
Glial Modulation of Energy Balance: The Dorsal Vagal Complex Is No Exception

 ,
, 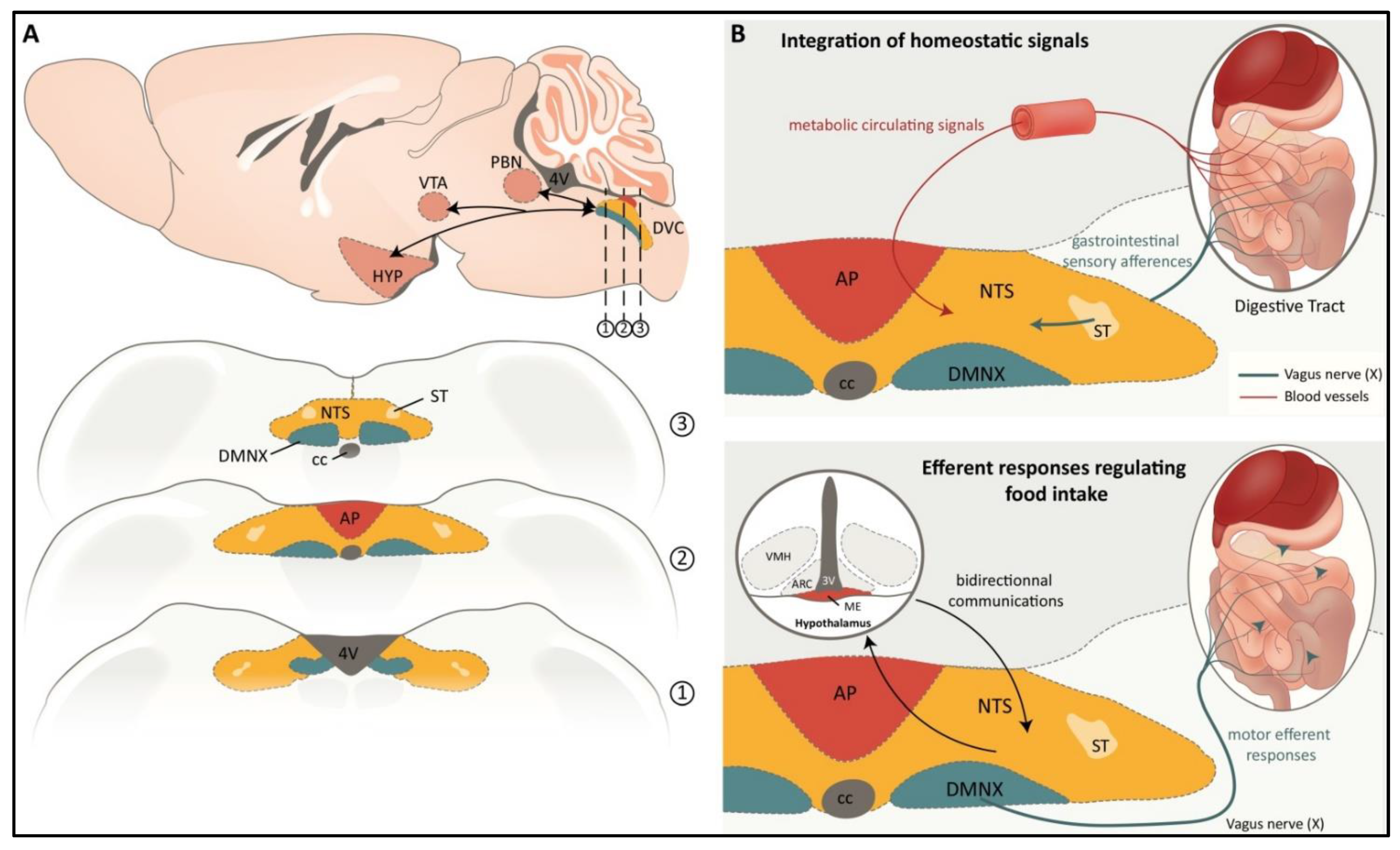{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. The Adult DVC: Structural and Functional Aspects
3. Cellular Diversity and Glial Organization within the Adult DVC
4. DVC Glial Cells, Glucodetection and Regulatory Responses
5. Integration of Viscerosensory Signals: Regulation by NTS Astrocytes
6. Modulation of Food Intake and Weight Gain by DVC Glial Cells
7. Open Questions and Future Avenues
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Obesity and Overweight. 2016. Available online: https://www.who.int/news-room/fact-sheets/de-854tail/obesity-and-overweight (accessed on 31 December 2016).
- Kim, K.S.; Seeley, R.J.; Sandoval, D.A. Signalling from the periphery to the brain that regulates energy homeostasis. Nat. Rev. Neurosci. 2018, 19, 185–196. [Google Scholar] [CrossRef]
- Bouret, S.G. Development of hypothalamic circuits that control food intake and energy balance. In Appetite and Food Intake: Central Control, 2nd ed.; Harris, R.B.S., Ed.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2017; Chapter 7. [Google Scholar] [CrossRef]
- García-Cáceres, C.; Balland, E.; Prevot, V.; Luquet, S.; Woods, S.C.; Koch, M.; Horvath, T.L.; Yi, C.X.; Chowen, J.A.; Araque, A.; et al. Role of astrocytes, microglia, and tanycytes in brain control of systemic metabolism. Nat. Neurosci. 2019, 22, 7–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacDonald, A.J.; Ellacott, K.L.J. Astrocytes in the nucleus of the solitary tract: Contributions to neural circuits controlling physiology. Physiol. Behav. 2020, 223, 112982. [Google Scholar] [CrossRef]
- Dallaporta, M.; Bonnet, M.S.; Horner, K.; Trouslard, J.; Jean, A.; Troadec, J.D. Glial cells of the nucleus tractus solitarius as partners of the dorsal hindbrain regulation of energy balance: A proposal for a working hypothesis. Brain Res. 2010, 1350, 35–42. [Google Scholar] [CrossRef]
- Martinez, D.; Kline, D.D. The role of astrocytes in the nucleus tractus solitarii in maintaining central control of autonomic function. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2021, 320, R418–R424. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, G.J.; Salorio, C.F.; Skoglund, C.; Moran, T.H. Gut vagal afferent lesions increase meal size but do not block gastric preload-induced feeding suppression. Am. J. Physiol. 1999, 276, R1623–R1629. [Google Scholar] [CrossRef]
- Jean, A. The nucleus tractus solitarius: Neuroanatomic, neurochemical and functional aspects. Arch. Int. Physiol. Biochim. Biophys. 1991, 99, A3–A52. [Google Scholar] [CrossRef]
- Norgren, R.; Smith, G.P. Central distribution of subdiaphragmatic vagal branches in the rat. J. Comp. Neurol. 1988, 273, 207–223. [Google Scholar] [CrossRef]
- Sobrino Crespo, C.; Perianes Cachero, A.; Puebla Jiménez, L.; Barrios, V.; Arilla Ferreiro, E. Peptides and food intake. Front. Endocrinol. 2014, 5, 58. [Google Scholar] [CrossRef] [Green Version]
- Andresen, M.C.; Kunze, D.L. Nucleus tractus solitarius-gateway to neural circulatory control. Annu. Rev. Physiol. 1994, 56, 93–116. [Google Scholar] [CrossRef] [PubMed]
- Wright, J.; Campos, C.; Herzog, T.; Covasa, M.; Czaja, K.; Ritter, R.C. Reduction of food intake by cholecystokinin requires activation of hindbrain NMDA-type glutamate receptors. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 301, R448–R455. [Google Scholar] [CrossRef]
- Baptista, V.; Browning, K.N.; Travagli, R.A. Effects of cholecystokinin-8s in the nucleus tractus solitarius of vagally deafferented rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 292, R1092–R1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Lartigue, G. Putative roles of neuropeptides in vagal afferent signaling. Physiol. Behav. 2014, 136, 155–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, C.J.; Hoyda, T.D.; Ferguson, A.V. The area postrema: A brain monitor and integrator of systemic autonomic state. Neuroscientist 2008, 14, 182–194. [Google Scholar] [CrossRef] [PubMed]
- Browning, K.N.; Carson, K.E. Central neurocircuits regulating food intake in response to gut inputs-preclinical evidence. Nutrients 2021, 13, 908. [Google Scholar] [CrossRef]
- Gilg, S.; Lutz, T.A. The orexigenic effect of peripheral ghrelin differs between rats of different age and with different baseline food intake, and it may in part be mediated by the area postrema. Physiol. Behav. 2006, 87, 353–359. [Google Scholar] [CrossRef]
- Dallaporta, M.; Pecchi, E.; Pio, J.; Jean, A.; Horner, K.C.; Troadec, J.D. Expression of leptin receptor by glial cells of the nucleus tractus solitarius: Possible involvement in energy homeostasis. J. Neuroendocrinol. 2009, 21, 57–67. [Google Scholar] [CrossRef]
- Durkee, C.A.; Araque, A. Diversity and specificity of astrocyte-neuron communication. Neuroscience 2019, 396, 73–78. [Google Scholar] [CrossRef]
- Bouzioukh, F.; Tell, F.; Jean, A.; Rougon, G. NMDA receptor and nitric oxide synthase activation regulate polysialylated neural cell adhesion molecule expression in adult brainstem synapses. J. Neurosci. 2001, 21, 4721–4730. [Google Scholar] [CrossRef] [Green Version]
- Pecchi, E.; Dallaporta, M.; Charrier, C.; Pio, J.; Jean, A.; Moyse, E.; Troadec, J.D. Glial fibrillary acidic protein (GFAP)-positive radial-like cells are present in the vicinity of proliferative progenitors in the nucleus tractus solitarius of adult rat. J. Comp. Neurol. 2007, 501, 353–368. [Google Scholar] [CrossRef]
- Litvin, D.G.; Denstaedt, S.J.; Borkowski, L.F.; Nichols, N.L.; Dick, T.E.; Smith, C.B.; Jacono, F.J. Peripheral-to-central immune communication at the area postrema glial-barrier following bleomycin-induced sterile lung injury in adult rats. Brain Behav. Immun. 2020, 87, 610–633. [Google Scholar] [CrossRef] [PubMed]
- Tashiro, Y.; Kawai, Y. Glial coverage of the small cell somata in the rat nucleus of tractus solitarius during postnatal development. Glia 2007, 55, 1619–1629. [Google Scholar] [CrossRef]
- Chounlamountry, K.; Kessler, J.P. The ultrastructure of perisynaptic glia in the nucleus tractus solitarii of the adult rat: Comparison between single synapses and multisynaptic arrangements. Glia 2011, 59, 655–663. [Google Scholar] [CrossRef]
- Hofmann, G.C.; Hasser, E.M.; Kline, D. Unilateral vagatomy alters astrocyte and microglial morphology in the nucleus tractus solitarii in of the rat. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2021, 320, R945–R959. [Google Scholar] [CrossRef] [PubMed]
- Tadmouri, A.; Champagnat, J.; Morin-Surun, M.P. Activation of microglia and astrocytes in the nucleus tractus solitarius during ventilatory acclimatization to 10% hypoxia in unanesthetized mice. J. Neurosci. Res. 2014, 92, 627–633. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Chen, F.; Fang, Z.; Zhao, H.; Zhan, C.; Li, C.; He, Y.; Huang, C.; Long, L.; Lai, K. Glial activation and inflammation in the NTS in a rat model after exposure to diesel exhaust particles. Environ. Toxicol. Pharmacol. 2021, 83, 103584. [Google Scholar] [CrossRef]
- Chounlamountry, K.; Boyer, B.; Penalba, V.; François-Bellan, A.M.; Bosler, O.; Kessler, J.P.; Strube, C. Remodeling of glial coverage of glutamatergic synapses in the rat nucleus tractus solitarii after ozone inhalation. J. Neurochem. 2015, 134, 857–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacDonald, A.J.; Holmes, F.E.; Beall, C.; Pickering, A.E.; Ellacott, K.L.J. Regulation of food intake by astrocytes in the brainstem dorsal vagal complex. Glia 2020, 68, 1241–1254. [Google Scholar] [CrossRef] [Green Version]
- Maolood, N.; Meister, B. Protein components of the blood-brain barrier (BBB) in the brainstem area postrema-nucleus tractus solitarius region. J. Chem. Neuroanat. 2009, 37, 182–195. [Google Scholar] [CrossRef]
- Sundholm-Peters, N.L.; Yang, H.K.C.; Goings, G.E.; Walker, A.S.; Szele, F.G. Radial glia-like cells at the base of the lateral ventricles in adult mice. J. Neurocytol. 2004, 33, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Kaye, J.A.; Cai, Z.; Wang, Y.; Prescott, S.L.; Liberles, S.D. Area postrema cell types that mediate nausea-associated behaviors. Neuron 2021, 109, 461–472.e5. [Google Scholar] [CrossRef]
- Ludwig, M.Q.; Cheng, W.; Gordian, D.; Lee, J.; Paulsen, S.J.; Hansen, S.N.; Egerod, K.L.; Barkholt, P.; Rhodes, C.J.; Secher, A.; et al. A genetic map of the mouse dorsal vagal complex and its role in obesity. Nat. Metab. 2021, 3, 530–545. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.N.; Macosko, E.Z.; Fenselau, H.; Pers, T.H.; Lyubetskaya, A.; Tenen, D.; Goldman, M.; Verstegen, A.M.J.; Resch, J.M.; McCarroll, S.A.; et al. A molecular census of arcuate hypothalamus and median eminence cell types. Nat. Neurosci. 2017, 20, 484–496. [Google Scholar] [CrossRef] [Green Version]
- Peruzzo, B.; Pastor, F.E.; Blázquez, J.L.; Schöbitz, K.; Peláez, B.; Amat, P.; Rodríguez, E.M. A second look at the barriers of the medial basal hypothalamus. Exp. Brain Res. 2000, 132, 10–26. [Google Scholar] [CrossRef] [PubMed]
- Mullier, A.; Bouret, S.G.; Prevot, V.; Dehouck, B. Differential distribution of tight junction proteins suggests a role for tanycytes in blood-hypothalamus barrier regulation in the adult mouse brain. J. Comp. Neurol. 2010, 518, 943–962. [Google Scholar] [CrossRef]
- Rodríguez, E.M.; Blázquez, J.L.; Guerra, M. The design of barriers in the hypothalamus allows the median eminence and the arcuate nucleus to enjoy private milieus: The former opens to the portal blood and the latter to the cerebrospinal fluid. Peptides 2010, 31, 757–776. [Google Scholar] [CrossRef] [PubMed]
- Maness, L.M.; Kastin, A.J.; Banks, W.A. Relative contributions of a CVO and the microvascular bed to delivery of blood-borne IL-1alpha to the brain. Am. J. Physiol. 1998, 275, E207–E212. [Google Scholar] [CrossRef]
- Willis, C.L.; Garwood, C.J.; Ray, D.E. A size selective vascular barrier in the rat area postrema formed by perivascular macrophages and the extracellular matrix. Neuroscience 2007, 150, 498–509. [Google Scholar] [CrossRef]
- Wang, Q.P.; Guan, J.L.; Pan, W.; Kastin, A.J.; Shioda, S. A diffusion barrier between the area postrema and nucleus tractus solitarius. Neurochem. Res. 2008, 33, 2035–2043. [Google Scholar] [CrossRef]
- Lebrun, B.; Barbot, M.; Tonon, M.C.; Prévot, V.; Leprince, J.; Troadec, J.D. Glial endozepines and energy balance: Old peptides with new tricks. Glia 2021, 69, 1079–1093. [Google Scholar] [CrossRef] [PubMed]
- Takagi, S.; Furube, E.; Nakano, Y.; Morita, M.; Miyata, S. Microglia are continuously activated in the circumventricular organs of mouse brain. J. Neuroimmunol. 2019, 331, 74–86. [Google Scholar] [CrossRef] [PubMed]
- Butler, M.J.; Perrini, A.A.; Eckel, L.A. Estradiol treatment attenuates high fat diet-induced microgliosis in ovariectomized rats. Horm. Behav. 2020, 120, 104675. [Google Scholar] [CrossRef] [PubMed]
- Lawson, L.J.; Perry, V.H.; Dri, P.; Gordon, S. Heterogeneity in the distribution and morphology of microglia in the normal adult mouse brain. Neuroscience 1990, 39, 151–170. [Google Scholar] [CrossRef]
- Perry, V.H. A revised view of the central nervous system microenvironment and major histocompatibility complex class II antigen presentation. J. Neuroimmunol. 1998, 90, 113–121. [Google Scholar] [CrossRef]
- Vaughn, A.C.; Cooper, E.M.; DiLorenzo, P.M.; O’Loughlin, L.J.; Konkel, M.E.; Peters, J.H.; Hajnal, A.; Sen, T.; Lee, S.H.; de La Serre, C.B.; et al. Energy-dense diet triggers changes in gut microbiota, reorganization of gut-brain vagal communication and increases body fat accumulation. Acta Neurobiol. Exp. 2017, 77, 18–30. [Google Scholar] [CrossRef] [Green Version]
- Watts, A.G.; Donovan, C.M. Sweet talk in the brain: Glucosensing, neural networks, and hypoglycemic counterregulation. Front. Neuroendocrinol. 2010, 31, 32–43. [Google Scholar] [CrossRef] [Green Version]
- Cottrell, G.T.; Ferguson, A.V. Sensory circumventricular organs: Central roles in integrated autonomic regulation. Regul. Pept. 2004, 117, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Yettefti, K.; Orsini, J.C.; El Ouazzani, T.; Himmi, T.; Boyer, A.; Perrin, J. Sensitivity of nucleus tractus solitarius neurons to induced moderate hyperglycemia, with special reference to catecholaminergic regions. J. Auton. Nerv. Syst. 1995, 51, 191–197. [Google Scholar] [CrossRef]
- Briski, K.P.; Marshall, E.S. Caudal brainstem Fos expression is restricted to periventricular catecholamine neuron-containing loci following intraventricular administration of 2-deoxy-D-glucose. Exp. Brain Res. 2000, 133, 547–551. [Google Scholar] [CrossRef]
- Himmi, T.; Dallaporta, M.; Perrin, J.; Orsini, J.C. Neuronal responses to delta 9-tetrahydrocannabinol in the solitary tract nucleus. Eur. J. Pharmacol. 1996, 312, 273–279. [Google Scholar] [CrossRef]
- Dallaporta, M.; Himmi, T.; Perrin, J.; Orsini, J.C. Solitary tract nucleus sensitivity to moderate changes in glucose level. Neuroreport 1999, 10, 2657–2660. [Google Scholar] [CrossRef]
- Madden, C.J.; Stocker, S.D.; Sved, A.F. Attenuation of homeostatic responses to hypotension and glucoprivation after destruction of catecholaminergic rostral ventrolateral medulla neurons. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 291, R751–R759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritter, S.; Llewellyn-Smith, I.; Dinh, T.T. Subgroups of hindbrain catecholamine neurons are selectively activated by 2-deoxy-D-glucose induced metabolic challenge. Brain Res. 1998, 805, 41–54. [Google Scholar] [CrossRef]
- Ritter, S.; Dinh, T.T.; Zhang, Y. Localization of hindbrain glucoreceptive sites controlling food intake and blood glucose. Brain Res. 2000, 856, 37–47. [Google Scholar] [CrossRef]
- Lamy, C.M.; Sanno, H.; Labouèbe, G.; Picard, A.; Magnan, C.; Chatton, J.Y.; Thorens, B. Hypoglycemia-activated GLUT2 neurons of the nucleus tractus solitarius stimulate vagal activity and glucagon secretion. Cell Metab. 2014, 19, 527–538. [Google Scholar] [CrossRef] [Green Version]
- Andrew, S.F.; Dinh, T.T.; Ritter, S. Localized glucoprivation of hindbrain sites elicits corticosterone and glucagon secretion. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 292, R1792–R1798. [Google Scholar] [CrossRef] [PubMed]
- Aklan, I.; Sayar Atasoy, N.; Yavuz, Y.; Ates, T.; Coban, I.; Koksalar, F.; Filiz, G.; Topcu, I.C.; Oncul, M.; Dilsiz, P.; et al. NTS catecholamine neurons mediate hypoglycemic hunger via medial hypothalamic feeding pathways. Cell Metab. 2020, 31, 313–326. [Google Scholar] [CrossRef]
- Roberts, B.L.; Zhu, M.; Zhao, H.; Dillon, C.; Appleyard, S.M. High glucose increases action potential firing of catecholamine neurons in the nucleus of the solitary tract by increasing spontaneous glutamate inputs. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2017, 313, R229–R239. [Google Scholar] [CrossRef] [Green Version]
- Lin, L.H.; Moore, S.A.; Jones, S.Y.; McGlashon, J.; Talman, W.T. Astrocytes in the rat nucleus tractus solitarii are critical for cardiovascular reflex control. J. Neurosci. 2013, 33, 18608–18617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogers, R.C.; Hermann, G.E. Hindbrain astrocytes and glucose counter-regulation. Physiol. Behav. 2019, 204, 140–150. [Google Scholar] [CrossRef]
- Young, J.K.; Baker, J.H.; Montes, M.I. The brain response to 2-deoxy glucose is blocked by a glial drug. Pharmacol. Biochem. Behav. 2000, 67, 233–239. [Google Scholar] [CrossRef]
- Rogers, R.C.; Ritter, S.; Hermann, G.E. Hindbrain cytoglucopenia-induced increases in systemic blood glucose levels by 2-deoxyglucose depend on intact astrocytes and adenosine release. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 310, R1102–R1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermann, G.E.; Viard, E.; Rogers, R.C. Hindbrain glucoprivation effects on gastric vagal reflex circuits and gastric motility in the rat are suppressed by the astrocyte inhibitor fluorocitrate. J. Neurosci. 2014, 34, 10488–10496. [Google Scholar] [CrossRef] [Green Version]
- McDougal, D.H.; Hermann, G.E.; Rogers, R.C. Astrocytes in the nucleus of the solitary tract are activated by low glucose or glucoprivation: Evidence for glial involvement in glucose homeostasis. Front. Neurosci. 2013, 7, 249. [Google Scholar] [CrossRef] [Green Version]
- McDougal, D.H.; Viard, E.; Hermann, G.H.; Rogers, R.C. Astrocytes in the hindbrain detect glucoprivation and regulate gastric motility. Auton. Neurosci. 2013, 175, 61–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogers, R.C.; McDougal, D.H.; Ritter, S.; Qualls-Creekmore, E.; Hermann, G.E. Response of catecholaminergic neurons in the mouse hindbrain to glucoprivic stimuli is astrocyte dependent. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 315, R153–R164. [Google Scholar] [CrossRef] [PubMed]
- Rogers, R.C.; Burke, S.J.; Collier, J.J.; Ritter, S.; Hermann, G.H. Evidence that hindbrain astrocytes in the rat detect low glucose with a glucose transporter 2-phospholipase C-calcium release mechanism. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2020, 318, R38–R48. [Google Scholar] [CrossRef] [Green Version]
- Leloup, C.; Arluison, M.; Lepetit, N.; Cartier, N.; Marfaing-Jallat, P.; Ferré, P.; Pénicaud, L. Glucose transporter 2 (GLUT 2): Expression in specific brain nuclei. Brain Res. 1994, 638, 221–226. [Google Scholar] [CrossRef]
- Burcelin, R.; Thorens, B. Evidence that extrapancreatic GLUT2-dependent glucose sensors control glucagon secretion. Diabetes 2001, 50, 1282–1289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marty, N.; Dallaporta, M.; Foretz, M.; Emery, M.; Tarussio, D.; Bady, I.; Binnert, C.; Beermann, F.; Thorens, B. Regulation of glucagon secretion by glucose transporter type 2 (glut2) and astrocyte-dependent glucose sensors. J. Clin. Investig. 2005, 115, 3545–3553. [Google Scholar] [CrossRef] [Green Version]
- Bady, I.; Marty, N.; Dallaporta, M.; Emery, M.; Gyger, J.; Tarussio, D.; Foretz, M.; Thorens, B. Evidence from glut2-null mice that glucose is a critical physiological regulator of feeding. Diabetes 2006, 55, 988–995. [Google Scholar] [CrossRef]
- McDougal, D.H.; Hermann, G.E.; Rogers, R.C. Vagal afferent stimulation activates astrocytes in the nucleus of the solitary tract via AMPA receptors: Evidence of an atypical neural-glial interaction in the brainstem. J. Neurosci. 2011, 31, 14037–14045. [Google Scholar] [CrossRef] [PubMed]
- Pasti, L.; Volterra, A.; Pozzan, T.; Carmignoto, G. Intracellular calcium oscillations in astrocytes: A highly plastic, bidirectional form of communication between neurons and astrocytes in situ. J. Neurosci. 1997, 17, 7817–7830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araque, A.; Martín, E.D.; Perea, G.; Arellano, J.I.; Buño, W. Synaptically released acetylcholine evokes Ca2+ elevations in astrocytes in hippocampal slices. J. Neurosci. 2002, 22, 2443–2450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Ascenzo, M.; Fellin, T.; Terunuma, M.; Revilla-Sanchez, R.; Meaney, D.F.; Auberson, Y.P.; Moss, S.J.; Haydon, P.G. mGluR5 stimulates gliotransmission in the nucleus accumbens. Proc. Natl. Acad. Sci. USA 2007, 104, 1995–2000. [Google Scholar] [CrossRef] [Green Version]
- Accorsi-Mendonça, D.; Zoccal, D.B.; Bonagamba, L.G.H.; Machado, B.H. Glial cells modulate the synaptic transmission of NTS neurons sending projections to ventral medulla of Wistar rats. Physiol. Rep. 2013, 1, e00080. [Google Scholar] [CrossRef] [Green Version]
- Accorsi-Mendonça, D.; Bonagamba, L.G.H.; Machado, B.H. Astrocytic modulation of glutamatergic synaptic transmission is reduced in NTS of rats submitted to short-term sustained hypoxia. J. Neurophysiol. 2019, 121, 1822–1830. [Google Scholar] [CrossRef]
- Accorsi-Mendonça, D.; Almado, C.E.L.; Bonagamba, L.G.H.; Castania, J.A.; Moraes, D.J.A.; Machado, B.H. Enhanced firing in NTS induced by short-term sustained hypoxia is modulated by glia-neuron interaction. J. Neurosci. 2015, 35, 6903–6917. [Google Scholar] [CrossRef] [Green Version]
- Naskar, K.; Stern, J.E. A functional coupling between extrasynaptic NMDA receptors and A-type K+ channels under astrocyte control regulates hypothalamic neurosecretory neuronal activity. J. Physiol. 2014, 592, 2813–2827. [Google Scholar] [CrossRef]
- Matott, M.P.; Ruyle, B.C.; Hasser, E.M.; Kline, D.D. Excitatory amino acid transporters tonically restrain NTS synaptic and neuronal activity to modulate cardiorespiratory function. J. Neurophysiol. 2016, 115, 1691–1702. [Google Scholar] [CrossRef] [Green Version]
- Matott, M.P.; Kline, D.D.; Hasser, E.M. Glial EAAT2 regulation of extracellular NTS glutamate critically controls neuronal activity and cardiorespiratory reflexes. J. Physiol. 2017, 595, 6045–6063. [Google Scholar] [CrossRef]
- Yamamoto, K.; Mifflin, S. Inhibition of glial glutamate transporter GLT1 in the nucleus of the solitary tract attenuates baroreflex control of sympathetic nerve activity and heart rate. Physiol. Rep. 2018, 6, e13877. [Google Scholar] [CrossRef]
- Pellerin, L.; Magistretti, P.J. Glutamate uptake into astrocytes stimulates aerobic glycolysis: A mechanism coupling neuronal activity to glucose utilization. Proc. Natl. Acad. Sci. USA 1994, 91, 10625–10629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagase, M.; Takahashi, Y.; Watabe, A.M.; Kubo, Y.; Kato, F. On-site energy supply at synapses through monocarboxylate transporters maintains excitatory synaptic transmission. J. Neurosci. 2014, 34, 2605–2617. [Google Scholar] [CrossRef] [Green Version]
- Thek, K.R.; Ong, S.J.M.; Carter, D.C.; Bassi, J.K.; Allen, A.M.; McDougall, S.J. Extensive inhibitory gating of viscerosensory signals by a sparse network of somatostatin neurons. J. Neurosci. 2019, 39, 8038–8050. [Google Scholar] [CrossRef]
- Mariotti, L.; Losi, G.; Lia, A.; Melone, M.; Chiavegato, A.; Gómez-Gonzalo, M.; Sessolo, M.; Bovetti, S.; Forli, A.; Zonta, M.; et al. Interneuron-specific signaling evokes distinctive somatostatin-mediated responses in adult cortical astrocytes. Nat. Commun. 2018, 9, 82. [Google Scholar] [CrossRef]
- Matos, M.; Bosson, A.; Riebe, I.; Reynell, C.; Vallée, J.; Laplante, I.; Panatier, A.; Robitaille, R.; Lacaille, J.C. Astrocytes detect and upregulate transmission at inhibitory synapses of somatostatin interneurons onto pyramidal cells. Nat. Commun. 2018, 9, 4254. [Google Scholar] [CrossRef] [PubMed]
- Pizarroso, N.A.; Fuciños, P.; Gonçalves, C.; Pastrana, L.; Amado, I.R. A review on the role of food-derived bioactive molecules and the microbiota-gut-brain axis in satiety regulation. Nutrients 2021, 13, 632. [Google Scholar] [CrossRef]
- Knudsen, L.B.; Lau, J. The discovery and development of liraglutide and semaglutide. Front. Endocrinol. 2019, 10, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakouri, A.; Kanti, G.; Kapantais, E.; Kokkinos, A.; Lanaras, L.; Farajian, P.; Galanakis, C.; Georgantopoulos, G.; Vlahos, N.F.; Mastorakos, G.; et al. New incretin combination treatments under investigation in obesity and metabolism: A systematic review. Pharmaceuticals 2021, 14, 869. [Google Scholar] [CrossRef]
- Jensen, C.B.; Pyke, C.; Rasch, M.G.; Dahl, A.B.; Knudsen, L.B.; Secher, A. Characterization of the Glucagonlike Peptide-1 Receptor in male mouse brain using a novel antibody and in situ hybridization. Endocrinology 2018, 159, 665–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabery, S.; Salinas, C.G.; Paulsen, S.J.; Ahnfelt-Rønne, J.; Alanentalo, T.; Baquero, A.F.; Buckley, S.T.; Farkas, E.; Fekete, C.; Frederiksen, K.S.; et al. Semaglutide lowers body weight in rodents via distributed neural pathways. JCI Insight 2020, 5, e133429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farkas, E.; Szilvásy-Szabó, A.; Ruska, Y.; Sinkó, R.; Rasch, M.G.; Egebjerg, T.; Pyke, C.; Gereben, B.; Knudsen, L.B.; Fekete, C. Distribution and ultrastructural localization of the glucagon-like peptide-1 receptor (GLP-1R) in the rat brain. Brain Struct. Funct. 2021, 226, 225–245. [Google Scholar] [CrossRef] [PubMed]
- Reiner, D.J.; Mietlicki-Baase, E.G.; McGrath, L.E.; Zimmer, D.J.; Bence, K.K.; Sousa, G.L.; Konanur, V.R.; Krawczyk, J.; Burk, D.H.; Kanoski, S.E.; et al. Astrocytes regulate GLP-1 receptor-mediated effects on energy balance. J. Neurosci. 2016, 36, 3531–3540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludwig, M.Q.; Todorov, P.V.; Egerod, K.L.; Olson, D.P.; Pers, T.H. Single-cell mapping of GLP-1 and GIP receptor expression in the dorsal vagal complex. Diabetes 2021, 70, 1945–1955. [Google Scholar] [CrossRef]
- Dowsett, G.K.C.; Lam, B.Y.H.; Tadross, J.A.; Cimino, I.; Rimmington, D.; Coll, A.P.; Polex-Wolf, J.; Knudsen, L.B.; Pyke, C.; Yeo, G.S.H. A survey of the mouse hindbrain in the fed and fasted states using single-nucleus RNA sequencing. Mol. Metab. 2021, 53, 101240. [Google Scholar] [CrossRef] [PubMed]
- Timper, K.; Del Río-Martín, A.; Cremer, A.L.; Bremser, S.; Alber, J.; Giavalisco, P.; Varela, L.; Heilinger, C.; Nolte, H.; Trifunovic, A.; et al. GLP-1 receptor signaling in astrocytes regulates fatty acid oxidation, mitochondrial integrity, and function. Cell Metab. 2020, 31, 1189–1205.e13. [Google Scholar] [CrossRef]
- Buyse, M.; Ovesjö, M.L.; Goïot, H.; Guilmeau, S.; Péranzi, G.; Moizo, L.; Walker, F.; Lewin, M.J.; Meister, B.; Bado, A. Expression and regulation of leptin receptor proteins in afferent and efferent neurons of the vagus nerve. Eur. J. Neurosci. 2001, 14, 64–72. [Google Scholar] [CrossRef]
- Hsuchou, H.; He, Y.; Kastin, A.J.; Tu, H.; Markadakis, E.N.; Rogers, R.C.; Fossier, P.B.; Pan, W. Obesity induces functional astrocytic leptin receptors in hypothalamus. Brain 2009, 132, 889–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsuchou, H.; Pan, W.; Barnes, M.J.; Kastin, A.J. Leptin receptor mRNA in rat brain astrocytes. Peptides 2009, 30, 2275–2280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stein, L.M.; Lhamo, R.; Cao, A.; Workinger, J.; Tinsley, I.; Doyle, R.P.; Grill, H.J.; Hermann, G.E.; Rogers, R.C.; Hayes, M.R. Dorsal vagal complex and hypothalamic glia differentially respond to leptin and energy balance dysregulation. Transl. Psychiatry 2020, 10, 90. [Google Scholar] [CrossRef] [Green Version]
- Brüning, J.C.; Gautam, D.; Burks, D.J.; Gillette, J.; Schubert, M.; Orban, P.C.; Klein, R.; Krone, W.; Müller-Wieland, D.; Kahn, C.R. Role of brain insulin receptor in control of body weight and reproduction. Science 2000, 289, 2122–2125. [Google Scholar] [CrossRef]
- Obici, S.; Zhang, B.B.; Karkanias, G.; Rossetti, L. Hypothalamic insulin signaling is required for inhibition of glucose production. Nat. Med. 2002, 8, 1376–1382. [Google Scholar] [CrossRef] [PubMed]
- Pocai, A.; Lam, T.K.; Gutierrez-Juarez, R.; Obici, S.; Schwartz, G.J.; Bryan, J.; Aguilar-Bryan, L.; Rossetti, L. Hypothalamic K(ATP) channels control hepatic glucose production. Nature 2005, 434, 1026–1031. [Google Scholar] [CrossRef]
- Obici, S.; Feng, Z.; Karkanias, G.; Baskin, D.G.; Rossetti, L. Decreasing hypothalamic insulin receptors causes hyperphagia and insulin resistance in rats. Nat. Neurosci. 2002, 5, 566–572. [Google Scholar] [CrossRef] [PubMed]
- Filippi, B.M.; Yang, C.S.; Tang, C.; Lam, T.K. Insulin activates Erk1/2 signaling in the dorsal vagal complex to inhibit glucose production. Cell Metab. 2012, 16, 500–510. [Google Scholar] [CrossRef] [Green Version]
- Abraham, M.A.; Filippi, B.M.; Kang, G.M.; Kim, M.S.; Lam, T.K. Insulin action in the hypothalamus and dorsal vagal complex. Exp. Physiol. 2014, 99, 1104–1109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahn, B.B. Type 2 diabetes: When insulin secretion fails to compensate for insulin resistance. Cell 1998, 92, 593–596. [Google Scholar] [CrossRef] [Green Version]
- Cai, D.; Khor, S. “Hypothalamic microinflammation” paradigm in aging and metabolic diseases. Cell Metab. 2019, 30, 19–35. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Diano, S. Mitochondrial dynamics and hypothalamic regulation of metabolism. Endocrinology 2018, 159, 3596–3604. [Google Scholar] [CrossRef] [PubMed]
- Jheng, H.F.; Tsai, P.J.; Guo, S.M.; Kuo, L.H.; Chang, C.S.; Su, I.J.; Chang, C.R.; Tsai, Y.S. Mitochondrial fission contributes to mitochondrial dysfunction and insulin resistance in skeletal muscle. Mol. Cell. Biol. 2012, 32, 309–319. [Google Scholar] [CrossRef] [Green Version]
- Gao, A.W.; Cantó, C.; Houtkooper, R.H. Mitochondrial response to nutrient availability and its role in metabolic disease. EMBO Mol. Med. 2014, 6, 580–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, I.D.; Brookler, K.H.; Kyriakidou, Y.; Elliott, B.T.; Crofts, C.A.P. Metabolic phenotypes and step by step evolution of type 2 diabetes: A New paradigm. Biomedicines 2021, 9, 800. [Google Scholar] [CrossRef]
- Wang, L.; Ishihara, T.; Ibayashi, Y.; Tatsushima, K.; Setoyama, D.; Hanada, Y.; Takeichi, Y.; Sakamoto, S.; Yokota, S.; Mihara, K.; et al. Disruption of mitochondrial fission in the liver protects mice from diet-induced obesity and metabolic deterioration. Diabetologia 2015, 58, 2371–2380. [Google Scholar] [CrossRef] [Green Version]
- Dietrich, M.O.; Liu, Z.W.; Horvath, T.L. Mitochondrial dynamics controlled by mitofusins regulate Agrp neuronal activity and diet-induced obesity. Cell 2013, 155, 188–199. [Google Scholar] [CrossRef] [Green Version]
- Schneeberger, M.; Dietrich, M.O.; Sebastián, D.; Imbernón, M.; Castaño, C.; Garcia, A.; Esteban, Y.; Gonzales-Franquesa, A.; Castrillon Rodriguez, I.; Bortolozzi, A.; et al. Mitofusin 2 in POMC neurons connects ER stress with leptin resistance and energy imbalance. Cell 2013, 155, 172–187. [Google Scholar] [CrossRef] [Green Version]
- Santoro, A.; Campolo, M.; Liu, C.; Sesaki, H.; Meli, R.; Liu, Z.W.; Kim, J.D.; Diano, S. DRP1 suppresses leptin and glucose sensing of POMC neurons. Cell Metab. 2017, 25, 647–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filippi, B.M.; Abraham, M.A.; Silva, P.N.; Rasti, M.; LaPierre, M.P.; Bauer, P.V.; Rocheleau, J.V.; Lam, T.K.T. Dynamin-Related Protein 1-Dependent mitochondrial fission changes in the dorsal vagal complex regulate insulin action. Cell Rep. 2017, 18, 2301–2309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, B.; New, L.E.; Griffiths, J.C.; Deuchars, J.; Filippi, B.M. Inhibition of mitochondrial fission and iNOS in the dorsal vagal complex protects from overeating and weight gain. Mol. Metab. 2021, 43, 101123. [Google Scholar] [CrossRef] [PubMed]
- Petrelli, F.; Bezzi, P. Novel insights into gliotransmitters. Curr. Opin. Pharmacol. 2016, 26, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Kittner, H.; Franke, H.; Harsch, J.I.; El-Ashmawy, I.M.; Seidel, B.; Krügel, U.; Illes, P. Enhanced food intake after stimulation of hypothalamic P2Y1 receptors in rats: Modulation of feeding behaviour by extracellular nucleotides. Eur. J. Neurosci. 2006, 24, 2049–2056. [Google Scholar] [CrossRef]
- Colldén, G.; Mangano, C.; Meister, B. P2X2 purinoreceptor protein in hypothalamic neurons associated with the regulation of food intake. Neuroscience 2010, 171, 62–78. [Google Scholar] [CrossRef] [PubMed]
- Stanley, B.G.; Urstadt, K.R.; Charles, J.R.; Kee, T. Glutamate and GABA in lateral hypothalamic mechanisms controlling food intake. Physiol. Behav. 2011, 104, 40–46. [Google Scholar] [CrossRef]
- Giaume, C.; Leybaert, L.; Naus, C.C.; Sáez, J.C. Connexin and pannexin hemichannels in brain glial cells: Properties, pharmacology, and roles. Front. Pharmacol. 2013, 4, 88. [Google Scholar] [CrossRef] [Green Version]
- Giaume, C.; Liu, X. From a glial syncytium to a more restricted and specific glial networking. J. Physiol. 2012, 106, 34–39. [Google Scholar] [CrossRef]
- Montero, T.D.; Orellana, J.A. Hemichannels: New pathways for gliotransmitter release. Neuroscience 2015, 286, 45–59. [Google Scholar] [CrossRef]
- Guillebaud, F.; Barbot, M.; Barbouche, R.; Brézun, J.M.; Poirot, K.; Vasile, F.; Lebrun, B.; Rouach, N.; Dallaporta, M.; Gaige, S.; et al. Blockade of glial connexin 43 hemichannels reduces food intake. Cells 2020, 9, 2387. [Google Scholar] [CrossRef]
- Tonon, M.C.; Vaudry, H.; Chuquet, J.; Guillebaud, F.; Fan, J.; Masmoudi-Kouki, O.; Vaudry, D.; Lanfray, D.; Morin, F.; Prevot, V.; et al. Endozepines and their receptors: Structure, functions and pathophysiological significance. Pharmacol. Ther. 2020, 208, 107386. [Google Scholar] [CrossRef]
- De Mateos-Verchere, J.G.; Leprince, J.; Tonon, M.C.; Vaudry, H.; Costentin, J. The octadecaneuropeptide [diazepam-binding inhibitor (33–50)] exerts potent anorexigenic effects in rodents. Eur. J. Pharmacol. 2001, 414, 225–231. [Google Scholar] [CrossRef]
- do Rego, J.C.; Orta, M.H.; Leprince, J.; Tonon, M.C.; Vaudry, H.; Costentin, J. Pharmacological characterization of the receptor mediating the anorexigenic action of the octadecaneuropeptide: Evidence for an endozepinergic tone regulating food intake. Neuropsychopharmacology 2007, 32, 1641–1648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanfray, D.; Arthaud, S.; Ouellet, J.; Compère, V.; Do Rego, J.L.; Leprince, J.; Lefranc, B.; Castel, H.; Bouchard, C.; Monge-Roffarello, B.; et al. Gliotransmission and brain glucose sensing: Critical role of endozepines. Diabetes 2013, 62, 801–810. [Google Scholar] [CrossRef] [Green Version]
- Bouyakdan, K.; Martin, H.; Liénard, F.; Budry, L.; Taib, B.; Rodaros, D.; Chrétien, C.; Biron, E.; Husson, Z.; Cota, D.; et al. The gliotransmitter ACBP controls feeding and energy homeostasis via the melanocortin system. J. Clin. Investig. 2019, 129, 2417–2430. [Google Scholar] [CrossRef]
- Guillebaud, F.; Duquenne, M.; Djelloul, M.; Pierre, C.; Poirot, K.; Roussel, G.; Riad, S.; Lanfray, D.; Morin, F.; Jean, A.; et al. Glial endozepines reverse high-fat diet-induced obesity by enhancing hypothalamic response to peripheral leptin. Mol. Neurobiol. 2020, 57, 3307–3333. [Google Scholar] [CrossRef]
- Bouyakdan, K.; Taïb, B.; Budry, L.; Zhao, S.; Rodaros, D.; Neess, D.; Mandrup, S.; Faergeman, N.J.; Alquier, T. A novel role for central ACBP/DBI as a regulator of long-chain fatty acid metabolism in astrocytes. J. Neurochem. 2015, 133, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Guillebaud, F.; Girardet, C.; Abysique, A.; Gaigé, S.; Barbouche, R.; Verneuil, J.; Jean, A.; Leprince, J.; Tonon, M.C.; Dallaporta, M.; et al. Glial endozepines inhibit feeding-related autonomic functions by acting at the brainstem level. Front. Neurosci. 2017, 11, 308. [Google Scholar] [CrossRef] [PubMed]
- Lalo, U.; Koh, W.; Lee, C.J.; Pankratov, Y. The tripartite glutamatergic synapse. Neuropharmacology 2021, 199, 108758. [Google Scholar] [CrossRef] [PubMed]
- Martinez, D.; Rogers, R.C.; Hermann, G.E.; Hasser, E.M.; Kline, D.D. Astrocytic glutamate transporters reduce the neuronal and physiological influence of metabotropic glutamate receptors in nucleus tractus solitarii. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2020, 318, R545–R564. [Google Scholar] [CrossRef] [PubMed]
- Martinez, D.; Rogers, R.C.; Hasser, E.M.; Hermann, G.E.; Kline, D.D. Loss of excitatory amino acid transporter restraint following chronic intermittent hypoxia contributes to synaptic alterations in nucleus tractus solitarii. J. Neurophysiol. 2020, 123, 2122–2135. [Google Scholar] [CrossRef]
- Clyburn, C.; Travagli, R.A.; Browning, K.N. Acute high-fat diet upregulates glutamatergic signaling in the dorsal motor nucleus of the vagus. Am. J. Physiol. Gastrointest. Liver Physiol. 2018, 314, G623–G634. [Google Scholar] [CrossRef] [PubMed]
- Clyburn, C.; Travagli, R.A.; Arnold, A.C.; Browning, K.N. DMV extrasynaptic NMDA receptors regulate caloric intake in rats. JCI Insight 2021, 6, e139785. [Google Scholar] [CrossRef]
- Thaler, J.P.; Yi, C.X.; Schur, E.A.; Guyenet, S.J.; Hwang, B.H.; Dietrich, M.O.; Zhao, X.; Sarruf, D.A.; Izgur, V.; Maravilla, K.R.; et al. Obesity is associated with hypothalamic injury in rodents and humans. J. Clin. Investig. 2012, 122, 153–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buckman, L.B.; Thompson, M.M.; Moreno, H.N.; Ellacott, K.L.J. Regional astrogliosis in the mouse hypothalamus in response to obesity. J. Comp. Neurol. 2013, 521, 1322–1333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elizondo-Vega, R.J.; Recabal, A.; Oyarce, K. Nutrient sensing by hypothalamic tanycytes. Front. Endocrinol. 2019, 10, 244. [Google Scholar] [CrossRef] [PubMed]
- Orellana, J.A.; Saez, P.J.; Cortes-Campos, C.; Elizondo, R.J.; Shoji, K.F.; Contreras-Duarte, S.; Figueroa, V.; Velarde, V.; Jiang, J.X.; Nualart, F.; et al. Glucose increases intracellular free Ca(2+) in tanycytes via ATP released through connexin 43 hemichannels. Glia 2012, 60, 53–68. [Google Scholar] [CrossRef] [Green Version]
- Sanders, N.M.; Dunn-Meynell, A.A.; Levin, B.E. Third ventricular alloxan reversibly impairs glucose counterregulatory responses. Diabetes 2004, 53, 1230–1236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakano, Y.; Furube, E.; Morita, S.; Wanaka, A.; Nakashima, T.; Miyata, S. Astrocytic TLR4 expression and LPS-induced nuclear translocation of STAT3 in the sensory circumventricular organs of adult mouse brain. J. Neuroimmunol. 2015, 278, 144–158. [Google Scholar] [CrossRef] [PubMed]
- Clayton, B.L.L.; Tesar, P.J. Oligodendrocyte progenitor cell fate and function in development and disease. Curr. Opin. Cell Biol. 2021, 73, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Djogo, T.; Robins, S.C.; Schneider, S.; Kryzskaya, D.; Liu, X.; Mingay, A.; Gillon, C.J.; Kim, J.H.; Storch, K.F.; Boehm, U.; et al. Adult NG2-glia are required for median eminence-mediated leptin sensing and body weight control. Cell Metab. 2016, 23, 797–810. [Google Scholar] [CrossRef]
- Ou, Z.; Ma, Y.; Sun, Y.; Zheng, G.; Wang, S.; Xing, R.; Chen, X.; Han, Y.; Wang, J.; Lu, Q.R.; et al. A GPR17-cAMP-lactate signaling axis in oligodendrocytes regulates whole-body metabolism. Cell Rep. 2019, 26, 2984–2997.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohnke, S.; Buller, S.; Nuzzaci, D.; Ridley, K.; Lam, B.; Pivonkova, H.; Bentsen, M.A.; Alonge, K.M.; Zhao, C.; Tadross, J.; et al. Nutritional regulation of oligodendrocyte differentiation regulates perineuronal net remodeling in the median eminence. Cell Rep. 2021, 36, 109362. [Google Scholar] [CrossRef]
- Alonge, K.M.; Mirzadeh, Z.; Scarlett, J.M.; Logsdon, A.F.; Brown, J.M.; Cabrales, E.; Chan, C.K.; Kaiyala, K.J.; Bentsen, M.A.; Banks, W.A.; et al. Hypothalamic perineuronal net assembly is required for sustained diabetes remission induced by fibroblast growth factor 1 in rats. Nat. Metab. 2020, 2, 1025–1033. [Google Scholar] [CrossRef] [PubMed]
- Levine, J.M.; Enquist, L.W.; Card, J.P. Reactions of oligodendrocyte precursor cells to alpha herpesvirus infection of the central nervous system. Glia 1998, 23, 316–328. [Google Scholar] [CrossRef]
- Holst, J.J.; Rosenkilde, M.M. GIP as a therapeutic target in diabetes and obesity: Insight from incretin co-agonists. J. Clin. Endocrinol. Metab. 2020, 105, e2710–e2716. [Google Scholar] [CrossRef] [PubMed]
- Schur, E.A.; Melhorn, S.J.; Oh, S.K.; Lacy, J.M.; Berkseth, K.E.; Guyenet, S.J.; Sonnen, J.A.; Tyagi, V.; Rosalynn, M.; De Leon, B.; et al. Radiologic evidence that hypothalamic gliosis is associated with obesity and insulin resistance in humans. Obesity 2015, 23, 2142–2148. [Google Scholar] [CrossRef]
- Van de Sande-Lee, S.; Melhorn, S.J.; Rachid, B.; Rodovalho, S.; De-Lima-Junior, J.C.; Campos, B.M.; Pedro, T.; Beltramini, G.C.; Chaim, E.A.; Pareja, J.C.; et al. Radiologic evidence that hypothalamic gliosis is improved after bariatric surgery in obese women with type 2 diabetes. Int. J. Obes. 2020, 44, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Rosenbaum, J.L.; Melhorn, S.J.; Schoen, S.; Webb, M.F.; De Leon, M.R.B.; Humphreys, M.; Utzschneider, K.M.; Schur, E.A. Evidence That Hypothalamic Gliosis Is Related to Impaired Glucose Homeostasis in Adults With Obesity. Diabetes Care 2021, dc211535. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Troadec, J.-D.; Gaigé, S.; Barbot, M.; Lebrun, B.; Barbouche, R.; Abysique, A. Glial Modulation of Energy Balance: The Dorsal Vagal Complex Is No Exception. Int. J. Mol. Sci. 2022, 23, 960. https://doi.org/10.3390/ijms23020960
Troadec J-D, Gaigé S, Barbot M, Lebrun B, Barbouche R, Abysique A. Glial Modulation of Energy Balance: The Dorsal Vagal Complex Is No Exception. International Journal of Molecular Sciences. 2022; 23(2):960. https://doi.org/10.3390/ijms23020960
Chicago/Turabian StyleTroadec, Jean-Denis, Stéphanie Gaigé, Manon Barbot, Bruno Lebrun, Rym Barbouche, and Anne Abysique. 2022. "Glial Modulation of Energy Balance: The Dorsal Vagal Complex Is No Exception" International Journal of Molecular Sciences 23, no. 2: 960. https://doi.org/10.3390/ijms23020960
APA StyleTroadec, J. -D., Gaigé, S., Barbot, M., Lebrun, B., Barbouche, R., & Abysique, A. (2022). Glial Modulation of Energy Balance: The Dorsal Vagal Complex Is No Exception. International Journal of Molecular Sciences, 23(2), 960. https://doi.org/10.3390/ijms23020960