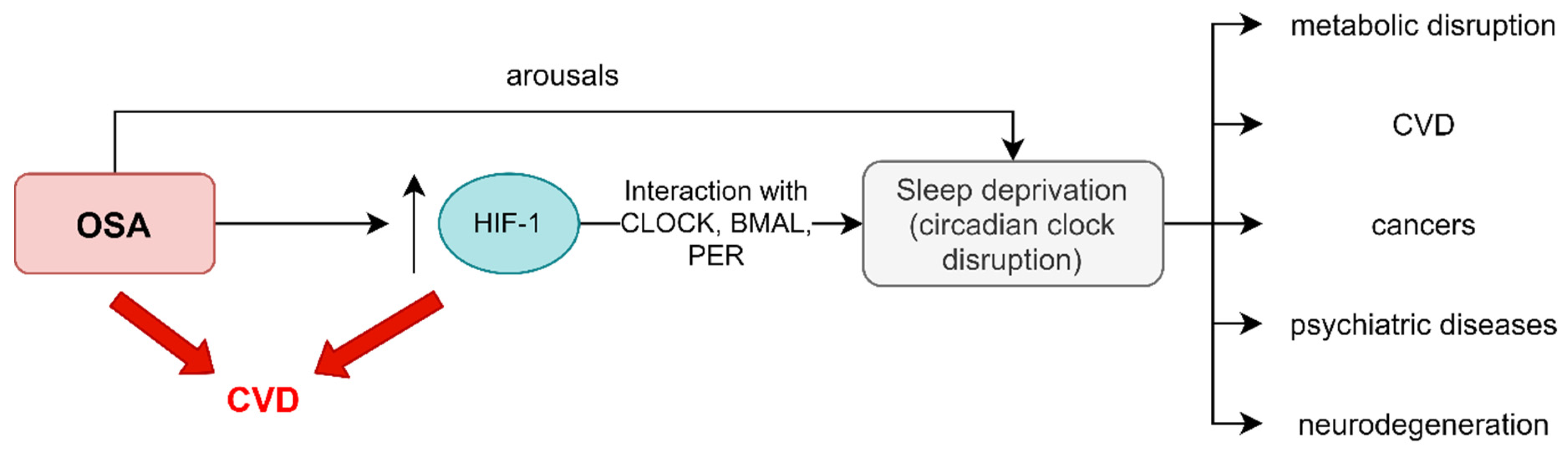
5. Clinical Implications of Circadian Rhythm Gene Disruption in OSA Patients
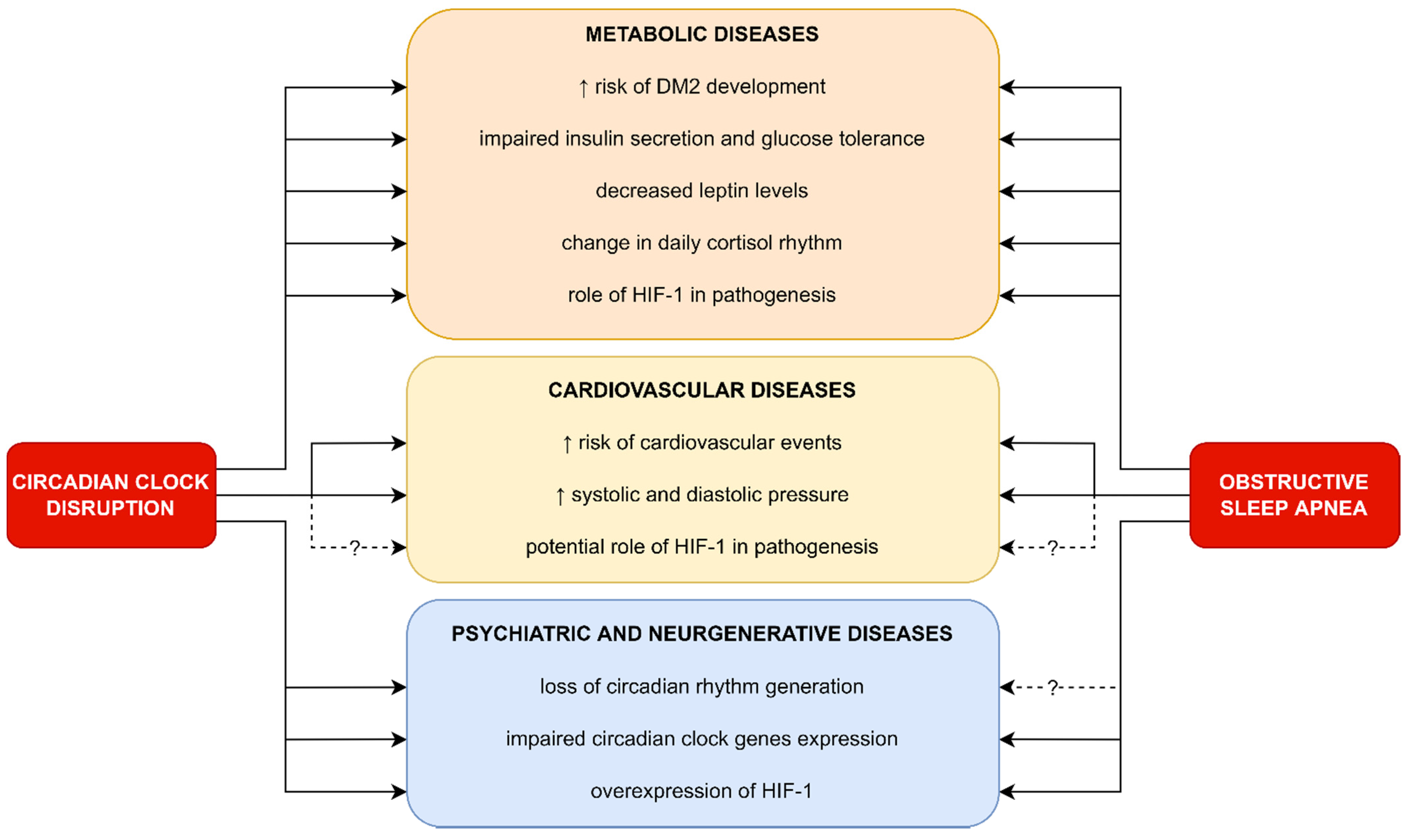
5.1. Metabolic Diseases
5.2. Cardiovascular Diseases
5.3. Psychiatric and Neurodegenerative Diseases

6. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AMPK | adenosine monophosphate-activated protein kinase |
| bHLH-PAS | basic helix-loop-helix PER-ARNT-SIM |
| BMAL1/ARNTL | brain and muscle ARNT-like 1/aryl hydrocarbon receptor nuclear translocator like |
| CamK | calcium/calmodulin-dependent protein kinases |
| cAMP | cyclic adenosine monophosphate; |
| CKI | casein kinases |
| CLOCK | clock circadian regulator/circadian locomotor output cycles protein kaput |
| CPAP | continuous positive airway pressure treatment |
| CREB | phosphorylate cAMP-responsive element-binding protein |
| Cry2 | cryptochrome 2 |
| CVD | cardio-vascular disease |
| DBP | albumin d-element binding protein |
| E4BP4 | E4 binding protein 4 |
| E-box | enhancer box; GSK3β—Glycogen synthase kinase 3β |
| HIF | hypoxia inducible factor |
| HMG-CoA | 3-hydroxy-3-methylglutaryl-coenzyme A synthase |
| HRE | hypoxia response element |
| IH | intermittent hypoxia |
| LDHA | lactate dehydrogenase A |
| MAPK | mitogen-activated protein kinases |
| mTOR | mechanistic target of rapamycin kinase |
| NMDAR | N-methyl-d-aspartate receptors |
| NPAS2 | neuronal PAS domain protein 2 |
| OSA | obstructive sleep apnea |
| PACAP | pituitary adenylate cyclase—activating polypeptide |
| PER1 | period protein 1 |
| PER2 | period protein 2 |
| PER3 | period protein 3 |
| PHD3 | prolyl hydroxylase 3 |
| PKA | protein kinase A |
| PP1 | protein phosphatase 1 |
| PP5 | protein phosphatase 5 |
| REV-ERBα | nuclear receptor subfamily 1 group D member 1 |
| ROR | RAR-related orphan receptor |
| RORE | ROR response elements |
| RORα | Nuclear retinoid-related orphan receptors α |
| SCF-Fbxl3 | E3 ubiquitin ligase complex |
| E3 | ubiquitin ligase complex |
| SCN | suprachiasmatic nucleus |
| T2DM | diabetes mellitus type 2 |
| VEGF | vascular endothelial growth factor |
References
- Partch, C.; Green, C.; Takahashi, J. Molecular architecture of the mammalian circadian clock. Trends Cell Biol. 2014, 24, 90–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welsh, D.; Takahashi, J.; Kay, S. Suprachiasmatic Nucleus: Cell Autonomy and Network Properties. Annu. Rev. Physiol. 2010, 72, 551–577. [Google Scholar] [CrossRef] [Green Version]
- Morin, L.; Allen, C. The circadian visual system. Brain Res. Rev. 2006, 51, 1–60. [Google Scholar] [CrossRef]
- Ozge, G.; Dogan, D.; Koylu, M.T.; Ayyildiz, O.; Akincioglu, D.; Mumcuoglu, T.; Mutlu, F.M. Retina nerve fiber layer and choroidal thickness changes in obstructive sleep apnea syndrome. Postgrad. Med. 2016, 128, 317–322. [Google Scholar] [CrossRef]
- Kraves, S.; Weitz, C. A role for cardiotrophin-like cytokine in the circadian control of mammalian locomotor activity. Nat. Neurosci. 2006, 9, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Meijer, J.H.; Schwartz, W.J. In search of the pathways for light-induced pacemaker resetting in the suprachiasmatic nucleus. J. Biol. Rhythms 2003, 18, 235–249. [Google Scholar] [CrossRef]
- Ginty, D.; Kornhauser, J.; Thompson, M.; Bading, H.; Mayo, K.; Takahashi, J.; Greenberg, M. Regulation of CREB phosphorylation in the suprachiasmatic nucleus by light and a circadian clock. Science 1993, 260, 238–241. [Google Scholar] [CrossRef]
- Travnickova-Bendova, Z.; Cermakian, N.; Reppert, S.; Sassone-Corsi, P. Bimodal regulation of mPeriod promoters by CREB-dependent signaling and CLOCK/BMAL1 activity. Proc. Natl. Acad. Sci. USA 2002, 99, 7728–7733. [Google Scholar] [CrossRef] [Green Version]
- Aton, S.; Herzog, E. Come Together, Right Now: Synchronization of Rhythms in a Mammalian Circadian Clock. Neuron 2005, 48, 531–534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, T.; Yamada, R.; Ukai, H.; Baggs, J.; Miraglia, L.; Kobayashi, T.; Welsh, D.; Kay, S.; Ueda, H.; Hogenesch, J. Feedback repression is required for mammalian circadian clock function. Nat. Genet. 2006, 38, 312–319. [Google Scholar] [CrossRef] [PubMed]
- Reischl, S.; Kramer, A. Kinases, and phosphatases in the mammalian circadian clock. FEBS Lett. 2011, 585, 1393–1399. [Google Scholar] [CrossRef] [Green Version]
- Huang, N.; Chelliah, Y.; Shan, Y.; Taylor, C.; Yoo, S.; Partch, C.; Green, C.B.; Zhang, H.; Takahashi, J.S. Crystal Structure of the Heterodimeric CLOCK: BMAL1 Transcriptional Activator Complex. Science 2012, 337, 189–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallego, M.; Virshup, D. Post-translational modifications regulate the ticking of the circadian clock. Nat. Rev. Mol. Cell Biol. 2007, 8, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Freeman, S.; Kwon, H.; Portolano, N.; Parkin, G.; Venkatraman Girija, U.; Basran, J.; Fielding, A.J.; Fairall, L.; Svistunenko, D.A.; Moody, P.C.E.; et al. Heme binding to human CLOCK affects interactions with the E-box. Proc. Natl. Acad. Sci. USA 2019, 116, 19911–19916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, S.; Mohawk, J.; Siepka, S.; Shan, Y.; Huh, S.K.; Hong, H.K.; Kornblum, I.; Kumar, V.; Koike, N.; Xu, M.; et al. Competing E3 Ubiquitin Ligases Govern Circadian Periodicity by Degradation of CRY in Nucleus and Cytoplasm. Cell 2013, 152, 1091–1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Challet, E. The circadian regulation of food intake. Nat. Rev. Endocrinol. 2019, 15, 393–405. [Google Scholar] [CrossRef]
- Reinke, H.; Asher, G. Crosstalk between metabolism and circadian clocks. Nat. Rev. Mol. Cell Biol. 2019, 20, 227–241. [Google Scholar] [CrossRef]
- Zhu, L.; Zee, P. Circadian Rhythm Sleep Disorders. Neurol. Clin. 2012, 30, 1167–1191. [Google Scholar] [CrossRef] [Green Version]
- Nam, D.; Guo, B.; Chatterjee, S.; Chen, M.; Nelson, D.; Yechoor, V.K.; Ma, K. The adipocyte clock controls brown adipogenesis via TGF-β/BMP signaling pathway. J. Cell Sci. 2015, 128, 1835–1847. [Google Scholar]
- Douma, L.; Gumz, M. Circadian clock-mediated regulation of blood pressure. Free Radic. Biol. Med. 2018, 119, 108–114. [Google Scholar] [CrossRef]
- Hergenhan, S.; Holtkamp, S.; Scheiermann, C. Molecular Interactions Between Components of the Circadian Clock and the Immune System. J. Mol. Biol. 2020, 432, 3700–3713. [Google Scholar] [CrossRef]
- Portaluppi, F.; Tiseo, R.; Smolensky, M.; Hermida, R.; Ayala, D.; Fabbian, F. Circadian rhythms and cardiovascular health. Sleep Med. Rev. 2012, 16, 151–166. [Google Scholar] [CrossRef] [PubMed]
- Genecards.org. 2021. Available online: https://www.genecards.org/ (accessed on 18 September 2021).
- Guillaumond, F.; Dardente, H.; Giguère, V.; Cermakian, N. Differential Control of Bmal1 Circadian Transcription by REV-ERB and ROR Nuclear Receptors. J. Biol. Rhythm. 2005, 20, 391–403. [Google Scholar] [CrossRef]
- Debruyne, J. Oscillating perceptions: The ups and downs of the CLOCK protein in the mouse circadian system. J. Genet. 2008, 87, 437–446. [Google Scholar] [CrossRef] [Green Version]
- Brown, S.; Kowalska, E.; Dallmann, R. (Re)inventing the Circadian Feedback Loop. Dev. Cell 2012, 22, 477–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Zhong, Z.; Zhong, Y.; Zhang, W.; Wang, H. The Zebrafish Period2 Protein Positively Regulates the Circadian Clock through Mediation of Retinoic Acid Receptor (RAR)-related Orphan Receptor α (Rorα). J. Biol. Chem. 2015, 290, 4367–4382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirota, T.; Lee, J.; Lewis, W.; Zhang, E.; Breton, G.; Liu, X.; Garcia, M.; Peters, E.C.; Etchegaray, J.P.; Traver, D.; et al. High-Throughput Chemical Screen Identifies a Novel Potent Modulator of Cellular Circadian Rhythms and Reveals CKIα as a Clock Regulatory Kinase. PLoS Biol. 2010, 8, e1000559. [Google Scholar] [CrossRef] [Green Version]
- Lévy, P.; Kohler, M.; McNicholas, W.; Barbé, F.; McEvoy, R.D.; Somers, V.K.; Lavie, L.; Pépin, J.L. Obstructive sleep apnoea syndrome. Nat. Rev. Dis. Primers 2015, 1, 1–21. [Google Scholar] [CrossRef]
- Mokros, Ł.; Kuczynski, W.; Gabryelska, A.; Franczak, Ł.; Spałka, J.; Białasiewicz, P. High Negative Predictive Value of Normal Body Mass Index for Obstructive Sleep Apnea in the Lateral Sleeping Position. J. Clin. Sleep Med. 2018, 14, 985–990. [Google Scholar] [CrossRef]
- Dewan, N.; Nieto, F.; Somers, V. Intermittent Hypoxemia and OSA. Chest 2015, 147, 266–274. [Google Scholar] [CrossRef] [Green Version]
- Semenza, G. Hypoxia-Inducible Factors in Physiology and Medicine. Cell 2012, 148, 399–408. [Google Scholar] [CrossRef] [Green Version]
- Kaelin, W.; Ratcliffe, P. Oxygen Sensing by Metazoans: The Central Role of the HIF Hydroxylase Pathway. Mol. Cell 2008, 30, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Semenza, G. Hypoxia-Inducible Factor 1 (HIF-1) Pathway. Sci. STKE 2007, 2007, cm8. [Google Scholar] [CrossRef] [PubMed]
- Chachami, G.; Paraskeva, E.; Mingot, J.; Braliou, G.; Görlich, D.; Simos, G. Transport of hypoxia-inducible factor HIF-1α into the nucleus involves importins 4 and 7. Biochem. Biophys. Res. Commun. 2009, 390, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Adamovich, Y.; Ladeuix, B.; Sobel, J.; Manella, G.; Neufeld-Cohen, A.; Assadi, M.H.; Golik, M.; Kuperman, Y.; Tarasiuk, A.; Koeners, M.P.; et al. Oxygen and Carbon Dioxide Rhythms Are Circadian Clock Controlled and Differentially Directed by Behavioral Signals. Cell Metab. 2019, 29, 1092–1103. [Google Scholar] [CrossRef]
- Adamovich, Y.; Ladeuix, B.; Golik, M.; Koeners, M.P.; Asher, G. Rhythmic Oxygen Levels Reset Circadian Clocks through HIF1α. Cell Metab. 2017, 25, 93–101. [Google Scholar] [CrossRef] [Green Version]
- Manella, G.; Aviram, R.; Bolshette, N.; Muvkadi, S.; Golik, M.; Smith, D.F.; Asher, G. Hypoxia induces a time- and tissue-specific response that elicits intertissue circadian clock misalignment. Proc. Natl. Acad. Sci. USA 2020, 117, 779–786. [Google Scholar] [CrossRef]
- Yu, C.; Yang, S.; Fang, X.; Jiang, J.; Sun, C.; Huang, T. Hypoxia disrupts the expression levels of circadian rhythm genes in hepatocellular carcinoma. Mol. Med. Rep. 2015, 11, 4002–4008. [Google Scholar] [CrossRef]
- Bozek, K.; Kiełbasa, S.M.; Kramer, A.; Herzel, H. Promoter analysis of Mammalian clock-controlled genes. Genome Inf. 2007, 18, 65–74. [Google Scholar]
- Chilov, D.; Hofer, T.; Bauer, C.; Wenger, R.; Gassmann, M. Hypoxia affects expression of circadian genes PER1 and CLOCK in mouse brain. FASEB J. 2001, 15, 2613–2622. [Google Scholar] [CrossRef]
- Okabe, T.; Kumagai, M.; Nakajima, Y.; Shirotake, S.; Kodaira, K.; Oyama, M.; Ueno, M.; Ikeda, M. The impact of HIF1α on the Per2 circadian rhythm in renal cancer cell lines. PLoS ONE 2014, 9, e109693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, M.; Morinibu, A.; Koyasu, S.; Goto, Y.; Hiraoka, M.; Harada, H. A circadian clock gene, PER 2, activates HIF-1 as an effector molecule for recruitment of HIF-1α to promoter regions of its downstream genes. FEBS J. 2017, 284, 3804–3816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semenza, G. Oxygen Sensing, Homeostasis, and Disease. N. Engl. J. Med. 2011, 365, 537–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabryelska, A.; Sochal, M.; Turkiewicz, S.; Białasiewicz, P. Relationship between HIF-1 and Circadian Clock Proteins in Obstructive Sleep Apnea Patients—Preliminary Study. J. Clin. Med. 2020, 9, 1599. [Google Scholar] [CrossRef] [PubMed]
- López-Cano, C.; Gutiérrez-Carrasquilla, L.; Barbé, F.; Sánchez, E.; Hernández, M.; Martí, R.; Ceperuelo-Mallafre, V.; Dalmases, M.; Fernández-Veledo, S.; Vendrell, J.; et al. Effect of Type 2 Diabetes Mellitus on the Hypoxia-Inducible Factor 1-Alpha Expression. Is There a Relationship with the Clock Genes? J. Clin. Med. 2020, 9, 2632. [Google Scholar] [CrossRef]
- Gabryelska, A.; Stawski, R.; Sochal, M.; Szmyd, B.; Białasiewicz, P. Influence of one-night CPAP therapy on the changes of HIF-1α protein in OSA patients: A pilot study. J. Sleep Res. 2020, 29, e12995. [Google Scholar] [CrossRef]
- Tang, X.; Guo, D.; Lin, C.; Shi, Z.; Qian, R.; Fu, W.; Liu, J.; Li, X.; Fan, L. Upregulation of the gene expression of CLOCK is correlated with hypoxia-inducible factor 1α in advanced varicose lesions. Mol. Med. Rep. 2015, 12, 6164–6170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabryelska, A.; Karuga, F.; Szmyd, B.; Białasiewicz, P. HIF-1α as a Mediator of Insulin Resistance, T2DM, and Its Complications: Potential Links with Obstructive Sleep Apnea. Front. Physiol. 2020, 11, 1035. [Google Scholar] [CrossRef]
- Ghorbel, M.; Coulson, J.; Murphy, D. Crosstalk between hypoxic and circadian pathways: Cooperative roles for hypoxia-inducible factor 1α and CLOCK in transcriptional activation of the vasopressin gene. Mol. Cell Neurosci. 2003, 22, 396–404. [Google Scholar] [CrossRef]
- Yuan, P.; Yang, T.; Mu, J.; Zhao, J.; Yang, Y.; Yan, Z.; Hou, Y.; Chen, C.; Xing, J.; Zhang, H.; et al. Circadian clock gene NPAS2 promotes reprogramming of glucose metabolism in hepatocellular carcinoma cells. Cancer Lett. 2020, 469, 498–509. [Google Scholar] [CrossRef]
- Hogenesch, J.; Gu, Y.; Jain, S.; Bradfield, C. The basic-helix-loop-helix-PAS orphan MOP3 forms transcriptionally active complexes with circadian and hypoxia factors. Proc. Natl. Acad. Sci. USA 1998, 95, 5474–5479. [Google Scholar] [CrossRef] [Green Version]
- Peek, C.B.; Levine, D.C.; Cedernaes, J.; Taguchi, A.; Kobayashi, Y.; Tsai, S.J.; Bonar, N.A.; McNulty, M.R.; Ramsey, K.M.; Bass, J. Circadian Clock Interaction with HIF1α Mediates Oxygenic Metabolism and Anaerobic Glycolysis in Skeletal Muscle. Cell Metab. 2017, 25, 86–92. [Google Scholar] [CrossRef] [Green Version]
- Peek, C.B. Metabolic Implications of Circadian-HIF Crosstalk. Trends Endocrinol. Metab. 2020, 31, 459–468. [Google Scholar] [CrossRef]
- Peek, C.; Affinati, A.; Ramsey, K.; Kuo, H.; Yu, W.; Sena, L.; Ilkayeva, O.; Marcheva, B.; Kobayashi, Y.; Omura, C.; et al. Circadian Clock NAD + Cycle Drives Mitochondrial Oxidative Metabolism in Mice. Science 2013, 342, 1243417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakabayashi, H.; Ohta, Y.; Yamamoto, M.; Susuki, Y.; Taguchi, A.; Tanabe, K.; Kondo, M.; Hatanaka, M.; Nagao, Y.; Tanizawa, Y. Clock-controlled output gene Dbp is a regulator of Arnt/Hif-1β gene expression in pancreatic islet β-cells. Biochem. Biophys. Res. Commun. 2013, 434, 370–375. [Google Scholar] [CrossRef]
- Walton, Z.; Patel, C.; Brooks, R.; Yu, Y.; Ibrahim-Hashim, A.; Riddle, M.; Porcu, A.; Jiang, T.; Ecker, B.L.; Tameire, F.; et al. Acid Suspends the Circadian Clock in Hypoxia through Inhibition of mTOR. Cell 2018, 174, 72–87. [Google Scholar] [CrossRef] [Green Version]
- Baron, K.; Reid, K. Circadian misalignment, and health. Int. Rev. Psychiatry 2014, 26, 139–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcheva, B.; Ramsey, K.; Peek, C.; Affinati, A.; Maury, E.; Bass, J. Circadian Clocks and Metabolism. Handb. Exp. Pharmacol. 2013, 217, 127–155. [Google Scholar]
- Gabryelska, A.; Szmyd, B.; Panek, M.; Szemraj, J.; Kuna, P.; Białasiewicz, P. Serum Hypoxia-Inducible Factor-1α protein level as a diagnostic marker of obstructive sleep apnea. Pol. Arch. Intern. Med. 2019, 130, 158–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabryelska, A.; Szmyd, B.; Szemraj, J.; Stawski, R.; Sochal, M.; Białasiewicz, P. Patients with obstructive sleep apnea present with chronic upregulation of serum HIF-1α protein. J. Clin. Sleep Med. 2020, 16, 1761–1768. [Google Scholar] [CrossRef]
- Xie, Y.; Tang, Q.; Chen, G.; Xie, M.; Yu, S.; Zhao, J.; Chen, L. New Insights into the Circadian Rhythm, and Its Related Diseases. Front. Physiol. 2019, 10, 682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pace, A.; Iannella, G.; Rossetti, V.; Visconti, I.C.; Gulotta, G.; Cavaliere, C.; De Vito, A.; Maniaci, A.; Cocuzza, S.; Magliulo, G.; et al. Diagnosis of Obstructive Sleep Apnea in Patients with Allergic and Non-Allergic Rhinitis. Medicina 2020, 56, 454. [Google Scholar] [CrossRef]
- Noda, A.; Yasuma, F.; Okada, T.; Yokota, M. Influence of movement arousal on circardian rhythm of blood pressure in obstructive sleep apnea syndrome. J. Hypertens. 2000, 18, 539–544. [Google Scholar] [CrossRef]
- Dallmann, R.; Viola, A.; Tarokh, L.; Cajochen, C.; Brown, S. The human circadian metabolome. Proc. Natl. Acad. Sci. USA 2012, 109, 2625–2629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mason, I.; Qian, J.; Adler, G.; Scheer, F. Impact of circadian disruption on glucose metabolism: Implications for type 2 diabetes. Diabetologia 2020, 63, 462–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimba, S.; Ogawa, T.; Hitosugi, S.; Ichihashi, Y.; Nakadaira, Y.; Kobayashi, M.; Tezuka, M.; Kosuge, Y.; Ishige, K.; Ito, Y.; et al. Deficient of a Clock Gene, Brain and Muscle Arnt-Like Protein-1 (BMAL1), Induces Dyslipidemia and Ectopic Fat Formation. PLoS ONE 2011, 6, e25231. [Google Scholar] [CrossRef] [Green Version]
- Serin, Y.; Acar Tek, N. Effect of Circadian Rhythm on Metabolic Processes and the Regulation of Energy Balance. Ann. Nutr. Metab. 2019, 74, 322–330. [Google Scholar] [CrossRef]
- Gabryelska, A.; Chrzanowski, J.; Sochal, M.; Kaczmarski, P.; Turkiewicz, S.; Ditmer, M.; Karuga, F.F.; Czupryniak, L.; Białasiewicz, P. Nocturnal Oxygen Saturation Parameters as Independent Risk Factors for Type 2 Diabetes Mellitus among Obstructive Sleep Apnea Patients. J. Clin. Med. 2021, 10, 3770. [Google Scholar] [CrossRef]
- Koopman, A.D.M.; Rauh, S.P.; van Riet, E.; Groeneveld, L.; van der Heijden, A.A.; Elders, P.J.; Dekker, J.M.; Nijpels, G.; Beulens, J.W.; Rutters, F. The Association between Social Jetlag, the Metabolic Syndrome, and Type 2 Diabetes Mellitus in the General Population: The New Hoorn Study. J. Biol. Rhythm. 2017, 32, 359–368. [Google Scholar] [CrossRef]
- Onaolapo, A.; Onaolapo, O. Circadian dysrhythmia-linked diabetes mellitus: Examining melatonin’s roles in prophylaxis and management. World J. Diabetes 2018, 9, 99–114. [Google Scholar] [CrossRef]
- Scheer, F.; Hilton, M.; Mantzoros, C.; Shea, S. Adverse metabolic and cardiovascular consequences of circadian misalignment. Proc. Natl. Acad. Sci. USA 2009, 106, 4453–4458. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Abellán, P.; Madrid, J.; Ordovás, J.; Garaulet, M. Aspectos cronobiológicos de la obesidad y el síndrome metabólico. Endocrinol. Nutr. 2012, 59, 50–61. [Google Scholar] [CrossRef]
- Maury, E.; Ramsey, K.; Bass, J. Circadian Rhythms and Metabolic Syndrome. Circ. Res. 2010, 106, 447–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaines, J.; Vgontzas, A.; Fernandez-Mendoza, J.; Bixler, E. Obstructive sleep apnea and the metabolic syndrome: The road to clinically meaningful phenotyping, improved prognosis, and personalized treatment. Sleep Med. Rev. 2018, 42, 211–219. [Google Scholar] [CrossRef]
- Mahmood, K.; Akhter, N.; Eldeirawi, K.; Onal, E.; Christman, J.W.; Carley, D.W.; Herdegen, J.J. Prevalence of type 2 diabetes in patients with obstructive sleep apnea in a multi-ethnic sample. J. Clin. Sleep Med. 2009, 5, 215–221. [Google Scholar] [CrossRef] [Green Version]
- Nagao, A.; Kobayashi, M.; Koyasu, S.; Chow, C.; Harada, H. HIF-1-Dependent Reprogramming of Glucose Metabolic Pathway of Cancer Cells and Its Therapeutic Significance. Int. J. Mol. Sci. 2019, 20, 238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tokuda, F.; Sando, Y.; Matsui, H.; Koike, H.; Yokoyama, T. Serum levels of adipocytokines, adiponectin and leptin, in patients with obstructive sleep apnea syndrome. Intern. Med. 2008, 47, 1843–1849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chopra, S.; Rathore, A.; Younas, H.; Pham, L.V.; Gu, C.; Beselman, A.; Kim, I.Y.; Wolfe, R.R.; Perin, J.; Polotsky, V.Y.; et al. Obstructive Sleep Apnea Dynamically Increases Nocturnal Plasma Free Fatty Acids, Glucose, and Cortisol During Sleep. J. Clin. Endocrinol. Metab. 2017, 102, 3172–3181. [Google Scholar] [CrossRef]
- D’Souza, H.; Kapoor, K.G. Retinal vascular manifestations of obstructive sleep apnea. Curr. Opin. Ophthalmol. 2020, 31, 508–513. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Lv, F.L.; Wang, G.H. Effects of HIF-1α on diabetic retinopathy angiogenesis and VEGF expression. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 5071–5076. [Google Scholar]
- Dobrosielski, D.A.; Papandreou, C.; Patil, S.P.; Salas-Salvadó, J. Diet and exercise in the management of obstructive sleep apnoea and cardiovascular disease risk. Eur. Respir. Rev. 2017, 26, 160110. [Google Scholar] [CrossRef]
- Kuniyoshi, F.H.; Garcia-Touchard, A.; Gami, A.S.; Romero-Corral, A.; van der Walt, C.; Pusalavidyasagar, S.; Kara, T.; Caples, S.M.; Pressman, G.S.; Vasquez, E.C.; et al. Day-night variation of acute myocardial infarction in obstructive sleep apnea. J. Am. Coll. Cardiol. 2008, 52, 343–346. [Google Scholar] [CrossRef] [Green Version]
- Gami, A.S.; Howard, D.E.; Olson, E.J.; Somers, V.K. Day-night pattern of sudden death in obstructive sleep apnea. N. Engl. J. Med. 2005, 352, 1206–1214. [Google Scholar] [CrossRef]
- Zeidan-Shwiri, T.; Aronson, D.; Atalla, K.; Blich, M.; Suleiman, M.; Marai, I.; Gepstein, L.; Lavie, L.; Lavie, P.; Boulos, M. Circadian pattern of life-threatening ventricular arrhythmia in patients with sleep-disordered breathing and implantable cardioverter-defibrillators. Heart Rhythm 2011, 8, 657–662. [Google Scholar] [CrossRef]
- Gottlieb, E.; Landau, E.; Baxter, H.; Werden, E.; Howard, M.; Brodtmann, A. The bidirectional impact of sleep and circadian rhythm dysfunction in human ischaemic stroke: A systematic review. Sleep Med. Rev. 2019, 45, 54–69. [Google Scholar] [CrossRef] [PubMed]
- Black, N.; D’Souza, A.; Wang, Y.; Piggins, H.; Dobrzynski, H.; Morris, G.; Boyett, M.R. Circadian rhythm of cardiac electrophysiology, arrhythmogenesis, and the underlying mechanisms. Heart Rhythm. 2019, 16, 298–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Portaluppi, F.; Hermida, R. Circadian rhythms in cardiac arrhythmias and opportunities for their chronotherapy. Adv. Drug Deliv. Rev. 2007, 59, 940–951. [Google Scholar] [CrossRef]
- Karaaggac, K.; Vatansever, F.; Tenekecioglu, E.; Arican Ozluk, O.; Kuzeytemiz, M.; Topal, D.; Yilmaz, M. The Relationship between Non-Dipper Blood Pressure and Thoracic Aortic Diameter in Metabolic Syndrome. Eurasian J. Med. 2014, 46, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, T.; Onishi, K.; Dohi, K.; Okinaka, T.; Ito, M.; Isaka, N.; Nakano, T. Circadian rhythm of blood pressure is transformed from a dipper to a non-dipper pattern in shift workers with hypertension. J. Hum. Hypertens. 2002, 16, 193–197. [Google Scholar] [CrossRef] [Green Version]
- Bankir, L.; Bochud, M.; Maillard, M.; Bovet, P.; Gabriel, A.; Burnier, M. Nighttime Blood Pressure and Nocturnal Dipping Are Associated with Daytime Urinary Sodium Excretion in African Subjects. Hypertension 2008, 51, 891–898. [Google Scholar] [CrossRef] [Green Version]
- Williams, D.; Croal, B.; Furnace, J.; Ross, S.; Witte, K.; Webster, M.; Critchen, W.; Webster, J. The prevalence of a raised aldosterone—Renin ratio (ARR) among new referrals to a hypertension clinic. Blood Press. 2006, 15, 164–168. [Google Scholar] [CrossRef] [PubMed]
- Goto, N.; Uchida, K.; Morozumi, K.; Ueki, T.; Matsuoka, S.; Katayama, A.; Haba, T.; Tominaga, Y.; Fukuda, M.; Nakao, A.; et al. Circadian Blood Pressure Rhythm Is Disturbed by Nephrectomy. Hypertens. Res. 2005, 28, 301–306. [Google Scholar] [CrossRef] [Green Version]
- Marques, F.; Campain, A.; Tomaszewski, M.; Zukowska-Szczechowska, E.; Yang, Y.H.J.; Charchar, F.J.; Morris, B.J. Gene Expression Profiling Reveals Renin mRNA Overexpression in Human Hypertensive Kidneys and a Role for MicroRNAs. Hypertension 2011, 58, 1093–1098. [Google Scholar] [CrossRef] [Green Version]
- Morris, C.; Purvis, T.; Hu, K.; Scheer, F. Circadian misalignment increases cardiovascular disease risk factors in humans. Proc. Natl. Acad. Sci. USA 2016, 113, E1402–E1411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, T.; Zhao, W.; Chen, Y.; Ahokas, R.; Sun, Y. Vascular endothelial growth factor (VEGF)-A: Role on cardiac angiogenesis following myocardial infarction. Microvasc. Res. 2010, 80, 188–194. [Google Scholar] [CrossRef] [Green Version]
- Gabryelska, A.; Łukasik, Z.M.; Makowska, J.S.; Białasiewicz, P. Obstructive Sleep Apnea: From Intermittent Hypoxia to Cardiovascular Complications via Blood Platelets. Front. Neurol. 2018, 9, 635. [Google Scholar] [CrossRef]
- Koyanagi, S.; Kuramoto, Y.; Nakagawa, H.; Aramaki, H.; Ohdo, S.; Soeda, S.; Shimeno, H. A molecular mechanism regulating circadian expression of vascular endothelial growth factor in tumor cells. Cancer Res. 2003, 63, 7277–7283. [Google Scholar] [PubMed]
- Virag, J.; Lust, R. Circadian influences on myocardial infarction. Front. Physiol. 2014, 5, 422. [Google Scholar] [CrossRef]
- Hermida, R.; Smolensky, M. Chronotherapy of hypertension. Curr. Opin. Nephrol. Hypertens. 2004, 13, 501–505. [Google Scholar] [CrossRef]
- Karkoulias, K.; Lykouras, D.; Sampsonas, F.; Drakatos, P.; Canova, S.; Tsoukalas, G.; Spiropoulos, K. The Role of Endothelin-1 in Obstructive Sleep Apnea Syndrome and Pulmonary Arterial Hypertension: Pathogenesis and Endothelin-1 Antagonists. Curr. Med. Chem. 2010, 17, 1059–1066. [Google Scholar] [CrossRef]
- Liu, M.; Galli, G.; Wang, Y.; Fan, Q.; Wang, Z.; Wang, X.; Xiao, W. Novel Therapeutic Targets for Hypoxia-Related Cardiovascular Diseases: The Role of HIF-1. Front. Physiol. 2020, 11, 774. [Google Scholar] [CrossRef]
- Yacoub, M.; Youssef, I.; Salifu, M.; McFarlane, S. Cardiovascular Disease Risk in Obstructive Sleep apnea: An Update. J. Sleep Disord. Ther. 2018, 7, 155. [Google Scholar] [CrossRef]
- Gabryelska, A.; Białasiewicz, P. Hitting two birds with one stone: The potential role of serum hypoxia-inducible factor-1α protein levels in obstructive sleep apnea—Related cardiovascular disease. Authors’ reply. Pol. Arch. Intern. Med. 2020, 130, 162. [Google Scholar] [CrossRef]
- Walker, W.; Walton, J.; DeVries, A.; Nelson, R. Circadian rhythm disruption and mental health. Transl. Psychiatry 2020, 10, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Hood, S.; Amir, S. Neurodegeneration, and the Circadian Clock. Front. Aging Neurosci. 2017, 9, 170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennedy, S. Core symptoms of major depressive disorder: Relevance to diagnosis and treatment. Dialogues Clin. Neurosci. 2008, 10, 271–277. [Google Scholar]
- Lee, A.; Myung, S.; Cho, J.; Jung, Y.; Yoon, J.; Kim, M. Night Shift Work and Risk of Depression: Meta-analysis of Observational Studies. J. Korean Med. Sci. 2017, 32, 1091. [Google Scholar] [CrossRef]
- Oldham, M.A.; Ciraulo, D.A. Bright light therapy for depression: A review of its effects on chronobiology and the autonomic nervous system. Chronobiol. Int. 2014, 31, 305–319. [Google Scholar] [CrossRef] [PubMed]
- Maes, M.; Galecki, P.; Chang, Y.S.; Berk, M. A review on the oxidative and nitrosative stress (O&NS) pathways in major depression and their possible contribution to the (neuro)degenerative processes in that illness. Prog. Neuropsychopharmacol. Biol. Psychiatry 2011, 35, 676–692. [Google Scholar]
- Shibata, T.; Yamagata, H.; Uchida, S.; Otsuki, K.; Hobara, T.; Higuchi, F.; Abe, N.; Watanabe, Y. The alteration of hypoxia inducible factor-1 (HIF-1) and its target genes in mood disorder patients. Prog. Neuropsychopharmacol. Biol. Psychiatry 2013, 43, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Lacher, S.E.; Levings, D.C.; Freeman, S.; Slattery, M. Identification of a functional antioxidant response element at the HIF1A locus. Redox Biol. 2018, 19, 401–411. [Google Scholar] [CrossRef]
- Li, H.S.; Zhou, Y.N.; Li, L.; Li, S.F.; Long, D.; Chen, X.L.; Zhang, J.B.; Feng, L.; Li, Y.P. HIF-1α protects against oxidative stress by directly targeting mitochondria. Redox Biol. 2019, 25, 101109. [Google Scholar] [CrossRef]
- Turkiewicz, S.; Ditmer, M.; Sochal, M.; Białasiewicz, P.; Strzelecki, D.; Gabryelska, A. Obstructive Sleep Apnea as an Acceleration Trigger of Cellular Senescence Processes through Telomere Shortening. Int. J. Mol. Sci. 2021, 22, 12536. [Google Scholar] [CrossRef]
- Kang, I.; Kondo, D.; Kim, J.; Lyoo, I.K.; Yurgelun-Todd, D.; Hwang, J.; Renshaw, P.F. Elevating the level of hypoxia inducible factor may be a new potential target for the treatment of depression. Med. Hypotheses 2021, 146, 110398. [Google Scholar] [CrossRef] [PubMed]
- Liu, I.; Varinthra, P. Molecular basis for the association between depression and circadian rhythm. Tzu Chi Med. J. 2019, 31, 67. [Google Scholar] [CrossRef] [PubMed]
- Jehan, S.; Auguste, E.; Pandi-Perumal, S.R.; Kalinowski, J.; Myers, A.K.; Zizi, F.; Rajanna, M.G.; Jean-Louis, G.; McFarlane, S.I. Depression, Obstructive Sleep Apnea and Psychosocial Health. Sleep Med. Disord. Int. J. 2017, 1, 00012. [Google Scholar]
- Melo, M.; Abreu, R.; Linhares Neto, V.; de Bruin, P.; de Bruin, V. Chronotype and circadian rhythm in bipolar disorder: A systematic review. Sleep Med. Rev. 2017, 34, 46–58. [Google Scholar] [CrossRef] [Green Version]
- Kelly, T.; Douglas, L.; Denmark, L.; Brasuell, G.; Lieberman, D. The high prevalence of obstructive sleep apnea among patients with bipolar disorders. J. Affect. Disord. 2013, 151, 54–58. [Google Scholar] [CrossRef]
- Joo, E.Y.; Tae, W.S.; Lee, M.J.; Kang, J.W.; Park, H.S.; Lee, J.Y.; Suh, M.; Hong, S.B. Reduced Brain Gray Matter Concentration in Patients with Obstructive Sleep Apnea Syndrome. Sleep 2010, 33, 235–241. [Google Scholar] [CrossRef] [Green Version]
- Di Mauro, P.; Cocuzza, S.; Maniaci, A.; Ferlito, S.; Rasà, D.; Anzivino, R.; Vicini, C.; Iannella, G.; La Mantia, I. The Effect of Adenotonsillectomy on Children’s Behavior and Cognitive Performance with Obstructive Sleep Apnea Syndrome: State of the Art. Children 2021, 8, 921. [Google Scholar] [CrossRef]
- Ashton, A.; Jagannath, A. Disrupted Sleep and Circadian Rhythms in Schizophrenia and Their Interaction with Dopamine Signaling. Front. Neurosci. 2020, 14, 636. [Google Scholar] [CrossRef] [PubMed]
- Johansson, A.; Owe-Larsson, B.; Hetta, J.; Lundkvist, G. Altered circadian clock gene expression in patients with schizophrenia. Schizophr. Res. 2016, 174, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Li, S.; Chen, F.; Zhang, Y.; Li, P.; Jin, M.; Sun, Y.; Wang, F.; Mi, W.F.; Shi, L. Diurnal neurobiological alterations after exposure to clozapine in first-episode schizophrenia patients. Psychoneuroendocrinology 2016, 64, 108–116. [Google Scholar] [CrossRef]
- Wu, Y.Y.; Chang, E.T.; Yang, Y.C.; Chen, S.F.; Hsu, C.Y.; Shen, Y.C. Risk of obstructive sleep apnea in patients with schizophrenia: A nationwide population-based cohort study. Soc. Psychiatry Psychiatr. Epidemiol. 2020, 55, 1671–1677. [Google Scholar] [CrossRef] [PubMed]
- Bélanger, V.; Picard, N.; Cermakian, N. The Circadian Regulation of Presenilin-2Gene Expression. Chronobiol. Int. 2006, 23, 747–766. [Google Scholar] [CrossRef] [PubMed]
- Breen, D.P.; Vuono, R.; Nawarathna, U.; Fisher, K.; Shneerson, J.M.; Reddy, A.B.; Barker, R.A. Sleep, and circadian rhythm regulation in early Parkinson disease. JAMA Neurol. 2014, 71, 589–595. [Google Scholar] [CrossRef]
- Cai, Y.; Liu, S.; Sothern, R.B.; Xu, S.; Chan, P. Expression of clock genes Per1 and Bmal1 in total leukocytes in health and Parkinson’s disease. Eur. J. Neurol. 2010, 17, 550–554. [Google Scholar] [CrossRef]
- Chou, P.S.; Lai, C.L.; Chou, Y.H.; Chang, W. Sleep apnea and the subsequent risk of Parkinson’s disease: A 3-year nationwide population-based study. Neuropsychiatr. Dis. Treat. 2017, 13, 959–965. [Google Scholar] [CrossRef] [Green Version]
- Tsai, M.S.; Li, H.Y.; Huang, C.G.; Wang, R.Y.L.; Chuang, L.P.; Chen, N.H.; Liu, C.H.; Yang, Y.H.; Liu, C.Y.; Hsu, C.M.; et al. Risk of Alzheimer’s Disease in Obstructive Sleep Apnea Patients with or Without Treatment: Real-World Evidence. Laryngoscope 2020, 130, 2292–2298. [Google Scholar] [CrossRef]
- Kaminska, M.; Lafontaine, A.; Kimoff, R. The Interaction between Obstructive Sleep Apnea and Parkinsons Dis: Possible Mechanisms and Implications for Cognitive Function. Parkinson’s Dis. 2015, 2015, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Gaeta, A.; Benítez, I.; Jorge, C.; Torres, G.; Dakterzada, F.; Minguez, O.; Huerto, R.; Pujol, M.; Carnes, A.; Dalmases, M.; et al. Prevalence of obstructive sleep apnea in Alzheimer’s disease patients. J. Neurol. 2019, 267, 1012–1022. [Google Scholar] [CrossRef] [PubMed]
- Ancoli-Israel, S.; Palmer, B.W.; Cooke, J.R.; Corey-Bloom, J.; Fiorentino, L.; Natarajan, L.; Liu, L.; Ayalon, L.; He, F.; Loredo, J.S. Cognitive effects of treating obstructive sleep apnea in Alzheimer’s disease: A randomized controlled study. J. Am. Geriatr. Soc. 2008, 56, 2076–2081. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Name | Gene Location on Chromosome | Size (Da) | Size (Amino Acids) | Circadian Clock Function |
|---|---|---|---|---|
| PER1 | 17p13.1 | 136,212 | 1290 | repressor |
| PER2 | 2q37.3 | 136,579 | 1255 | repressor |
| PER3 | 1p36.23 | 131,888 | 1201 | repressor |
| Cry1 | 12q23.3 | 66,395 | 586 | repressor |
| Cry2 | 11p11.2 | 66,947 | 593 | repressor |
| BMAL1 (ARNTL) | 11p15.3 | 68,762 | 626 | activator |
| CLOCK | 4q12 | 95,304 | 846 | activator |
| RORα | 15q22.2 | 58,975 | 523 | regulator |
| REV-ERBα | 17q21.1 | 66,805 | 614 | regulator |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gabryelska, A.; Turkiewicz, S.; Karuga, F.F.; Sochal, M.; Strzelecki, D.; Białasiewicz, P. Disruption of Circadian Rhythm Genes in Obstructive Sleep Apnea Patients—Possible Mechanisms Involved and Clinical Implication. Int. J. Mol. Sci. 2022, 23, 709. https://doi.org/10.3390/ijms23020709
Gabryelska A, Turkiewicz S, Karuga FF, Sochal M, Strzelecki D, Białasiewicz P. Disruption of Circadian Rhythm Genes in Obstructive Sleep Apnea Patients—Possible Mechanisms Involved and Clinical Implication. International Journal of Molecular Sciences. 2022; 23(2):709. https://doi.org/10.3390/ijms23020709
Chicago/Turabian StyleGabryelska, Agata, Szymon Turkiewicz, Filip Franciszek Karuga, Marcin Sochal, Dominik Strzelecki, and Piotr Białasiewicz. 2022. "Disruption of Circadian Rhythm Genes in Obstructive Sleep Apnea Patients—Possible Mechanisms Involved and Clinical Implication" International Journal of Molecular Sciences 23, no. 2: 709. https://doi.org/10.3390/ijms23020709
APA StyleGabryelska, A., Turkiewicz, S., Karuga, F. F., Sochal, M., Strzelecki, D., & Białasiewicz, P. (2022). Disruption of Circadian Rhythm Genes in Obstructive Sleep Apnea Patients—Possible Mechanisms Involved and Clinical Implication. International Journal of Molecular Sciences, 23(2), 709. https://doi.org/10.3390/ijms23020709