
Transcriptome Analysis Reveals Genes and Pathways Associated with Salt Tolerance during Seed Germination in Suaeda liaotungensis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Salinity Tolerance of S. liaotungensis during Brown Seed Germination
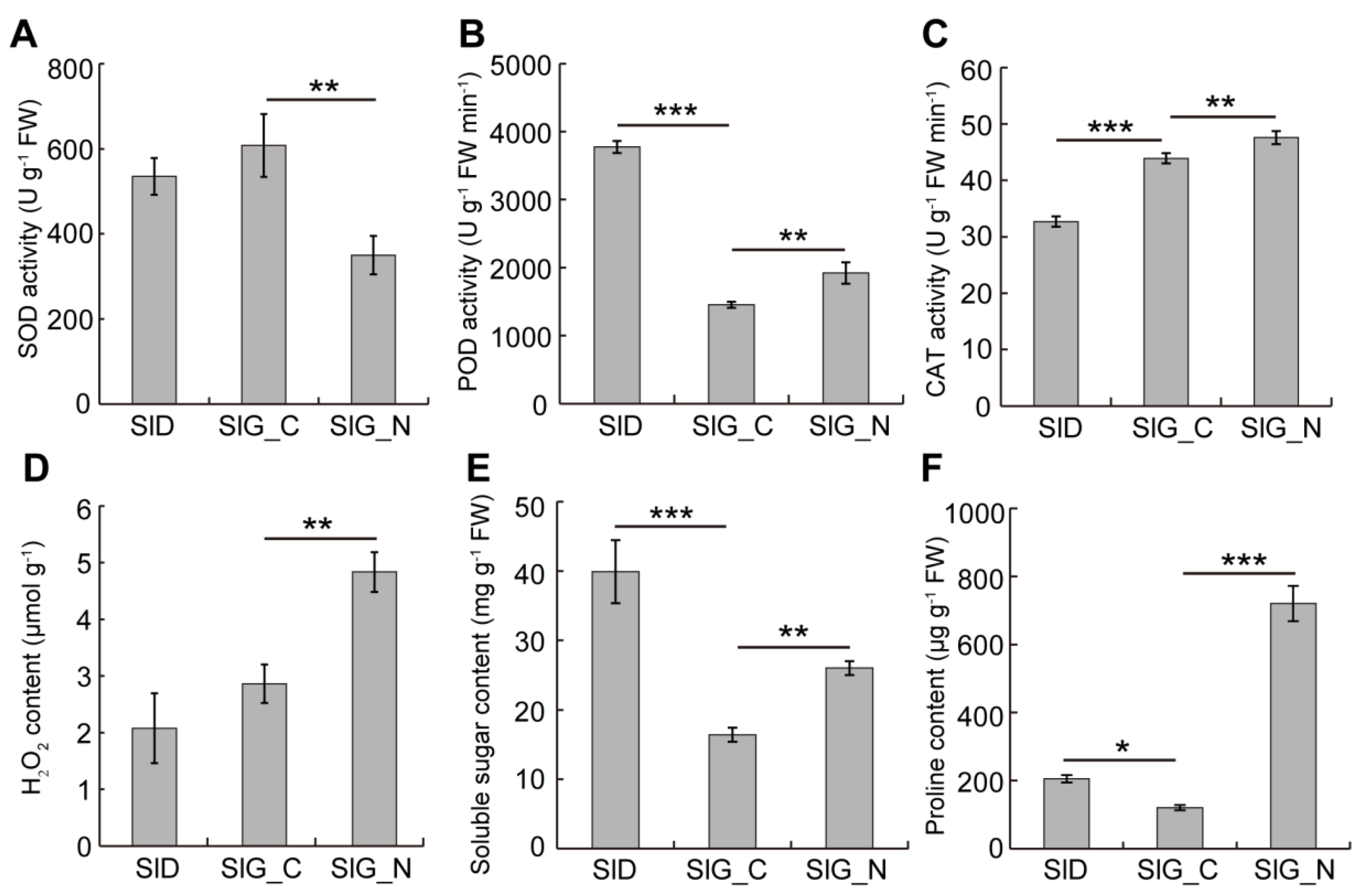
2.2. Determination of Physiological Indexes under Salt Stress
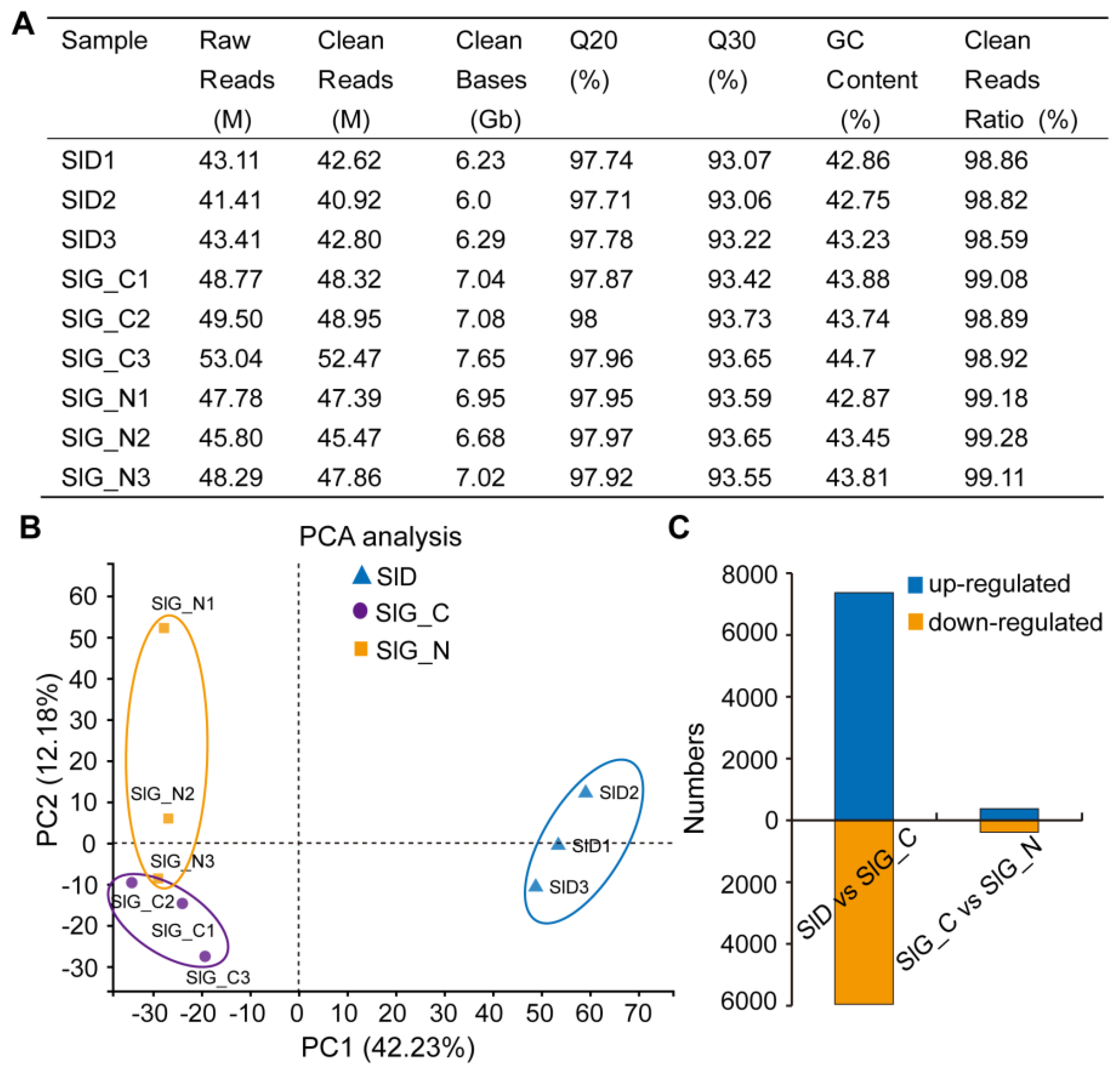
2.3. Sequencing, De Novo Transcriptome Assembly and Annotation
2.4. Identification of Differentially Expressed Genes (DEGs)
2.5. GO and KEGG Enrichment Analysis of DEGs
2.6. Identification of DEGs Associated with Transcription Factor and Hormone Signal Transduction
2.7. DEGs Related to Metabolic Pathways
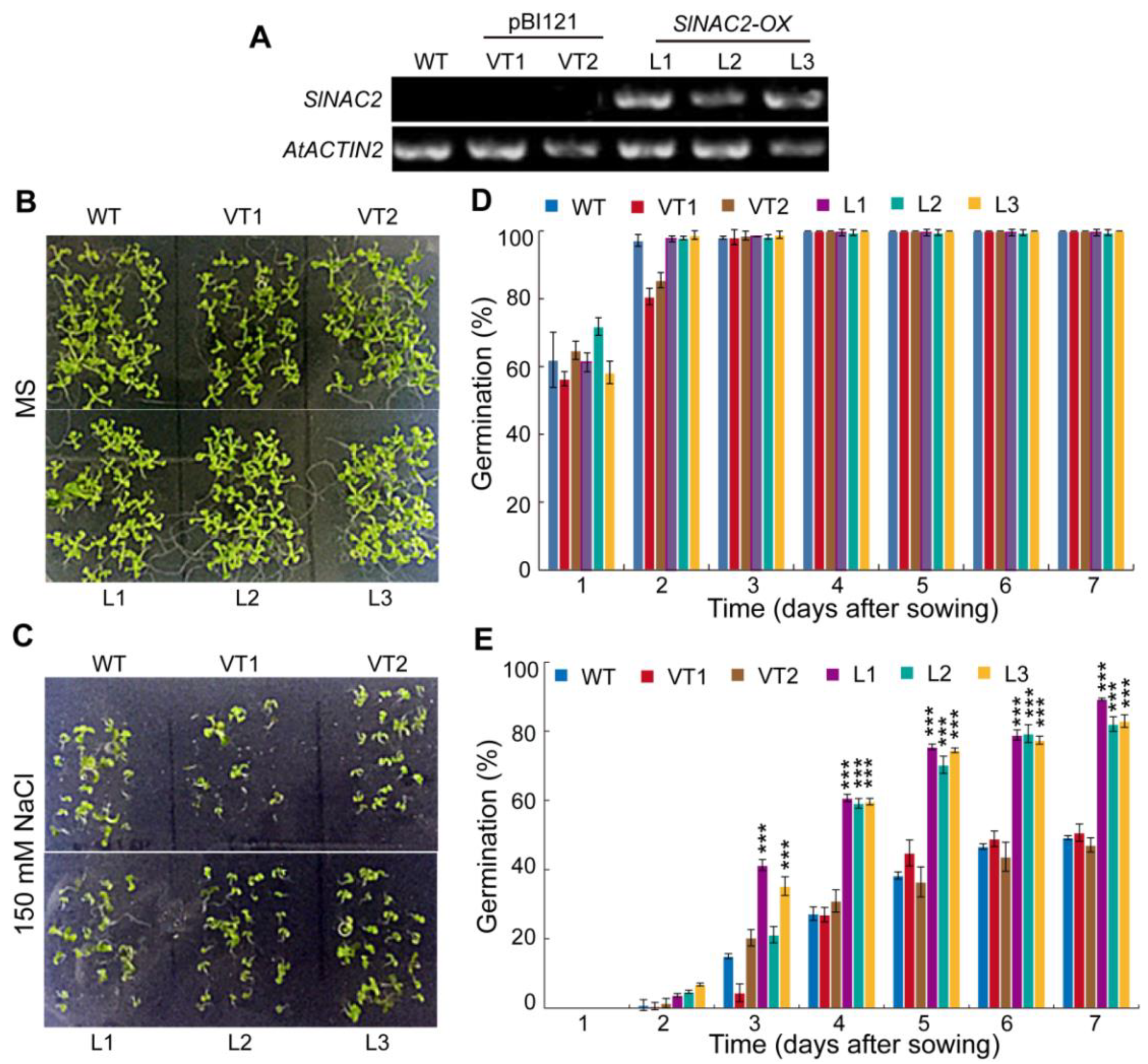
2.8. Verification of DEG by RT-qPCR
2.9. Functional Characterization of SlNAC2 in Seed Germination under Salt Stress in Arabidopsis
3. Discussion
3.1. Plant Hormone Signal Transduction Involved in Seed Germination under Salt Stress
3.2. Transcription Factors Involved in Seed Germination under Salt Stress
3.3. ROS Regulation during Seed Germination under Salt Stress
3.4. Cell Wall Remodeling during Seed Germination under Salt Stress
3.5. Carbohydrate Metabolic Pathway Involved in Seed Germination under Salt Stress
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Seed Germination Assay
4.3. Imbibition Tests
4.4. Determination of Physiological Indexes
4.5. Sample Collection for the Transcriptome Sequencing Analysis
4.6. Library Construction, Sequencing and Transcriptome De Novo Assembly
4.7. Analysis of Differentially Expression Genes (DEGs) and Functional Enrichment
4.8. RT-PCR and Reverse Transcription Quantitative PCR (RT-qPCR)
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.X.; Vinocur, B.; Altman, A. Plant responses to drought, salinity and extreme temperatures: Towards genetic engineering for stress tolerance. Planta 2003, 218, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Rajjou, L.; Duval, M.; Gallardo, K.; Catusse, J.; Bally, J.; Job, C.; Job, D. Seed germination and vigor. Annu. Rev. Plant Biol. 2012, 63, 507–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manz, B.; Müller, K.; Kucera, B.; Volke, F.; Leubner-Metzger, G. Water uptake and distribution in germinating tobacco seeds investigated in vivo by nuclear magnetic resonance imaging. Plant Physiol. 2005, 138, 1538–1551. [Google Scholar] [CrossRef] [Green Version]
- Barrôco, R.M.; Poucke, K.V.; Bergervoet, J.H.W.; Veylder, L.D.; Groot, S.P.C.; Inzé, D.; Engler, G. The role of the cell cycle machinery in resumption of postembryonic development. Plant Physiol. 2005, 137, 127–140. [Google Scholar] [CrossRef] [Green Version]
- Penfield, S. Seed dormancy and germination. Curr. Biol. 2017, 27, R874–R878. [Google Scholar] [CrossRef] [Green Version]
- Finch-Savage, W.E.; Leubner-Metzger, G. Seed dormancy and the control of germination. New Phytol. 2006, 171, 501–523. [Google Scholar] [CrossRef]
- Shu, K.; Liu, X.D.; Xie, Q.; He, Z.H. Two faces of one seed: Hormonal regulation of dormancy and germination. Mol. Plant 2016, 9, 34–45. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, E. A Seed priming to alleviate salinity stress in germinating seeds. J. Plant Physiol. 2016, 192, 38–46. [Google Scholar] [CrossRef]
- Sen, A.; Puthur, J.T. Influence of different seed priming techniques on oxidative and antioxidative responses during the germination of Oryza sativa varieties. Physiol. Mol. Biol. Plants 2020, 26, 551–565. [Google Scholar] [CrossRef]
- Zhang, C.H.; Luo, W.C.; Li, Y.; Zhang, X.; Bai, X.T.; Niu, Z.M.; Zhang, X.; Li, Z.J.; Wan, D.S. Transcriptomic analysis of seed germination under salt stress in two desert sister species (Populus euphratica and P. pruinosa). Front. Genet. 2019, 10, 231. [Google Scholar] [CrossRef]
- Zhang, X.; Li, M.; Yang, H.H.; Li, X.X.; Cui, Z.J. Physiological responses of Suaeda glauca and Arabidopsis thaliana in phytoremediation of heavy metals. J. Environ. Manag. 2018, 223, 132–139. [Google Scholar] [CrossRef]
- Wu, D.D.; Sun, Y.H.; Wang, H.F.; Shi, H.; Su, M.X.; Shan, H.Y.; Li, T.T.; Li, Q.L. The SlNAC8 gene of the halophyte Suaeda liaotungensis enhances drought and salt stress tolerance in transgenic Arabidopsis thaliana. Gene 2018, 662, 10–20. [Google Scholar] [CrossRef]
- Li, Q.L.; Lai, L.D.; Yu, X.D.; Wang, J.Y. The SlCMO gene driven by its own promoter enhances salt tolerance of transgenic tomato without affecting growth and yield. Plant Mol. Biol. Rep. 2018, 36, 596–604. [Google Scholar] [CrossRef]
- Yang, X.; Hu, Y.X.; Li, X.L.; Yu, X.D.; Li, Q.L. Molecular characterization and function analysis of SlNAC2 in Suaeda liaotungensis K. Gene 2014, 543, 190–197. [Google Scholar] [CrossRef]
- Zhu, J.K. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol. 2002, 53, 247–273. [Google Scholar] [CrossRef] [Green Version]
- Tuteja, N. Mechanisms of high salinity tolerance in plants. Methods Enzymol. 2007, 428, 419–438. [Google Scholar]
- Jin, H.X.; Dong, D.K.; Yang, Q.H.; Zhu, D.H. Salt-responsive transcriptome profiling of Suaeda glauca via RNA sequencing. PLoS ONE 2016, 11, e0150504. [Google Scholar] [CrossRef]
- Guo, S.M.; Tan, Y.; Chu, H.J.; Sun, M.X.; Xing, J.C. Transcriptome sequencing revealed molecular mechanisms underlying tolerance of Suaeda salsa to saline stress. PLoS ONE 2019, 14, e0219979. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; He, S.C.; Fu, Y.; Yu, R.L.; Gao, X.; Wang, Z.H.; Liu, Z.J.; Guo, Y.; Chen, M.X. Transcriptome analysis reveals key genes in response to salinity stress during seed germination in Setaria italica. Environ. Exp. Bot. 2021, 191, 104604. [Google Scholar] [CrossRef]
- He, Y.Q.; Yang, B.; He, Y.; Zhan, C.F.; Cheng, Y.H.; Zhang, J.H.; Zhang, H.S.; Cheng, J.P.; Wang, Z.F. A quantitative trait locus, qSE3, promotes seed germination and seedling establishment under salinity stress in rice. Plant J. 2019, 97, 1089–1104. [Google Scholar] [CrossRef]
- Hirayama, T.; Shinozaki, K. Perception and transduction of abscisic acid signals: Keys to the function of the versatile plant hormone ABA. Trends Plant Sci. 2007, 12, 343–351. [Google Scholar] [CrossRef]
- Singh, A.; Jha, S.K.; Bagri, J.; Pandey, G.K. ABA inducible rice protein phosphatase 2C confers ABA insensitivity and abiotic stress tolerance in Arabidopsis. PLoS ONE 2015, 10, e0125168. [Google Scholar] [CrossRef] [Green Version]
- Park, S.Y.; Fung, P.; Nishimura, N.; Jensen, D.R.; Fujii, H.; Zhao, Y.; Lumba, S.; Santiago, J.; Rodrigues, A.; Chow, T.F.; et al. Abscisic acid inhibits type 2C protein phosphatases via the PYR/PYL family of START proteins. Science 2009, 324, 1068–1071. [Google Scholar] [CrossRef] [Green Version]
- Steber, C.M.; McCourt, P. A role for brassinosteroids in germination in Arabidopsis. Plant Physiol. 2001, 125, 763–769. [Google Scholar] [CrossRef] [Green Version]
- Krishna, P. Brassinosteroid-mediated stress responses. J. Plant Growth Regul. 2003, 22, 289–297. [Google Scholar] [CrossRef]
- Kagale, S.; Divi, U.K.; Krochko, J.E.; Keller, W.A.; Krishna, P. Brassinosteroid confers tolerance in Arabidopsis thaliana and Brassica napus to a range of abiotic stresses. Planta 2007, 225, 353–364. [Google Scholar] [CrossRef]
- Zhang, M.; Zhao, J.F.; Li, L.; Gao, Y.N.; Zhao, L.L.; Patil, S.B.; Fang, J.J.; Zhang, W.H.; Yang, Y.H.; Li, M.; et al. The Arabidopsis U-box E3 ubiquitin ligase PUB30 negatively regulates salt tolerance by facilitating BRI1 kinase inhibitor 1 (BKI1) degradation. Plant Cell Environ. 2017, 40, 2831–2843. [Google Scholar] [CrossRef]
- Wang, H.Y.; Wang, H.L.; Shao, H.B.; Tang, X.L. Recent advances in utilizing transcription factors to improve plant abiotic stress tolerance by transgenic technology. Front. Plant Sci. 2016, 7, 67. [Google Scholar] [CrossRef] [Green Version]
- Baillo, E.H.; Kimotho, R.N.; Zhang, Z.B.; Xu, P. Transcription factors associated with abiotic and biotic stress tolerance and their potential for crops improvement. Genes 2019, 10, 771. [Google Scholar] [CrossRef] [Green Version]
- Shin, D.J.; Koo, Y.D.; Lee, J.; Lee, H.J.; Baek, D.; Lee, S.; Cheon, C.I.; Kwak, S.S.; Lee, S.Y.; Yun, D.J. Athb-12, a homeobox-leucine zipper domain protein from Arabidopsis thaliana, increases salt tolerance in yeast by regulating sodium exclusion. Biochem. Biophys. Res. Commun. 2004, 323, 534–540. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.G.; Lee, A.K.; Yoon, H.K.; Park, C.M. A membrane-bound NAC transcription factor NTL8 regulates gibberellic acid-mediated salt signaling in Arabidopsis seed germination. Plant J. 2008, 55, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Kim, Y.S.; Kim, S.G.; Jung, J.H.; Woo, J.C.; Park, C.M. Integration of auxin and salt signals by the NAC transcription factor NTM2 during seed germination in Arabidopsis. Plant Physiol. 2011, 156, 537–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailly, C. Active oxygen species and antioxidants in seed biology. Seed Sci. Res. 2004, 14, 93–107. [Google Scholar] [CrossRef]
- Zelm, E.V.; Zhang, Y.X.; Testerink, C. Salt tolerance mechanisms of plants. Annu. Rev. Plant Biol. 2020, 71, 403–433. [Google Scholar] [CrossRef] [Green Version]
- Blokhina, O.; Virolainen, E.; Fagerstedt, K.V. Antioxidants, oxidative damage and oxygen deprivation stress: A review. Ann. Bot. 2003, 91, 179–194. [Google Scholar] [CrossRef] [Green Version]
- Jemmat, A.M.; Ranocha, P.; Ru, A.L.; Neel, M.; Jauneau, A.; Raggi, S.; Ferrari, S.; Burlat, V.; Dunand, C. Coordination of five class III peroxidase-encoding genes for early germination events of Arabidopsis thaliana. Plant Sci. 2020, 298, 110565. [Google Scholar] [CrossRef]
- Shigeto, J.; Tsutsumi, Y. Diverse functions and reactions of class III peroxidases. New Phytol. 2016, 209, 1395–1402. [Google Scholar] [CrossRef] [Green Version]
- Schopfer, P.; Plachy, C.; Frahry, G. Release of reactive oxygen intermediates (superoxide radicals, hydrogen peroxide, and hydroxyl radicals) and peroxidase in germinating radish seeds controlled by light, gibberellin, and abscisic acid. Plant Physiol. 2001, 125, 1591–1602. [Google Scholar] [CrossRef] [Green Version]
- Linkies, A.; Schuster-Sherpa, U.; Tintelnot, S.; Leubner-Metzger, G.; Müller, K. Peroxidases identified in a subtractive cDNA library approach show tissue-specific transcript abundance and enzyme activity during seed germination of Lepidium sativum. J. Exp. Bot. 2010, 61, 491–502. [Google Scholar] [CrossRef] [Green Version]
- Roberts, E.; Kolattukudy, P.E. Molecular cloning, nucleotide sequence, and abscisic acid induction of a suberization-associated highly anionic peroxidase. Mol. Gen. Genom. 1989, 217, 223–232. [Google Scholar] [CrossRef]
- Su, P.; Yan, J.; Li, W.; Wang, L.; Zhao, J.X.; Ma, X.; Li, A.F.; Wang, H.W.; Kong, L.R. A member of wheat class III peroxidase gene family, TaPRX-2A, enhanced the tolerance of salt stress. BMC Plant Biol. 2020, 20, 392. [Google Scholar] [CrossRef]
- Carpita, N.C.; Gibeaut, D.M. Structural models of primary cell walls in flowering plants: Consistency of molecular structure with the physical properties of the walls during growth. Plant J. 1993, 3, 1–30. [Google Scholar] [CrossRef]
- Steinbrecher, T.; Leubner-Metzger, G. Tissue and cellular mechanics of seeds. Curr. Opin. Genet. Dev. 2018, 51, 1–10. [Google Scholar] [CrossRef]
- Rose, J.K.C.; Braam, J.; Fry, S.C.; Nishitani, K. The XTH family of enzymes involved in xyloglucan endotransglucosylation and endohydrolysis: Current perspectives and a new unifying nomenclature. Plant Cell Physiol. 2002, 43, 1421–1435. [Google Scholar] [CrossRef] [Green Version]
- Xu, P.P.; Fang, S.; Chen, H.Y.; Cai, W.M. The brassinosteroid-responsive xyloglucan endotransglucosylase/hydrolase 19 (XTH19) and XTH23 genes are involved in lateral root development under salt stress in Arabidopsis. Plant J. 2020, 104, 59–75. [Google Scholar] [CrossRef]
- Yennawar, N.H.; Li, L.C.; Dudzinski, D.M.; Tabuchi, A.; Cosgrove, D.J. Crystal structure and activities of EXPB1 (Zea m 1), a beta-expansin and group-1 pollen allergen from maize. Proc. Natl. Acad. Sci. USA 2006, 103, 14664–14671. [Google Scholar] [CrossRef] [Green Version]
- Lü, P.; Kang, M.; Jiang, X.Q.; Dai, F.W.; Gao, J.P.; Zhang, C.Q. RhEXPA4, a rose expansin gene, modulates leaf growth and confers drought and salt tolerance to Arabidopsis. Planta 2013, 237, 1547–1559. [Google Scholar] [CrossRef]
- Kim, J.; Shiu, S.H.; Thoma, S.; Li, W.H.; Patterson, S.E. Patterns of expansion and expression divergence in the plant polygalacturonase gene family. Genome Biol. 2006, 7, R87. [Google Scholar]
- Grant, G.T.; Morris, E.R.; Rees, D.A.; Smith, P.J.C.; Thorn, D. Biological interactions between polysaccharides and divalent cations: The egg-box model. FEBS Lett. 1973, 32, 195–198. [Google Scholar] [CrossRef] [Green Version]
- Phan, T.D.; Bo, W.; West, G.; Lycett, G.W.; Tucker, G.A. Silencing of the major salt-dependent isoform of pectinesterase in tomato alters fruit softening. Plant Physiol. 2007, 144, 1960–1967. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.X.; Schopfer, P. Hydroxyl-radical production in physiological reactions. A novel function of peroxidase. Eur. J. Biochem. 1999, 260, 726–735. [Google Scholar] [CrossRef]
- Müller, K.; Linkies, A.; Vreeburg, R.A.M.; Fry, S.C.; Krieger-Liszkay, A.; Leubner-Metzger, G. In vivo cell wall loosening by hydroxyl radicals during cress (Lepidium sativum L.) seed germination and elongation growth. Plant Physiol. 2009, 150, 1855–1865. [Google Scholar] [CrossRef] [Green Version]
- Damaris, R.N.; Lin, Z.Y.; Yang, P.F.; He, D.L. The rice alpha-amylase, conserved regulator of seed maturation and germination. Int. J. Mol. Sci. 2019, 20, 450. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Lin, R.Q.; Jiang, Y.T.; Jiang, S.Z.; Xiong, Y.F.; Lian, H.; Zeng, Q.M.; Liu, X.D.; Liu, Z.J.; Chen, S.Q. Transcriptome analysis and identification of genes associated with starch metabolism in Castanea henryi seed (Fagaceae). Int. J. Mol. Sci. 2020, 21, 431. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.L.; Tian, T.; Chen, J.Z.; Wang, D.; Tong, B.L.; Liu, J. Transcriptome analysis of Cinnamomum migao seed germination in medicinal plants of Southwest China. BMC Plant Biol. 2021, 21, 270. [Google Scholar] [CrossRef]
- Noronha, H.; Silva, A.; Dai, Z.W.; Gallusci, P.; Rombolà, A.D.; Delrot, S.; Gerós, H.A. Molecular perspective on starch metabolism in woody tissues. Planta 2018, 248, 559–568. [Google Scholar] [CrossRef] [Green Version]
- Sonnewald, U.; Lerchl, J.; Zrenner, R.; Frommer, W. Manipulation of source-sink relations in transgenic plants. Plant Cell Environ. 1994, 17, 649–658. [Google Scholar] [CrossRef]
- Fan, W.; Zhang, Z.L.; Zhang, Y.L. Cloning and molecular characterization of fructose-1,6-bisphosphate aldolase gene regulated by high-salinity and drought in Sesuvium portulacastrum. Plant Cell Rep. 2009, 28, 975–984. [Google Scholar] [CrossRef]
- Zhang, X.N.; Lin, C.F.; Chen, H.Y.; Wang, H.; Qu, Z.C.; Zhang, H.W.; Yao, J.H.; Shen, D.L. Cloning of a NaCl-induced fructose-1,6-diphosphate aldolase cDNA from Dualiella salina and its expression in tobacco. Sci. China C Life Sci. 2003, 46, 49–57. [Google Scholar]
- Feng, L.L.; Han, Y.J.; Liu, G.; An, B.G.; Yang, J.; Yang, G.H.; Li, Y.S.; Zhu, Y.G. Overexpression of sedoheptulose-1,7-bisphosphatase enhances photosynthesis and growth under salt stress in transgenic rice plants. Funct. Plant Biol. 2007, 34, 822–834. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, M.; Kiegle, E.; Leo, G.; Ezquer, I. Carbohydrate reserves and seed development: An overview. Plant Reprod. 2018, 31, 263–290. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Khan, H.; Faheem, M.; Zeb, A.; Badshah, M.; Chung, Y.R. Isolation and characterization of an acidic, salt-tolerant endoglucanase Cel5A from a bacterial strain Martelella endophytica YC6887 genome. Mol. Biotechnol. 2021, 63, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Leah, R.; Kigel, J.; Svendsen, I.; Mundy, J. Biochemcial and molecular characterization of a barley seed ß-glucosidase. J. Biol. Chem. 1995, 270, 15789–15797. [Google Scholar] [CrossRef] [Green Version]
- Gallardo, K.; Job, C.; Groot, S.P.C.; Puype, M.; Demol, H.; Vandekerckhove, J.; Job, D. Proteomics of Arabidopsis seed germination. A comparative study of wild-type and gibberellin-deficient seeds. Plant Physiol. 2002, 129, 823–837. [Google Scholar] [CrossRef] [Green Version]
- Kwon, Y.R.; Kim, S.H.; Jung, M.S.; Kim, M.S.; Oh, J.E.; Ju, H.W.; Kim, K.I.; Vierling, E.; Lee, H.; Hong, S.W. Arabidopsis hot2 encodes an endochitinase-like protein that is essential for tolerance to heat, salt and drought stresses. Plant J. 2007, 49, 184–193. [Google Scholar] [CrossRef] [Green Version]
- Baskin, J.M.; Davis, B.H.; Baskin, C.C.; Gleason, S.M.; Cordell, S. Physical dormancy in seeds of Dodonaea viscosa (Sapindales, Sapindaceae) from Hawaii. Seed Sci. Res. 2004, 14, 81–90. [Google Scholar] [CrossRef]
- Wang, Y.G.; Stevanato, P.; Yu, L.H.; Zhao, H.J.; Sun, X.W.; Sun, F.; Li, J.; Geng, G. The physiological and metabolic changes in sugar beet seedlings under different levels of salt stress. J. Plant Res. 2017, 130, 1079–1093. [Google Scholar] [CrossRef]
- Bailey, R.W. Reaction of pentoses with anthrone. Biochem. J. 1958, 68, 669–672. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Xian, A.; Lin, F.; Raychowdhury, R.; Zeng, Q.D.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotech. 2011, 29, 644–652. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, J.; Liu, X.; Li, X.; Wang, H.; Chu, R.; Qu, F.; Zhang, S.; Li, Q. Transcriptome Analysis Reveals Genes and Pathways Associated with Salt Tolerance during Seed Germination in Suaeda liaotungensis. Int. J. Mol. Sci. 2022, 23, 12229. https://doi.org/10.3390/ijms232012229
Song J, Liu X, Li X, Wang H, Chu R, Qu F, Zhang S, Li Q. Transcriptome Analysis Reveals Genes and Pathways Associated with Salt Tolerance during Seed Germination in Suaeda liaotungensis. International Journal of Molecular Sciences. 2022; 23(20):12229. https://doi.org/10.3390/ijms232012229
Chicago/Turabian StyleSong, Jieqiong, Xinjie Liu, Xinxin Li, Hongfei Wang, Ruowen Chu, Fangfang Qu, Sixue Zhang, and Qiuli Li. 2022. "Transcriptome Analysis Reveals Genes and Pathways Associated with Salt Tolerance during Seed Germination in Suaeda liaotungensis" International Journal of Molecular Sciences 23, no. 20: 12229. https://doi.org/10.3390/ijms232012229
APA StyleSong, J., Liu, X., Li, X., Wang, H., Chu, R., Qu, F., Zhang, S., & Li, Q. (2022). Transcriptome Analysis Reveals Genes and Pathways Associated with Salt Tolerance during Seed Germination in Suaeda liaotungensis. International Journal of Molecular Sciences, 23(20), 12229. https://doi.org/10.3390/ijms232012229