
Two-Dimensional Interfacial Exchange Diffusion Has the Potential to Augment Spatiotemporal Precision of Ca2+ Signaling


{kind=link}
{kind=link}
Abstract
:1. Introduction
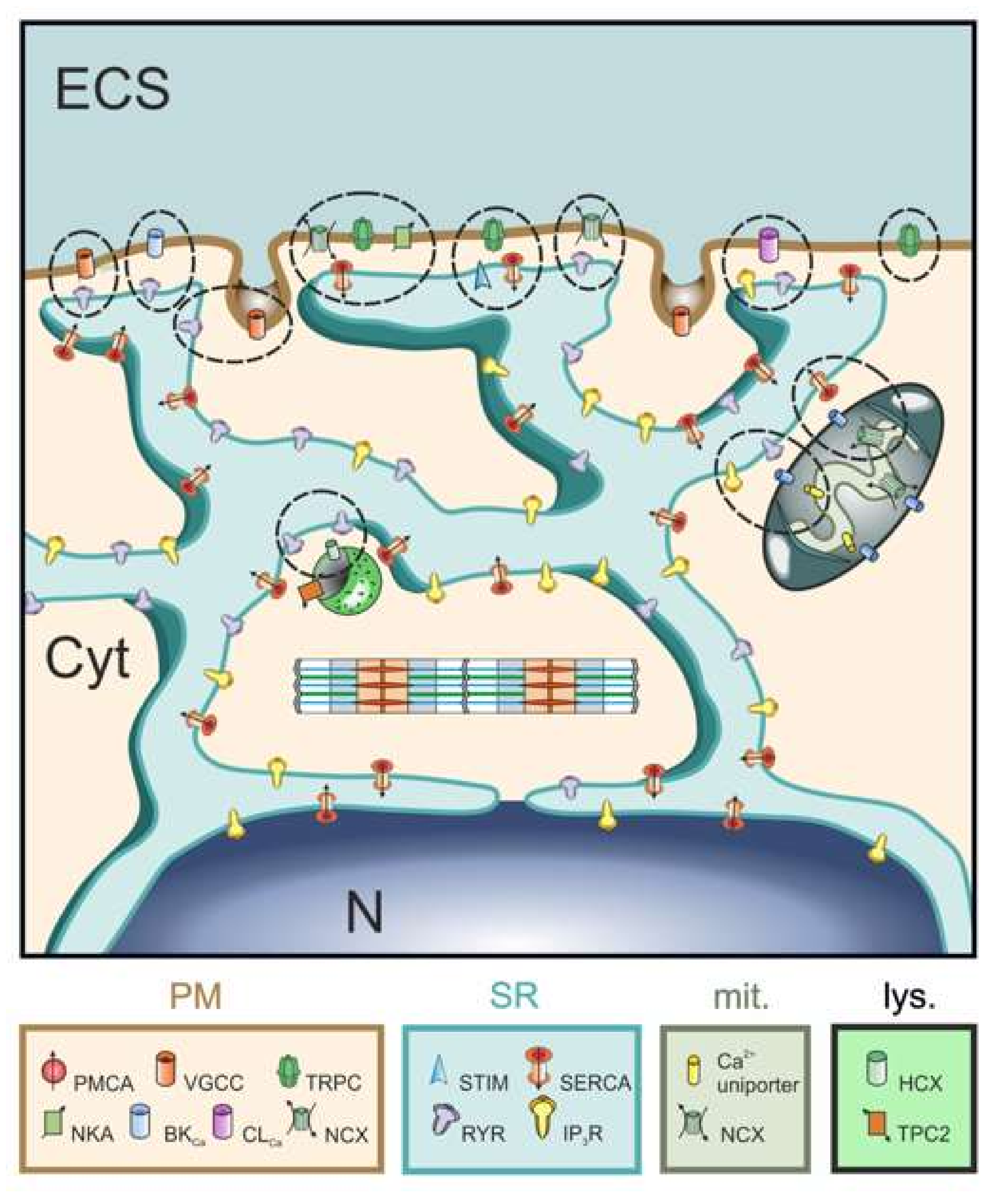
2. Background
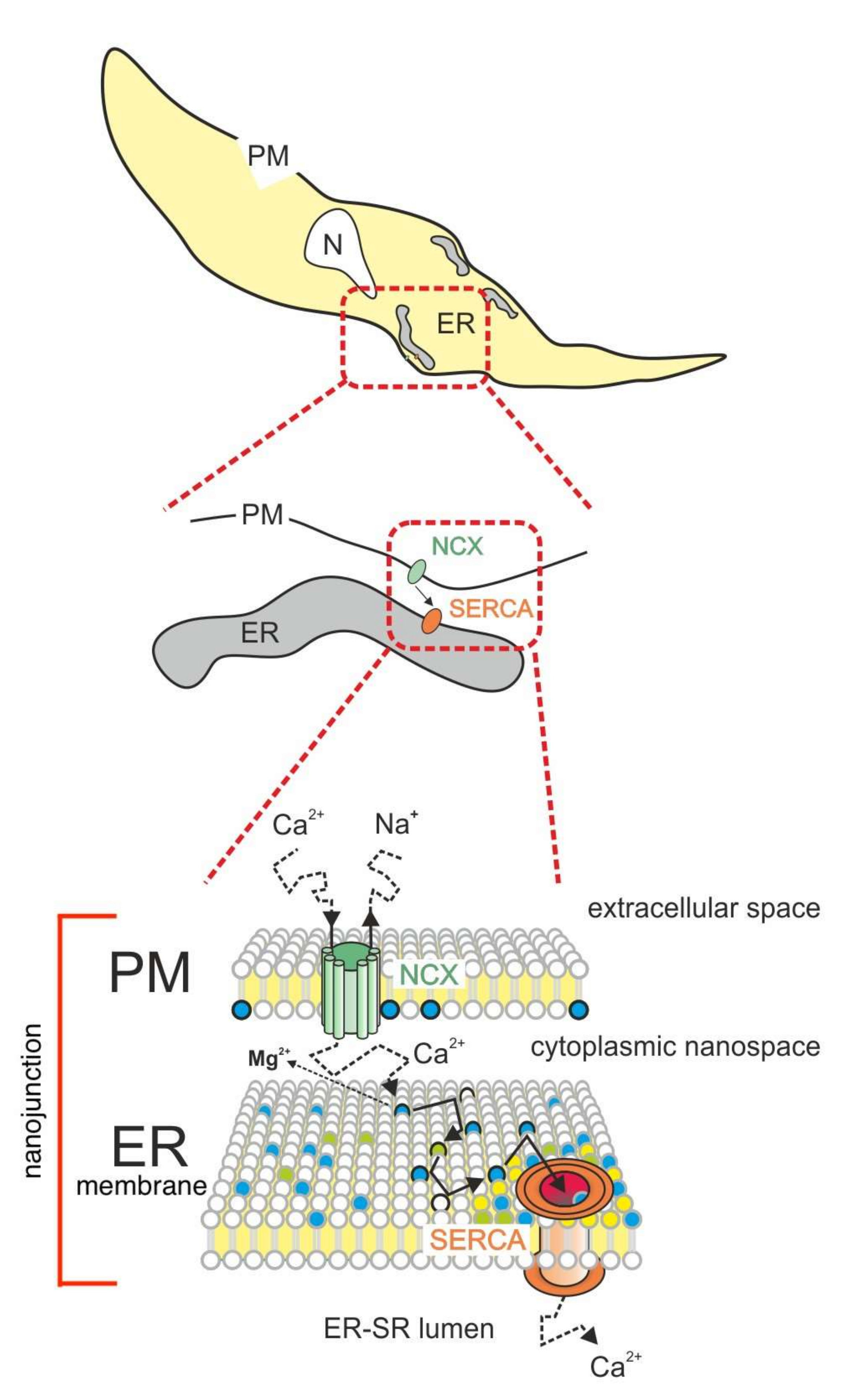
3. Model Description
4. Discussion
4.1. Rationale of the Proposed Model
4.2. Implications for Human Physiopathology
5. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ebashi, S.; Lipmann, F. Adenosine triphosphate-linked concentration of calcium ions in a par-ticulate fraction of rabbit muscle. J. Cell Biol. 1962, 14, 389–400. [Google Scholar] [CrossRef] [Green Version]
- Van Breemen, C. Blockade of Membrane Calcium Fluxes by Lanthanum in Relation to Vascular Smooth Muscle Contractility. Arch. Int. Physiol. Biochim. 1969, 77, 710–716. [Google Scholar] [CrossRef] [PubMed]
- Van Breemen, C.; Fameli, N.; Evans, A.M. Pan-junctional sarcoplasmic reticulum in vascular smooth muscle: Nanospace Ca2+ transport for site- and function-specific Ca2+ signaling. J. Physiol. 2013, 591, 2043–2054. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Qian, T.; He, R.; Wan, C.; Liu, Y.; Yu, H. Endoplasmic Reticulum–Plasma Membrane Contact Sites: Regulators, Mechanisms, and Physiological Functions. Front. Cell Dev. Biol. 2021, 9, 126. [Google Scholar] [CrossRef] [PubMed]
- Muallem, S.; Chung, W.Y.; Jha, A.; Ahuja, M. Lipids at membrane contact sites: Cell signaling and ion transport. EMBO Rep. 2017, 18, 1893–1904. [Google Scholar] [CrossRef]
- Lee, C.-H.; Kuo, K.-H.; Dai, J.; Leo, J.M.; Seow, C.Y.; van Breemen, C. Calyculin-A disrupts subplasmalemmal junction and recurring Ca2+ waves in vascular smooth muscle. Cell Calcium 2005, 37, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Fameli, N.; Van Breemen, C.; Kuo, K.-H. A quantitative model for linking Na+/Ca2+ exchanger to SERCA during refilling of the sarcoplasmic reticulum to sustain [Ca2+] oscillations in vascular smooth muscle. Cell Calcium 2007, 42, 565–575. [Google Scholar] [CrossRef] [Green Version]
- Pritchard, H.; Griffin, C.; Yamasaki, E.; Thakore, P.; Lane, C.; Greenstein, A.; Earley, S. Nanoscale coupling of junctophilin-2 and ryanodine receptors regulates vascular smooth muscle cell contractility. Proc. Natl. Acad. Sci. USA 2019, 116, 21874–21881. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.-H.; Rahimian, R.; Szado, T.; Sandhu, J.; Poburko, D.; Behra, T.; Chan, L.; Van Breemen, C. Sequential opening of IP3-sensitive Ca2+channels and SOC during α-adrenergic activation of rabbit vena cava. Am. J. Physiol. Circ. Physiol. 2002, 282, H1768–H1777. [Google Scholar] [CrossRef] [Green Version]
- Jay, E.E.; Mallinson, P.M.; Fong, S.K.; Metcalfe, B.L.; Grimes, R.W. Divalent cation diffusion in calcium fluorapatite. J. Mater. Sci. 2011, 46, 7459–7465. [Google Scholar] [CrossRef] [Green Version]
- Van Breemen, D.; Van Breemen, C. Calcium Exchange Diffusion in a Porous Phospholipid Ion-exchange Membrane. Nature 1969, 223, 898–900. [Google Scholar] [CrossRef] [PubMed]
- Tobias, J.M.; Agin, D.P.; Pawlowski, R. Phospholipid-Cholesterol Membrane Model. J. Gen. Physiol. 1962, 45, 989–1001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Breemen, C. Permselectivity of a porous phospholipid-cholesterol artificial membrane. Calcium and lanthanum effects. Biochem. Biophys. Res. Commun. 1968, 32, 977–983. [Google Scholar] [CrossRef]
- McLaughlin, S.; Brown, J. Diffusion of calcium ions in retinal rods. A theoretical calculation. J. Gen. Physiol. 1981, 77, 475–487. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Schwartz, D.K. Non-Brownian Interfacial Diffusion: Flying, Hopping, and Crawling. J. Phys. Chem. C 2020, 124. [Google Scholar] [CrossRef]
- Han, K.; Gericke, A.; Pastor, R.W. Characterization of Specific Ion Effects on PI(4,5)P2 Clustering: Molecular Dynamics Simulations and Graph-Theoretic Analysis. J. Phys. Chem. B 2020, 124, 1183–1196. [Google Scholar] [CrossRef]
- Bradley, R.P.; Slochower, D.R.; Janmey, P.A.; Radhakrishnan, R. Divalent cations bind to phosphoinositides to induce ion and isomer specific propensities for nano-cluster initiation in bilayer membranes. R. Soc. Open Sci. 2020, 7, 192208. [Google Scholar] [CrossRef]
- Melcrova, A.; Pokorna, S.; Pullanchery, S.; Kohagen, M.; Jurkiewicz, P.; Hof, M.; Jungwirth, P.; Cremer, P.S.; Cwiklik, L. The complex nature of calcium cation interactions with phospholipid bilayers. Sci. Rep. 2016, 6, 38035. [Google Scholar] [CrossRef]
- Wen, Y.; Vogt, V.M.; Feigenson, G.W. Multivalent Cation-Bridged PI(4,5)P2 Clusters Form at Very Low Concentrations. Biophys. J. 2018, 114, 2630–2639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujiwara, T.; Iwasawa, K.; Kalay, Z.; Tsunoyama, T.-A.; Watanabe, Y.; Umemura, Y.M.; Murakoshi, H.; Suzuki, K.G.N.; Nemoto, Y.L.; Morone, N.; et al. Confined diffusion of transmembrane proteins and lipids induced by the same actin meshwork lining the plasma membrane. Mol. Biol. Cell 2016, 27, 1101–1119. [Google Scholar] [CrossRef]
- Sarmento, M.J.; Coutinho, A.; Fedorov, A.; Prieto, M.; Fernandes, F. Ca2+ induces PI(4,5)P2 clusters on lipid bilayers at physiological PI(4,5)P2 and Ca2+ concentrations. Biochim. Biophys. Acta (BBA) Biomembr. 2014, 1838, 822–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drachmann, N.D.; Olesen, C.; Møller, J.V.; Guo, Z.; Nissen, P.; Bublitz, M. Comparing crystal structures of Ca2+-ATPase in the presence of different lipids. FEBS J. 2014, 281, 4249–4262. [Google Scholar] [CrossRef]
- Gao, Y.; Cao, E.; Julius, D.; Cheng, Y. TRPV1 structures in nanodiscs reveal mechanisms of ligand and lipid action. Nature 2016, 534, 347–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conrard, L.; Tyteca, D. Regulation of Membrane Calcium Transport Proteins by the Surrounding Lipid Environment. Biomolecules 2019, 9, 513. [Google Scholar] [CrossRef] [Green Version]
- Poveda, J.; Giudici, A.; Renart, M.; Molina, M.; Montoya, E.; Fernández-Carvajal, A.; Fernández-Ballester, G.; Encinar, J.; González-Ros, J. Lipid modulation of ion channels through specific binding sites. Biochim. Biophys. Acta (BBA) Biomembr. 2014, 1838, 1560–1567. [Google Scholar] [CrossRef] [Green Version]
- Lichtenegger, M.; Tiapko, O.; Svobodova, B.; Stockner, T.; Glasnov, T.; Schreibmayer, W.; Platzer, D.; De La Cruz, G.G.; Krenn, S.; Schober, R.; et al. An optically controlled probe identifies lipid-gating fenestrations within the TRPC3 channel. Nat. Chem. Biol. 2018, 14, 396–404. [Google Scholar] [CrossRef]
- Cai, R.; Liu, X.; Zhang, R.; Hofmann, L.; Zheng, W.; Amin, R.; Wang, L.; Hu, Q.; Peng, J.-B.; Michalak, M.; et al. Autoinhibition of TRPV6 Channel and Regulation by PIP2. Iscience 2020, 23, 101444. [Google Scholar] [CrossRef] [PubMed]
- Barth, D.; Lückhoff, A.; Kühn, F. Species-Specific Regulation of TRPM2 by PI(4,5)P2 via the Membrane Interfacial Cavity. Int. J. Mol. Sci. 2021, 22, 4637. [Google Scholar] [CrossRef]
- Espinoza-Fonseca, L.M. Probing the effects of nonannular lipid binding on the stability of the calcium pump SERCA. Sci. Rep. 2019, 9, 3349. [Google Scholar] [CrossRef] [Green Version]
- Toyoshima, C.; Nakasako, M.; Nomura, H.; Ogawa, H. Crystal structure of the calcium pump of sarcoplasmic reticulum at 2.6 Å resolution. Nature 2000, 405, 647–655. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Bi, Y.; Yang, W.; Guo, X.; Jiang, Y.; Wan, C.; Li, L.; Bai, Y.; Guo, J.; Wang, Y.; et al. Ca2+ regulates T-cell receptor activation by modulating the charge property of lipids. Nat. Cell Biol. 2012, 493, 111–115. [Google Scholar] [CrossRef]
- Yoshida, T.; Taga, K.; Okabayashi, H.; Kamaya, H.; Issaku, U. Changes in surface capacitance and conductance parallel to phospholipid membranes associated with phase transition: Effects of halothane. Biochim. Biophys. Acta (BBA) Biomembr. 1989, 984, 253–256. [Google Scholar] [CrossRef]
- Navarro-Dorado, J.; García-Alonso, M.; Van Breemen, C.; Tejerina, T.; Fameli, N. Calcium oscillations in human mesenteric vascular smooth muscle. Biochem. Biophys. Res. Commun. 2014, 445, 84–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, J.M.; Syyong, H.; Navarro-Dorado, J.; Redondo, S.; Alonso, M.; van Breemen, C.; Tejerina, T. A comparative study of α-adrenergic receptor mediated Ca2+ signals and contraction in intact human and mouse vascular smooth muscle. Eur. J. Pharmacol. 2010, 629, 82–88. [Google Scholar] [CrossRef]
- Piccialli, I.; Ciccone, R.; Secondo, A.; Boscia, F.; Tedeschi, V.; de Rosa, V.; Cepparulo, P.; Annunziato, L.; Pannaccione, A. The Na+/Ca2+ Exchanger 3 Is Functionally Coupled With the NaV1.6 Voltage-Gated Channel and Promotes an Endoplasmic Reticulum Ca2+ Refilling in a Transgenic Model of Alzheimer’s Disease. Front. Pharmacol. 2021, 12, 3582. [Google Scholar] [CrossRef]
- Franzini-Armstrong, C. The relationship between form and function throughout the history of excitation–contraction coupling. J. Gen. Physiol. 2018, 150, 189–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, O.H.; Courjaret, R.; Machaca, K. Ca2+ tunnelling through the ER lumen as a mechanism for delivering Ca2+entering via store-operated Ca2+channels to specific target sites. J. Physiol. 2017, 595, 2999–3014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, J.; Navarro-Dorado, J.; Clark, J.H.; Kinnear, N.P.; Meinke, P.; Schirmer, E.C.; Evans, A.M. The cell-wide web coordinates cellular processes by directing site-specific Ca2+ flux across cytoplasmic nanocourses. Nat. Commun. 2019, 10, 2299. [Google Scholar] [CrossRef] [Green Version]
- Iino, M.; Kasai, H.; Yamazawa, T. Visualization of neural control of intracellular Ca2+ concentration in single vascular smooth muscle cells in situ. EMBO J. 1994, 13, 5026–5031. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Breemen, C.; Fameli, N.; Groschner, K. Two-Dimensional Interfacial Exchange Diffusion Has the Potential to Augment Spatiotemporal Precision of Ca2+ Signaling. Int. J. Mol. Sci. 2022, 23, 850. https://doi.org/10.3390/ijms23020850
van Breemen C, Fameli N, Groschner K. Two-Dimensional Interfacial Exchange Diffusion Has the Potential to Augment Spatiotemporal Precision of Ca2+ Signaling. International Journal of Molecular Sciences. 2022; 23(2):850. https://doi.org/10.3390/ijms23020850
Chicago/Turabian Stylevan Breemen, Cornelis, Nicola Fameli, and Klaus Groschner. 2022. "Two-Dimensional Interfacial Exchange Diffusion Has the Potential to Augment Spatiotemporal Precision of Ca2+ Signaling" International Journal of Molecular Sciences 23, no. 2: 850. https://doi.org/10.3390/ijms23020850
APA Stylevan Breemen, C., Fameli, N., & Groschner, K. (2022). Two-Dimensional Interfacial Exchange Diffusion Has the Potential to Augment Spatiotemporal Precision of Ca2+ Signaling. International Journal of Molecular Sciences, 23(2), 850. https://doi.org/10.3390/ijms23020850