



Oxidative Stress Amelioration of Novel Peptides Extracted from Enzymatic Hydrolysates of Chinese Pecan Cake

 , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Separation of Antioxidant Peptides
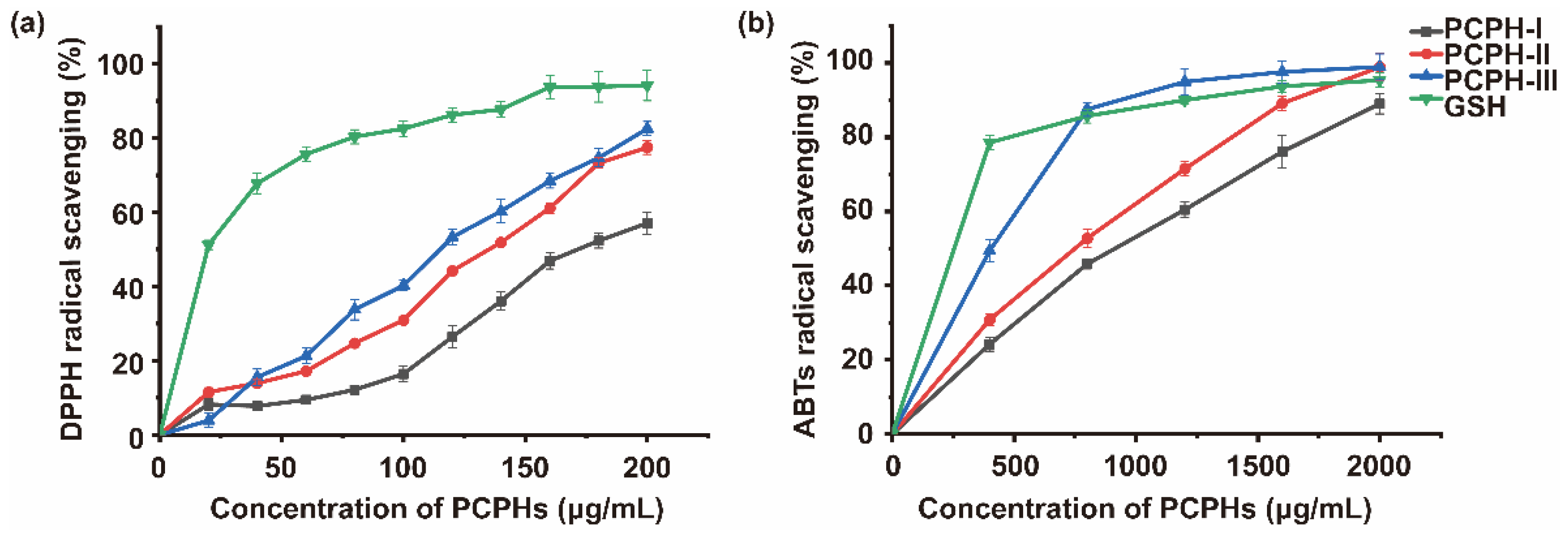
2.1.1. Antioxidant Capacity of Membrane Ultrafiltration Fractions
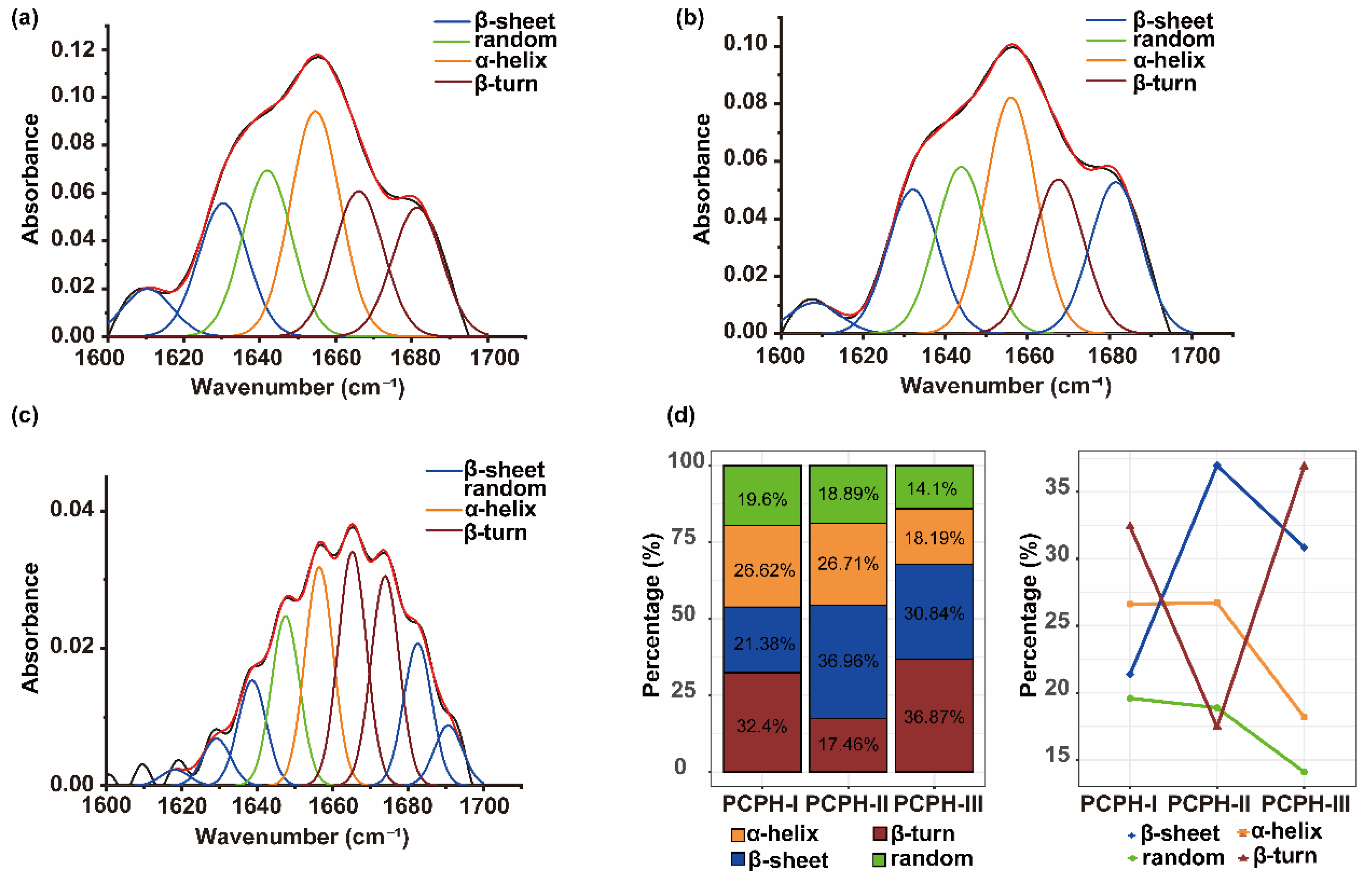
2.1.2. Analysis of Ultrafiltration Components by FTIR Spectroscopy and Amino Acid Composition
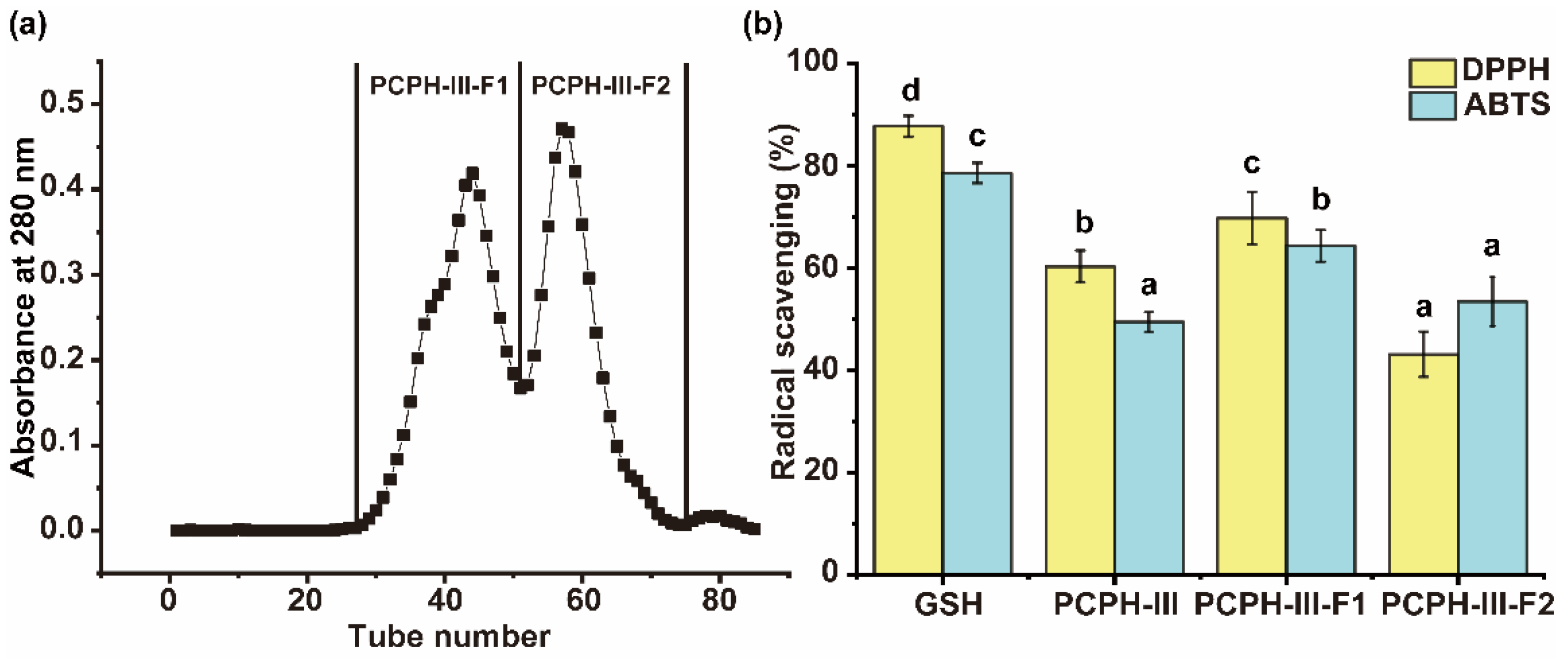
2.1.3. Antioxidant Capacity of gel Chromatography Fractions from PCPH-III
2.2. Identification of Antioxidant Peptides by LC-MS/MS
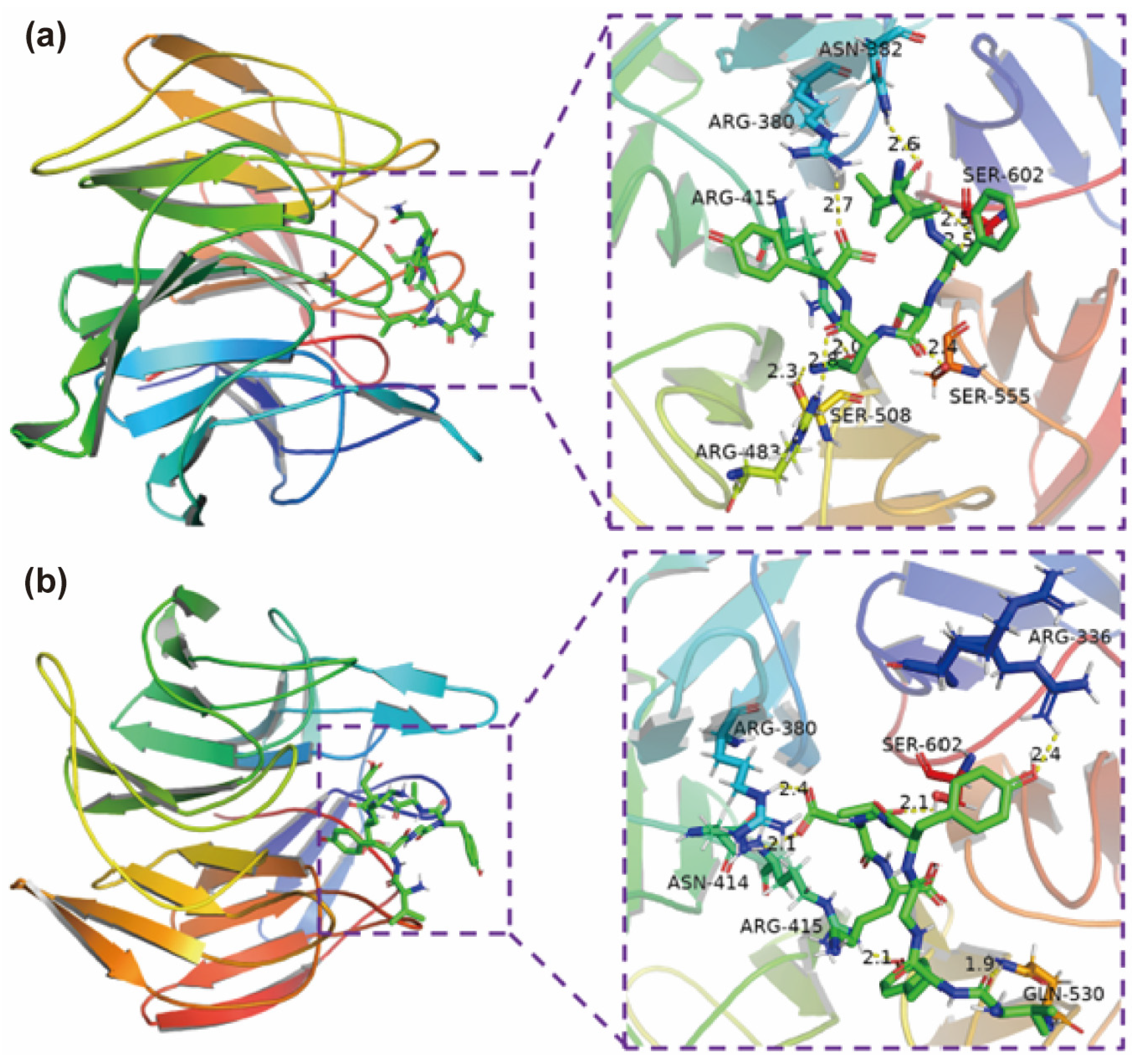
2.3. Molecular Docking for Potential Antioxidant Mechanisms
2.4. Effects of Peptides on Oxidative Stress Induced by H2O2
2.5. Determination of the Crucial Antioxidant Residues of Antioxidant Peptides
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Preparation of Pecan Cake Protein Hydrolysates (PCPHs)
4.3. Separation and Analysis of Pecan Cake Peptides
4.3.1. Ultrafiltration
4.3.2. Gel Filtration Chromatography
4.3.3. Analysis of Amino Acid Composition
4.4. Antioxidant Capacity
4.4.1. 1,1-Diphenyl-2-Picrylhydrazyl (DPPH) Free Radical Scavenging Capacity
4.4.2. 2,2′-Azino-bis (3-Ethylbenzothiazoline-6-Sulfonic Acid) Diammonium Salt (ABTS) Radical Scavenging Capacity
4.5. Fourier Transform Infrared Spectroscopy (FTIR) Determination of Secondary Structure
4.6. Identification of Peptide by Nano LC-MS/MS
4.7. Peptide Synthesis
4.8. Molecular Docking
4.9. Measurement of ROS Content, SOD and CAT Activities
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sies, H. Oxidative stress: A concept in redox biology and medicine. Redox. Biol. 2015, 4, 180–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, F.; Sono, Y.; Ito, T. Measurement and Clinical Significance of Lipid Peroxidation as a Biomarker of Oxidative Stress: Oxidative Stress in Diabetes, Atherosclerosis, and Chronic Inflammation. Antioxidants 2019, 8, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; et al. Oxidative stress, aging, and diseases. Clin. Interv. Aging 2018, 13, 752–772. [Google Scholar] [CrossRef] [Green Version]
- Izzo, C.; Vitillo, P.; Di Pietro, P.; Visco, V.; Strianese, A.; Virtuoso, N.; Ciccarelli, M.; Galasso, G.; Carrizzo, A.; Vecchione, C. The Role of Oxidative Stress in Cardiovascular Aging and Cardiovascular Diseases. Life 2021, 11, 60. [Google Scholar] [CrossRef] [PubMed]
- Bobadilla, M.; Garcia-Sanmartin, J.; Martinez, A. Natural Food Supplements Reduce Oxidative Stress in Primary Neurons and in the Mouse Brain, Suggesting Applications in the Prevention of Neurodegenerative Diseases. Antioxidants 2021, 10, 46. [Google Scholar] [CrossRef] [PubMed]
- Forman, H.J.; Zhang, H. Targeting oxidative stress in disease: Promise and limitations of antioxidant therapy. Nat. Rev. Drug Discov. 2021, 20, 689–709. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.S.; Choi, Y.H.; Choi, Y.S.; Alam, M.B.; Lee, S.H.; Yoo, J.C. A novel antioxidant peptide, purified from Bacillus amyloliquefaciens, showed strong antioxidant potential via Nrf-2 mediated heme oxygenase-1 expression. Food Chem. 2018, 239, 502–510. [Google Scholar] [CrossRef] [PubMed]
- Toldra, F.; Reig, M.; Aristoy, M.C.; Mora, L. Generation of bioactive peptides during food processing. Food Chem. 2018, 267, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Sheng, J.; Yang, X.; Chen, J.; Peng, T.; Yin, X.; Liu, W.; Liang, M.; Wan, J.; Yang, X. Antioxidative Effects and Mechanism Study of Bioactive Peptides from Defatted Walnut (Juglans regia L.) Meal Hydrolysate. J. Agr. Food Chem. 2019, 67, 3305–3312. [Google Scholar] [CrossRef]
- Yang, J.; Huang, J.; Dong, X.; Zhang, Y.; Zhou, X.; Huang, M.; Zhou, G. Purification and identification of antioxidant peptides from duck plasma proteins. Food Chem. 2020, 319, 126534. [Google Scholar] [CrossRef]
- Chai, T.T.; Xiao, J.; Mohana Dass, S.; Teoh, J.Y.; Ee, K.Y.; Ng, W.J.; Wong, F.C. Identification of antioxidant peptides derived from tropical jackfruit seed and investigation of the stability profiles. Food Chem. 2021, 340, 127876. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, M.; Zhang, G.; Tian, Y.; Kong, F.; Xiong, S.; Zhao, S.; Jia, D.; Manyande, A.; Du, H. Identification of novel antioxidant peptides from snakehead (Channa argus) soup generated during gastrointestinal digestion and insights into the anti-oxidation mechanisms. Food Chem. 2021, 337, 127921. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Ma, M.; Yu, Z.; Du, S.K. Preparation and identification of antioxidant peptides from cottonseed proteins. Food Chem. 2021, 352, 129399. [Google Scholar] [CrossRef]
- Orsini Delgado, M.C.; Nardo, A.; Pavlovic, M.; Rogniaux, H.; Anon, M.C.; Tironi, V.A. Identification and characterization of antioxidant peptides obtained by gastrointestinal digestion of amaranth proteins. Food Chem. 2016, 197, 1160–1167. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Tong, X.; Sui, X.; Wang, Z.; Qi, B.; Li, Y.; Jiang, L. Antioxidant activity and protective effects of Alcalase-hydrolyzed soybean hydrolysate in human intestinal epithelial Caco-2 cells. Food Res. Int. 2018, 111, 256–264. [Google Scholar] [CrossRef]
- Etemadian, Y.; Ghaemi, V.; Shaviklo, A.R.; Pourashouri, P.; Mahoonak, A.R.S.; Rafipour, F. Development of animal/plant-based protein hydrolysate and its application in food, feed and nutraceutical industries: State of the art. J. Clean. Prod. 2021, 278, 123219. [Google Scholar] [CrossRef]
- Maciel, L.G.; Ribeiro, F.L.; Teixeira, G.L.; Molognoni, L.; dos Santos, J.N.; Nunes, I.L.; Block, J.M. The potential of the pecan nut cake as an ingredient for the food industry. Food Res. Int. 2020, 127, 108717. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; He, Z.; Wang, Q.; Wu, F.; Liu, X. Response Surface Optimization of Enzymatic Hydrolysis of Peptides of Chinese Pecan (Carya cathayensis) and Analysis of Their Antioxidant Capacities and Structures. Int. J. Pept. Res. Ther. 2021, 27, 1239–1251. [Google Scholar] [CrossRef]
- Tonolo, F.; Folda, A.; Cesaro, L.; Scalcon, V.; Marin, O.; Ferro, S.; Bindoli, A. Milk-derived bioactive peptides exhibit antioxidant activity through the Keap1-Nrf2 signaling pathway. J. Funct. Foods 2020, 64, 103696. [Google Scholar] [CrossRef]
- Wen, C.; Zhang, J.; Zhang, H.; Duan, Y.; Ma, H. Plant protein-derived antioxidant peptides: Isolation, identification, mechanism of action and application in food systems: A review. Trends Food Sci. Technol. 2020, 105, 308–322. [Google Scholar] [CrossRef]
- Yang, R.; Hui, Q.; Jiang, Q.; Liu, S.; Zhang, H.; Wu, J.; Lin, F.; Karmin, O.; Yang, C. Effect of Manitoba-Grown Red-Osier Dogwood Extracts on Recovering Caco-2 Cells from H2O2-Induced Oxidative Damage. Antioxidants 2019, 8, 250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khor, B.-H.; Tallman, D.A.; Karupaiah, T.; Khosla, P.; Chan, M.; Kopple, J.D. Nutritional Adequacy of Animal-Based and Plant-Based Asian Diets for Chronic Kidney Disease Patients: A Modeling Study. Nutrients 2021, 13, 3341. [Google Scholar] [CrossRef] [PubMed]
- Lassoued, I.; Mora, L.; Nasri, R.; Jridi, M.; Toldra, F.; Aristoy, M.-C.; Barkia, A.; Nasri, M. Characterization and comparative assessment of antioxidant and ACE inhibitory activities of thornback ray gelatin hydrolysates. J. Funct. Foods 2015, 13, 225–238. [Google Scholar] [CrossRef]
- Samaranayaka, A.G.P.; Li-Chan, E.C.Y. Food-derived peptidic antioxidants: A review of their production, assessment, and potential applications. J. Funct. Foods 2011, 3, 229–254. [Google Scholar] [CrossRef]
- Zhang, J.; Wen, C.; Li, C.; Duan, Y.; Zhang, H.; Ma, H. Antioxidant Peptide Fractions Isolated from Wheat Germ Protein with Subcritical Water Extraction and Its Transport Across Caco-2 Cells. J. Food Sci. 2019, 84, 2139–2146. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Huang, X.; Tu, F.; Wang, C.; Yang, F. Preparation, antioxidant activity evaluation, and identification of antioxidant peptide from black soldier fly (Hermetia illucens L.) larvae. J. Food Biochem. 2020, 44, e13186. [Google Scholar] [CrossRef]
- Bongirwar, V.; Mokhade, A.S. Different methods, techniques and their limitations in protein structure prediction: A review. Prog. Biophys. Mol. Biol. 2022, 173, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Banc, A.; Desbat, B.; Cavagnat, D. Ab Initio Calculations of Proline Vibrations with and without Water: Consequences on the Infrared Spectra of Proline-Rich Proteins. Appl. Spectrosc. 2011, 65, 817–819. [Google Scholar] [CrossRef]
- Ghumman, A.; Singh, N.; Kaur, A.; Rana, J.C. Diversity in protein secondary structure, molecular weight, mineral and amino acid composition of lentil and horse gram germplasm. J. Food Sci. Technol. 2019, 56, 1601–1612. [Google Scholar] [CrossRef]
- MacArthur, M.W.; Thornton, J.M. Influence of proline residues on protein conformation. J. Mol. Biol. 1991, 218, 397–412. [Google Scholar] [CrossRef]
- Akita, M.; Nishikawa, Y.; Shigenobu, Y.; Ambe, D.; Morita, T.; Morioka, K.; Adachi, K. Correlation of proline, hydroxyproline and serine content, denaturation temperature and circular dichroism analysis of type I collagen with the physiological temperature of marine teleosts. Food Chem. 2020, 329, 126775. [Google Scholar] [CrossRef] [PubMed]
- Sila, A.; Bougatef, A. Antioxidant peptides from marine by-products: Isolation, identification and application in food systems. A review. J. Funct. Foods 2016, 21, 10–26. [Google Scholar] [CrossRef]
- Zheng, Q.; Qiu, D.; Liu, X.; Zhang, L.; Cai, S.; Zhang, X. Antiproliferative effect of Dendrobium catenatum Lindley polypeptides against human liver, gastric and breast cancer cell lines. Food Funct. 2015, 6, 1489–1495. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Sun, J.; Ding, S.; Qi, B. Isolation and identification of novel antioxidant and antimicrobial oligopeptides from enzymatically hydrolyzed anchovy fish meal. Process Biochem. 2018, 74, 148–155. [Google Scholar] [CrossRef]
- Ahmed, A.S.; El-Bassiony, T.; Elmalt, L.M.; Ibrahim, H.R. Identification of potent antioxidant bioactive peptides from goat milk proteins. Food Res. Int. 2015, 74, 80–88. [Google Scholar] [CrossRef]
- Puchalska, P.; Luisa Marina, M.; Concepcion Garcia, M. Isolation and identification of antioxidant peptides from commercial soybean-based infant formulas. Food Chem. 2014, 148, 147–154. [Google Scholar] [CrossRef]
- Bamdad, F.; Ahmed, S.; Chen, L. Specifically designed peptide structures effectively suppressed oxidative reactions in chemical and cellular systems. J. Funct. Foods 2015, 18, 35–46. [Google Scholar] [CrossRef]
- Wang, L.; Ding, L.; Yu, Z.; Zhang, T.; Ma, S.; Liu, J. Intracellular ROS scavenging and antioxidant enzyme regulating capacities of corn gluten meal-derived antioxidant peptides in HepG2 cells. Food Res. Int. 2016, 90, 33–41. [Google Scholar] [CrossRef]
- Wen, C.; Zhang, J.; Zhang, H.; Duan, Y.; Ma, H. Effects of divergent ultrasound pretreatment on the structure of watermelon seed protein and the antioxidant activity of its hydrolysates. Food Chem. 2019, 299, 125165. [Google Scholar] [CrossRef]
- Yu, Z.; Kang, L.; Zhao, W.; Wu, S.; Ding, L.; Zheng, F.; Liu, J.; Li, J. Identification of novel umami peptides from myosin via homology modeling and molecular docking. Food Chem. 2021, 344, 128728. [Google Scholar] [CrossRef]
- Magesh, S.; Chen, Y.; Hu, L. Small molecule modulators of Keap1-Nrf2-ARE pathway as potential preventive and therapeutic agents. Med. Res. Rev. 2012, 32, 687–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo, S.-C.; Li, X.; Henzl, M.T.; Beamer, L.J.; Hannink, M. Structure of the Keap1:Nrf2 interface provides mechanistic insight into Nrf2 signaling. EMBO J. 2006, 25, 3605–3617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, W.; Lin, G.; Zhang, R.; Liang, Z.; Wu, W. Studies on the bioactivities and molecular mechanism of antioxidant peptides by 3D-QSAR, In Vitro evaluation and molecular dynamic simulations. Food Funct. 2020, 11, 3043–3052. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Liu, J.; Nie, S.; Ding, L.; Wang, L.; Liu, J.; Liu, W.Z.; Zhang, T. Direct inhibition of Keap1–Nrf2 interaction by egg-derived peptides DKK and DDW revealed by molecular docking and fluorescence polarization. RSC Adv. 2017, 7, 34963–34971. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Cai, X.; Yan, A.; Tian, Y.; Du, M.; Wang, S. A specific antioxidant peptide: Its properties in controlling oxidation and possible action mechanism. Food Chem. 2020, 327, 126984. [Google Scholar] [CrossRef]
- Tyler, M.I. Amino acid analysis. An overview. Methods Mol. Biol. 2000, 159, 1–7. [Google Scholar]
- Gaspari, M.; Cuda, G. Nano LC-MS/MS: A robust setup for proteomic analysis. Methods Mol. Biol. 2011, 790, 115–126. [Google Scholar]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide Sequence | Protein Source | MW (Da) | Intensity | DPPH (%) | ABTS (%) |
|---|---|---|---|---|---|
| FYSLHTF | Cytochrome b6 | 913.43 | 4.17 × 108 | 12.40 ± 3.07 a,b | 13.83 ± 0.41 b |
| VLFSNY | Photosystem II reaction center protein L | 741.37 | 2.17 × 108 | 17.50 ± 3.36 b | 26.95 ± 2.45 d |
| SSGHTLPAGV | E3 ubiquitin protein ligase RIN2 | 966.48 | 3.78 × 108 | 18.90 ± 3.39 b | 4.47 ± 0.61 a |
| VYGYADK | Protein TIC 214 | 814.39 | 2.11 × 108 | 32.93 ± 5.21 c | 22.62 ± 0.82 c,d |
| TFQGPPHG | Ribulose bisphosphate carboxylase large chain | 839.4 | 1.56 × 107 | 6.71 ± 1.61 a | 3.24 ± 0.31 a |
| YTPEYQTK | Ribulose bisphosphate carboxylase large chain | 1028.48 | 2.39 × 107 | 11.38 ± 2.82 a,b | 21.33 ± 4.28 c |
| Ligand Sequence | Interaction Energy (kcal/mol) | Number of Hydrogen Bonds | Binding Amino Acid Residues of Keap1 |
|---|---|---|---|
| FYSLHTF | −8.5 | 9 | ARG380, ASN414, ARG415, SER431, GLN530, SER602 |
| VLFSNY | −8.7 | 8 | ARG380, ASN382, ARG415, ARG483, SER508, SER555, SER602 |
| SSGHTLPAGV | −7.5 | 9 | ARG380, ARG415, ARG483, TYR525, GLN530, SER555 |
| VYGYADK | −8.8 | 6 | ARG336, ARG380, ASN414, ARG415, GLN530, SER602 |
| TFQGPPHG | −7.9 | 11 | ARG380, ASN382, ASN414, ARG415, SER431, GLY433, SER555, SER602 |
| YTPEYQTK | −6.8 | 14 | TYR334, SER363, ARG380, ASN382, ARG415, SER431, HIS436, ARG483, TYR525, GLN530, SER555, SER602 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Wu, S.; Wang, Q.; Yuan, Q.; Li, Y.; Reboredo-Rodríguez, P.; Varela-López, A.; He, Z.; Wu, F.; Hu, H.; et al. Oxidative Stress Amelioration of Novel Peptides Extracted from Enzymatic Hydrolysates of Chinese Pecan Cake. Int. J. Mol. Sci. 2022, 23, 12086. https://doi.org/10.3390/ijms232012086
Zhang J, Wu S, Wang Q, Yuan Q, Li Y, Reboredo-Rodríguez P, Varela-López A, He Z, Wu F, Hu H, et al. Oxidative Stress Amelioration of Novel Peptides Extracted from Enzymatic Hydrolysates of Chinese Pecan Cake. International Journal of Molecular Sciences. 2022; 23(20):12086. https://doi.org/10.3390/ijms232012086
Chicago/Turabian StyleZhang, Jiaojiao, Shaozhen Wu, Qingqing Wang, Qinjie Yuan, Yane Li, Patricia Reboredo-Rodríguez, Alfonso Varela-López, Zhiping He, Fenghua Wu, Hao Hu, and et al. 2022. "Oxidative Stress Amelioration of Novel Peptides Extracted from Enzymatic Hydrolysates of Chinese Pecan Cake" International Journal of Molecular Sciences 23, no. 20: 12086. https://doi.org/10.3390/ijms232012086
APA StyleZhang, J., Wu, S., Wang, Q., Yuan, Q., Li, Y., Reboredo-Rodríguez, P., Varela-López, A., He, Z., Wu, F., Hu, H., & Liu, X. (2022). Oxidative Stress Amelioration of Novel Peptides Extracted from Enzymatic Hydrolysates of Chinese Pecan Cake. International Journal of Molecular Sciences, 23(20), 12086. https://doi.org/10.3390/ijms232012086