
Interactive Dynamics of Cell Volume and Cell Death in Human Erythrocytes Exposed to α-Hemolysin from Escherichia coli

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results
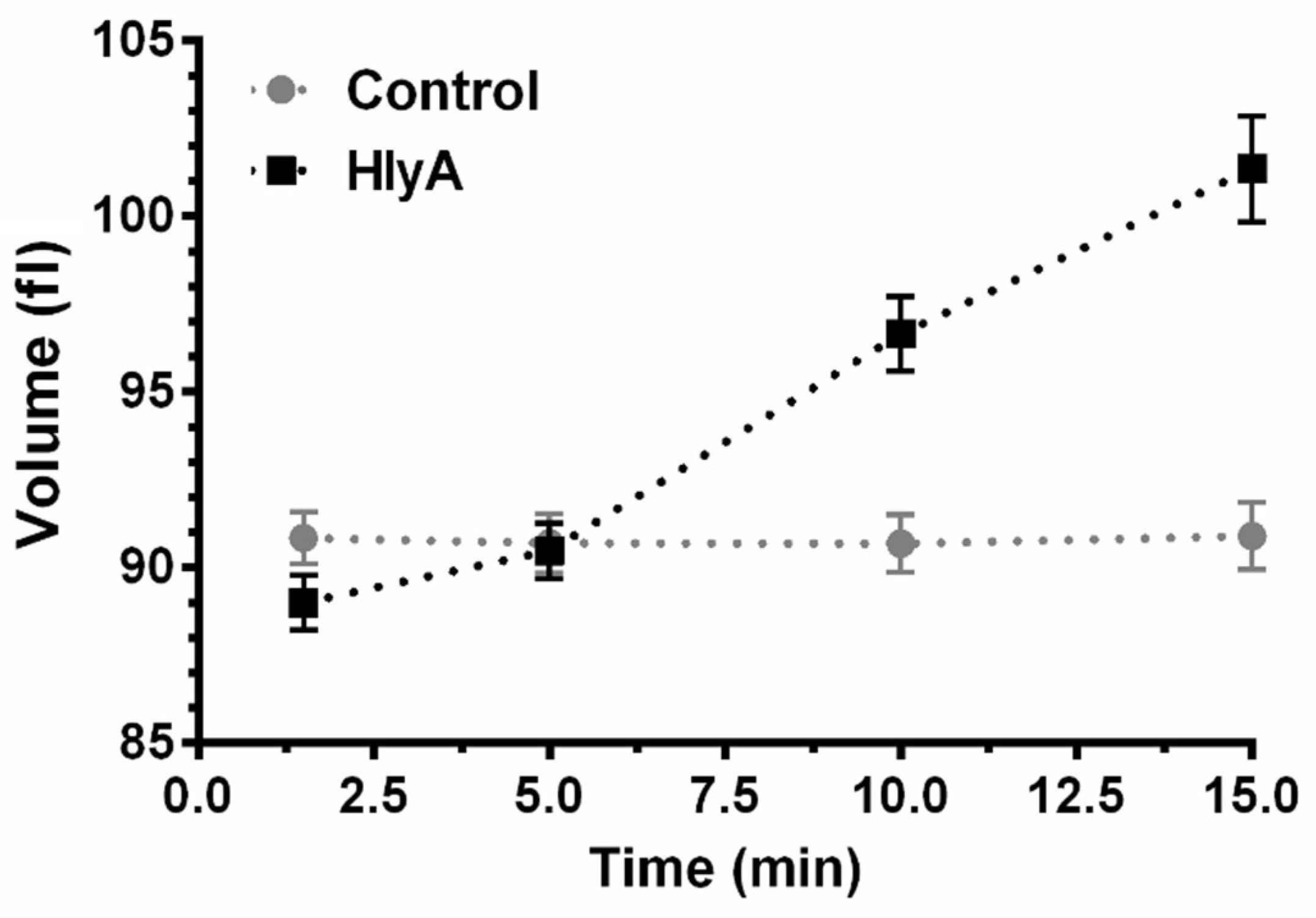
2.1. Pre-Lytic Volume Changes
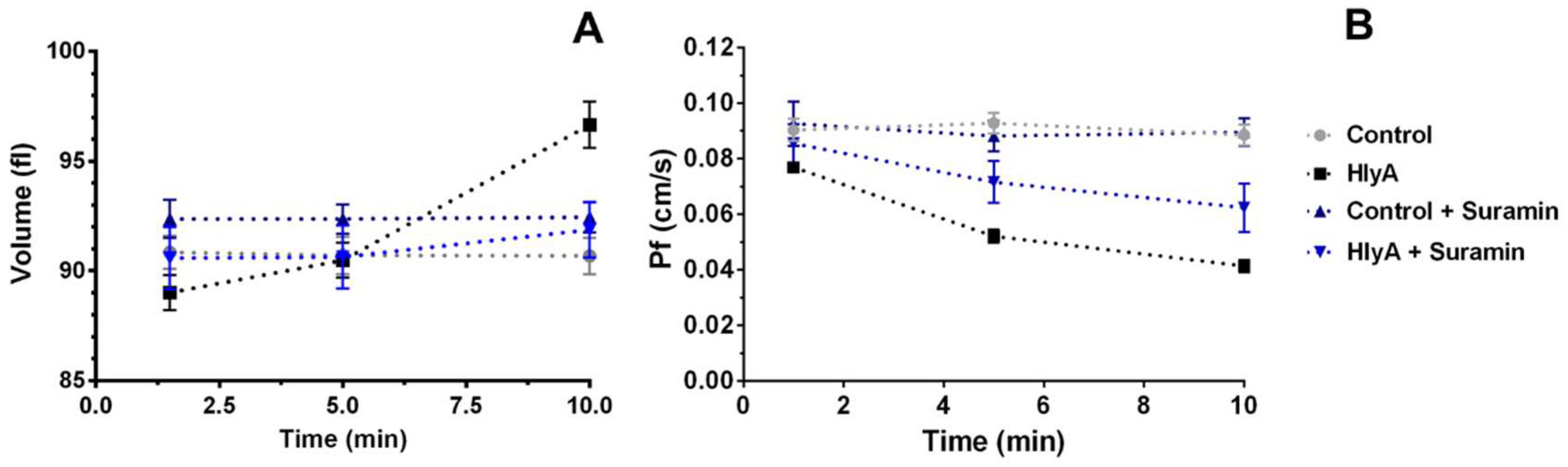
2.2. Pf in HlyA Treated RBCs
2.3. Effect of Purinergic Signalling on Pf
2.4. Effects of MCV on Pf
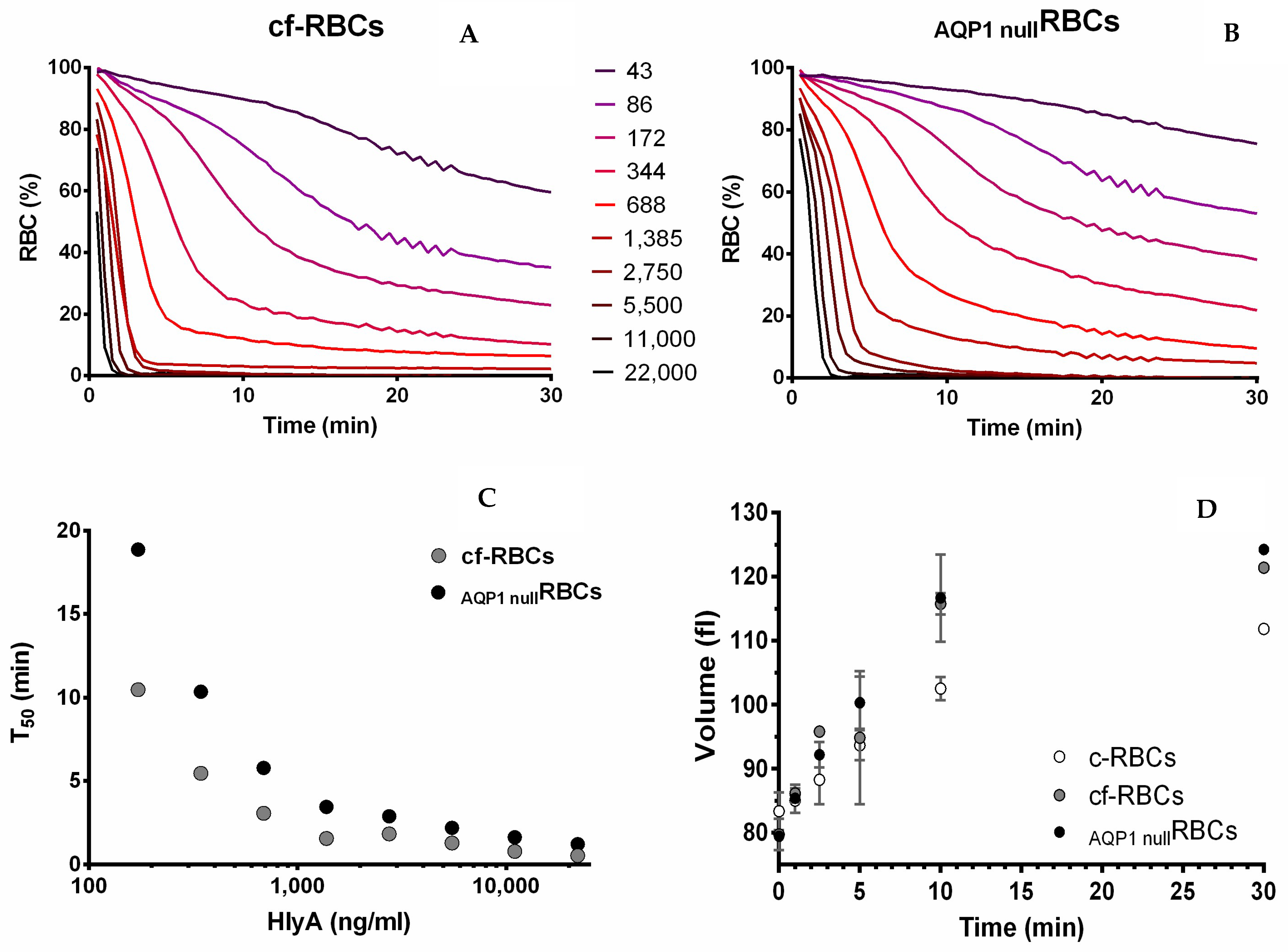
2.5. Role of AQP1 in HlyA Activity
Kinetics of Hemolysis, and Its Relationship to Changes in MCV and Pf
2.6. How Cell Volume Is Linked to Hemolysis
2.7. Mathematical Models Explaining HlyA-Dependent Kinetics of MCV and Hemolysis
- Model 1. Changes MCV and hemolysis on hypo-osmotic media
- Model 2. HlyA-dependent changes in MCV and hemolysis
3. Discussion
4. Methods
4.1. Media Used
- Isosmotic medium: (in mM) 137 NaCl, 2.7 KCl, 2.5 Na2HPO4, 1.50 KH2PO4, 1.32 CaCl2, 1.91 MgSO4, 5 glucose, pH 7.4 at 25 °C, 300 mosM.
- Hypo-osmotic medium: isosmotic medium reduced in NaCl content to obtain 55, 82, 110, 137, 164, 192, 219, 247 and 274 mosM.
- Tris-Chloride (TC) buffer: (in mM) 20 Tris, 150 NaCl, pH 7.4 at 25 °C.
4.2. Isolation of RBCs
- Fresh RBCs: RBCs were obtained from healthy individuals by venipuncture. After blood collection, the sample was centrifuged at 900× g for 3 min and the plasma, the platelets and the leucocytes were removed and discarded. Isolated RBCs were resuspended and washed three times in isosmotic medium. Finally, packed RBCs were resuspended at the corresponding hematocrit. The procedure was approved by the Ethics Committee on Clinical Investigation of the School of Pharmacy and Biochemistry, University of Buenos Aires (Res RESCD-2020-281-E-UBA-DCT_FFYB).
- Frozen RBCs: Control RBCs (cf-RBCs) and RBCs from an individual lacking the major water channel aquaporin 1 (AQP1nullRBCs) [27] were provided by the Centre National de Référence des Groupes Sanguins (Paris, France). Before the study, cryopreserved RBCs of both conditions were thawed and washed in isosmotic medium as described for fresh RBCs.
4.3. Toxin Purification
4.4. RBCs Treatment
4.5. Microscopy
4.5.1. Videomicroscopy
4.5.2. Scanning Electron Microscopy
4.6. Hemolysis Assays
4.6.1. Quantification of Hemoglobin Release at 405 nm
4.6.2. Measurement of Light Scattering at 595 nm
4.7. Stopped-Flow Assay
4.8. Pf Calculation
4.9. MCV Determinations
4.10. RBCs Forward Light Scattering (FSC)
4.11. Western Blot of AQP1
4.12. Model Fit
4.13. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | 40 ng/mL HlyA without Suramin | 40 ng/mL HlyA with Suramin |
|---|---|---|
| A (fmol/(cell*min)) | 0.92 ± 0.06 | 0.61 ± 0.05 |
| σΔosm (fmol/cell) | 5.0 ± 0.6 | 4.3 ± 0.7 |
| B (fmol/cell) | 4.3 ± 1.1 | |
| k1 (min−1) | <1.15 | |
References
- Gadeberg, O.V.; Orskov, I.; Rhodes, J.M. Cytotoxic Effect of an Alpha-Hemolytic Escherichia coli Strain on Human Blood Monocytes and Granulocytes in Vitro. Infect. Immun. 1983, 41, 358–364. [Google Scholar] [CrossRef] [Green Version]
- Keane, W.F.; Welch, R.; Gekker, G.; Peterson, P.K. Mechanism of Escherichia coli Alpha-Hemolysin-Induced Injury to Isolated Renal Tubular Cells. Am. J. Pathol. 1987, 126, 350–357. [Google Scholar]
- Wiles, T.J.; Mulvey, M.A. The RTX Pore-Forming Toxin α-Hemolysin of Uropathogenic Escherichia coli: Progress and Perspectives. Futur. Microbiol. 2013, 8, 73–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linhartová, I.; Bumba, L.; Mašín, J.; Basler, M.; Osička, R.; Kamanová, J.; Procházková, K.; Adkins, I.; Hejnová-Holubová, J.; Sadílková, L.; et al. RTX Proteins: A Highly Diverse Family Secreted by a Common Mechanism. FEMS Microbiol. Rev. 2010, 34, 1076–1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Hanley, P.; Low, D.; Romero, I.; Lark, D.; Vosti, K.; Falkow, S.; Schoolnik, G. Gal-Gal Binding and Hemolysin Phenotypes and Genotypes Associated with Uropathogenic Escherichia coli. New Engl. J. Med. 1985, 313, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Nagy, G.; Altenhoefer, A.; Knapp, O.; Maier, E.; Dobrindt, U.; Blum-Oehler, G.; Benz, R.; Emődy, L.; Hacker, J. Both α-Haemolysin Determinants Contribute to Full Virulence of Uropathogenic Escherichia coli Strain. Microbes Infect. 2006, 8, 2006–2012. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.R. Virulence Factors in Escherichia coli Urinary Tract Infection. Clin. Microbiol. Rev. 1991, 4, 80–128. [Google Scholar] [CrossRef] [PubMed]
- Ulleryd, P.; Lincoln, K.; Scheutz, F.; Sandberg, T. Virulence Characteristics of Escherichia coli in Relation to Host Response in Men with Symptomatic Urinary Tract Infection. Clin. Infect. Dis. 1994, 18, 579–584. [Google Scholar] [CrossRef] [PubMed]
- Marrs, C.F.; Zhang, L.; Foxman, B. Escherichia coli mediated Urinary Tract Infections: Are There Distinct Uropathogenic, E. coli (UPEC) Pathotypes? FEMS Microbiol. Lett. 2005, 252, 183–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnsen, N.; Hamilton, A.D.M.; Greve, A.S.; Christensen, M.G.; Therkildsen, J.R.; Wehmöller, J.; Skals, M.; Praetorius, H.A. α-Haemolysin Production, as a Single Factor, Causes Fulminant Sepsis in a Model of Escherichia coli-Induced Bacteraemia. Cell. Microbiol. 2019, 21, e13017. [Google Scholar] [CrossRef] [PubMed]
- Ristow, L.; Welch, R.A. Hemolysin of Uropathogenic Escherichia coli: A Cloak or a Dagger? Biochim. Biophys. Acta (BBA)-Biomembr. 2016, 1858, 538–545. [Google Scholar] [CrossRef] [PubMed]
- Herlax, V.; Henning, M.F.; Bernasconi, A.M.; Goni, F.M.; Bakas, L. The Lytic Mechanism of Escherichia coli α-Hemolysin Associated to Outer Membrane Vesicles. Health 2010, 2, 484–492. [Google Scholar] [CrossRef] [Green Version]
- Benz, R. Channel Formation by RTX-Toxins of Pathogenic Bacteria: Basis of Their Biological Activity. Biochim. Biophys. Acta (BBA)-Biomembr. 2016, 1858, 526–537. [Google Scholar] [CrossRef] [PubMed]
- Valeva, A.; Palmer, M.; Bhakdi, S. Staphylococcal α-Toxin: Formation of the Heptameric Pore Is Partially Cooperative and Proceeds through Multiple Intermediate Stages. Biochemistry 1997, 36, 13298–13304. [Google Scholar] [CrossRef] [PubMed]
- Gouaux, J.E.; Braha, O.; Hobaugh, M.R.; Song, L.; Cheley, S.; Shustak, C.; Bayley, H. Subunit Stoichiometry of Staphylococcal Alpha-Hemolysin in Crystals and on Membranes: A Heptameric Transmembrane Pore. Proc. Natl. Acad. Sci. USA 1994, 91, 12828–12831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moayeri, M.; Welch, R.A. Effects of Temperature, Time, and Toxin Concentration on Lesion Formation by the Escherichia coli Hemolysin. Infect. Immun. 1994, 62, 4124–4134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menestrina, G. Escherichia coli hemolysin Permeabilizes Small Unilamellar Vesicles Loaded with Calcein by a Single-Hit Mechanism. FEBS Lett. 1988, 232, 217–220. [Google Scholar] [CrossRef] [Green Version]
- Ludwig, A.; Benz, R.; Goebel, W. Oligomerization of Escherichia coli Haemolysin (HlyA) is Involved in Pore Formation. Mol. Genet. Genom. 1993, 241, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Herlax, V.; Maté, S.; Rimoldi, O.; Bakás, L. Relevance of Fatty Acid Covalently Bound to Escherichia coli α-Hemolysin and Membrane Microdomains in the Oligomerization Process. J. Biol. Chem. 2009, 284, 25199–25210. [Google Scholar] [CrossRef] [Green Version]
- Skals, M.; Jensen, U.B.; Ousingsawat, J.; Kunzelmann, K.; Leipziger, J.; Praetorius, H.A. Escherichia coli α-Hemolysin Triggers Shrinkage of Erythrocytes Via KCa3.1 and TMEM16A Channels with Subsequent Phosphatidylserine Exposure. J. Biol. Chem. 2010, 285, 15557–15565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, E.; Qadri, S.M.; Lang, F. Killing Me Softly–Suicidal Erythrocyte Death. Int. J. Biochem. Cell Biol. 2012, 44, 1236–1243. [Google Scholar] [CrossRef]
- Velásquez, F.C.; Maté, S.; Bakás, L.; Herlax, V. Induction of Eryptosis by Low Concentrations of E. coli Alpha-Hemolysin. Biochim. Biophys. Acta (BBA)-Biomembr. 2015, 1848, 2779–2788. [Google Scholar] [CrossRef] [Green Version]
- Denis, M.L.; Lefevre, S.; Alvarez, C.; Lauri, N.; Enrique, N.; Rinaldi, D.; Gonzalez-Lebrero, R.; Vecchio, L.; Espelt, M.; Stringa, P.; et al. Regulation of Extracellular ATP of Human Erythrocytes Treated with α-Hemolysin. Effects of Cell Volume, Morphology, Rheology and Hemolysis. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2019, 1866, 896–915. [Google Scholar] [CrossRef]
- Skals, M.; Jorgensen, N.R.; Leipziger, J.; Praetorius, H.A. α-Hemolysin from Escherichia coli Uses Endogenous Amplification through P2X Receptor Activation to Induce Hemolysis. Proc. Natl. Acad. Sci. USA 2009, 106, 4030–4035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Preston, G.M.; Carroll, T.P.; Guggino, W.B.; Agre, P. Appearance of Water Channels in Xenopus Oocytes Expressing Red Cell CHIP28 Protein. Science 1992, 256, 385–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roudier, N.; Verbavatz, J.-M.; Maurel, C.; Ripoche, P.; Tacnet, F. Evidence for the Presence of Aquaporin-3 in Human Red Blood Cells. J. Biol. Chem. 1998, 273, 8407–8412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azouzi, S.; Gueroult, M.; Ripoche, P.; Genetet, S.; Colin Aronovicz, Y.; Le Van Kim, C.; Etchebest, C.; Mouro-Chanteloup, I. Energetic and Molecular Water Permeation Mechanisms of the Human Red Blood Cell Urea Transporter, B. PLoS ONE 2013, 8, e82338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathai, J.C.; Mori, S.; Smith, B.L.; Preston, G.M.; Mohandas, N.; Collins, M.; van Zijl, P.C.M.; Zeidel, M.L.; Agre, P. Functional Analysis of Aquaporin-1 Deficient Red Cells. J. Biol. Chem. 1996, 271, 1309–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vazquez, R.F.; Maté, S.M.; Bakás, L.S.; Fernández, M.M.; Malchiodi, E.L.; Herlax, V.S. Novel Evidence for the Specific Interaction Between Cholesterol and α-Haemolysin of Escherichia coli. Biochem. J. 2014, 458, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Munksgaard, P.S.; Vorup-Jensen, T.; Reinholdt, J.; Söderström, C.M.; Poulsen, K.; Leipziger, J.; Praetorius, H.A.; Skals, M. Leukotoxin from Aggregatibacter actinomycetemcomitans causes Shrinkage and P2X Receptor-Dependent Lysis of Human Erythrocytes. Cell. Microbiol. 2012, 14, 1904–1920. [Google Scholar] [CrossRef] [PubMed]
- Denis, F.L.; Alvarez, H.A.; Lauri, N.; Alvarez, C.L.; Chara, O.; Schwarzbaum, P.J. Dynamic Regulation of Cell Volume and Extracellular ATP of Human Erythrocytes. PLoS ONE 2016, 11, e0158305. [Google Scholar] [CrossRef] [Green Version]
- Genetet, S.; Ripoche, P.; Picot, J.; Bigot, S.; Delaunay, J.; Armari-Alla, C.; Colin, Y.; Mouro-Chanteloup, I. Human RhAG Ammonia Channel is Impaired by the Phe65Ser Mutation in Overhydrated Stomatocytic Red Cells. Am. J. Physiol. Physiol. 2012, 302, C419–C428. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, R.F.; MatéS, M.; Bakás, L.S.; Muñoz-Garay, C.; Herlax, V.S. Relationship between Intracellular Calcium and Morphologic Changes in Rabbit Erythrocytes: Effects of the Acylated and Unacylated Forms of E. coli Alpha-Hemolysin. Biochim. Biophys. Acta (BBA)-Biomembr. 2016, 1858, 1944–1953. [Google Scholar] [CrossRef] [PubMed]
- Pafundo, D.; Alvarez, C.L.; Krumschnabel, G.; Schwarzbaum, P.J. A Volume Regulatory Response Can Be Triggered by Nucleosides in Human Erythrocytes, a Perfect Osmometer No Longer. J. Biol. Chem. 2010, 285, 6134–6144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, B.; Verkman, A.S. Analysis of Double Knockout Mice Lacking Aquaporin-1 and Urea Transporter UT-B. J. Biol. Chem. 2002, 277, 36782–36786. [Google Scholar] [CrossRef] [Green Version]
- King, L.S.; Choi, M.; Fernandez, P.C.; Cartron, J.-P.; Agre, P. Defective Urinary Concentrating Ability Due to a Complete Deficiency of Aquaporin-1. New Engl. J. Med. 2001, 345, 175–179. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Montalbetti, N.; Denis, M.F.L.; Pignataro, O.P.; Kobatake, E.; Lazarowski, E.R.; Schwarzbaum, P.J. Homeostasis of Extracellular ATP in Human Erythrocytes. J. Biol. Chem. 2011, 286, 38397–38407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Heeswijk, M.P.E.; van Os, C.H. Osmotic Water Permeabilities of Brush Border and Basolateral Membrane Vesicles from Rat Renal Cortex and Small Intestine. J. Membr. Biol. 1986, 92, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Freedman, J.C.; Hoffman, J.F. Ionic and Osmotic Equilibria of Human Red Blood Cells Treated with Nystatin. J. Gen. Physiol. 1979, 74, 157–185. [Google Scholar] [CrossRef] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saffioti, N.A.; Lauri, N.; Cané, L.; Gonzalez-Lebrero, R.; Alleva, K.; Mouro-Chanteloup, I.; Ostuni, M.A.; Herlax, V.; Schwarzbaum, P.J. Interactive Dynamics of Cell Volume and Cell Death in Human Erythrocytes Exposed to α-Hemolysin from Escherichia coli. Int. J. Mol. Sci. 2022, 23, 872. https://doi.org/10.3390/ijms23020872
Saffioti NA, Lauri N, Cané L, Gonzalez-Lebrero R, Alleva K, Mouro-Chanteloup I, Ostuni MA, Herlax V, Schwarzbaum PJ. Interactive Dynamics of Cell Volume and Cell Death in Human Erythrocytes Exposed to α-Hemolysin from Escherichia coli. International Journal of Molecular Sciences. 2022; 23(2):872. https://doi.org/10.3390/ijms23020872
Chicago/Turabian StyleSaffioti, Nicolas A., Natalia Lauri, Lucia Cané, Rodolfo Gonzalez-Lebrero, Karina Alleva, Isabelle Mouro-Chanteloup, Mariano A. Ostuni, Vanesa Herlax, and Pablo Julio Schwarzbaum. 2022. "Interactive Dynamics of Cell Volume and Cell Death in Human Erythrocytes Exposed to α-Hemolysin from Escherichia coli" International Journal of Molecular Sciences 23, no. 2: 872. https://doi.org/10.3390/ijms23020872
APA StyleSaffioti, N. A., Lauri, N., Cané, L., Gonzalez-Lebrero, R., Alleva, K., Mouro-Chanteloup, I., Ostuni, M. A., Herlax, V., & Schwarzbaum, P. J. (2022). Interactive Dynamics of Cell Volume and Cell Death in Human Erythrocytes Exposed to α-Hemolysin from Escherichia coli. International Journal of Molecular Sciences, 23(2), 872. https://doi.org/10.3390/ijms23020872