
Blockade of Wnt Secretion Attenuates Myocardial Ischemia–Reperfusion Injury by Modulating the Inflammatory Response
Abstract
:1. Introduction
2. Results
2.1. RNAseq Analysis Reveals Altered Gene Expression of Inflammation-Associated Genes upon LGK974 Administration
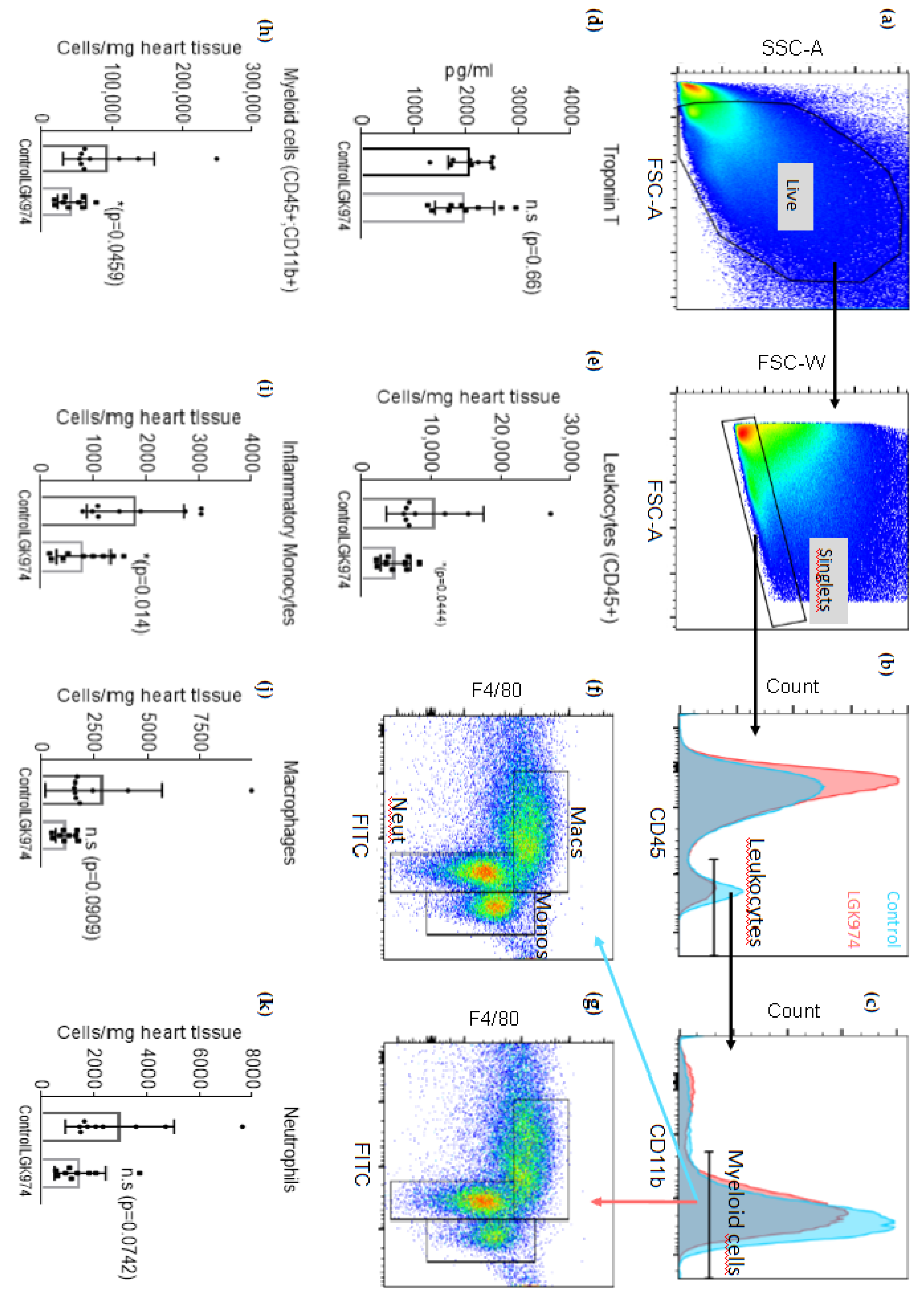
2.2. Blockade of Wnt Protein Secretion by LGK-974 Attenuates the Inflammatory Response after MI
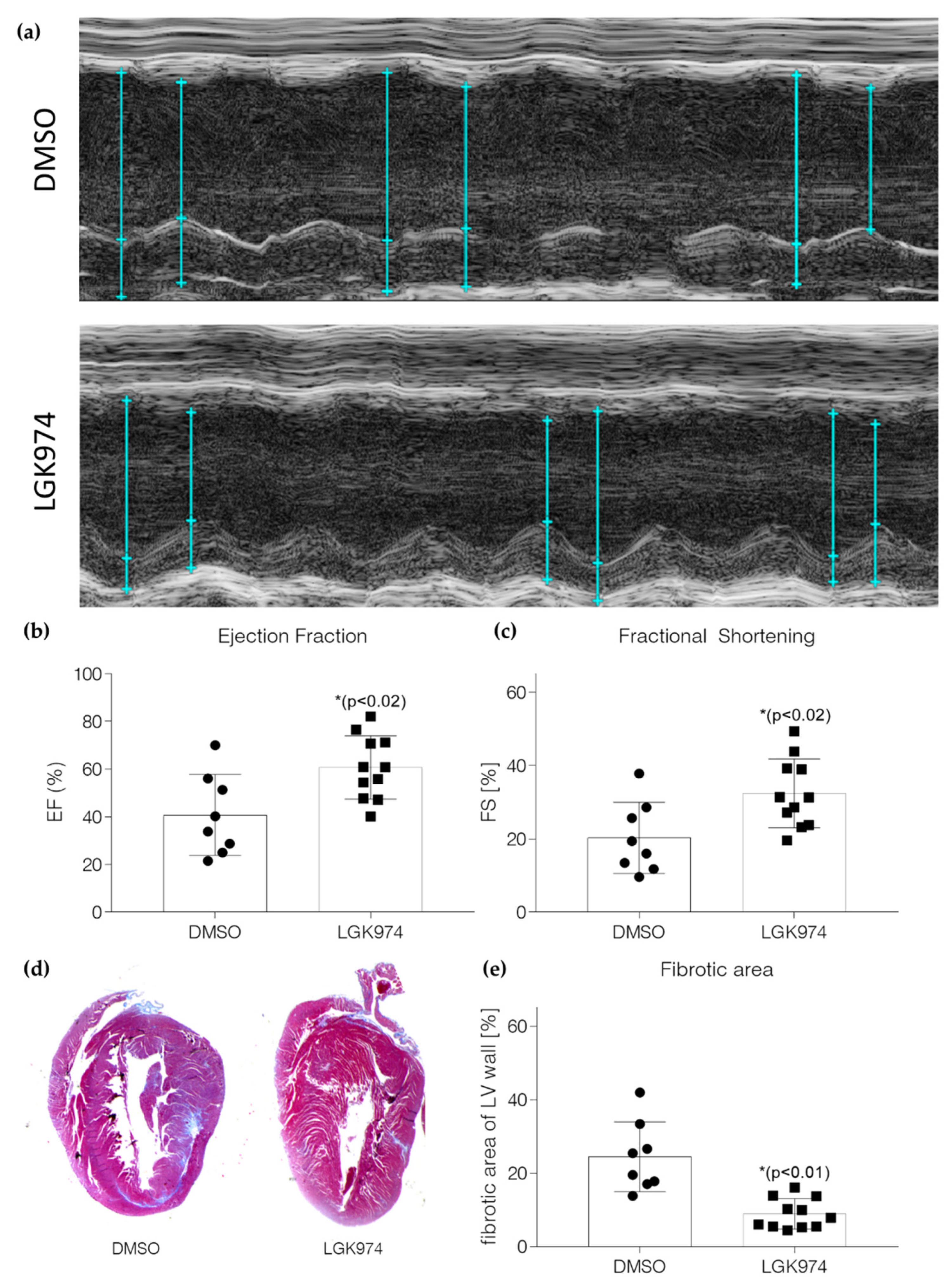
2.3. Blockade of Wnt Secretion by LGK-974 Preserves Heart Function
2.4. Inhibition of Wnt Secretion Inhibits Monocyte/Macrophage Activation In Vitro
3. Discussion
4. Materials and Methods
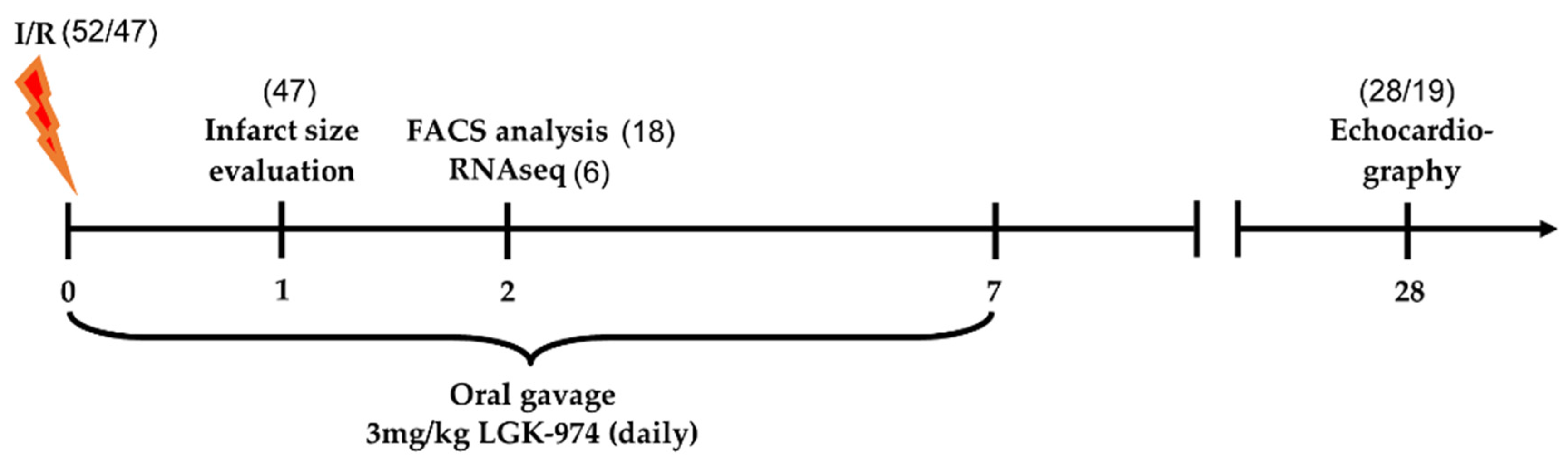
4.1. Induction of Ischemia–Reperfusion Injury and LGK-974 Treatment
4.2. RNA-Sequencing Analyses
4.3. Flow Cytometry Analysis
4.4. Histology
4.5. Ethical Statement
4.6. High-Sensitive Troponin T Determination
4.7. In Vitro Experiments
4.8. Echocardiography
4.9. Quantitative Real-Time PCR
4.10. Western Blotting
4.11. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. World Heart Day 2017; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- Hellermann, J.P.; Jacobsen, S.J.; Gersh, B.J.; Rodeheffer, R.J.; Reeder, G.S.; Roger, V.L. Heart failure after myocardial infarction: A review. Am. J. Med. 2002, 113, 324–330. [Google Scholar] [CrossRef]
- Burchfield, J.S.; Xie, M.; Hill, J.A. Pathological ventricular remodeling: Mechanisms: Part 1 of 2. Circulation 2013, 128, 388–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cahill, T.J.; Kharbanda, R.K. Heart failure after myocardial infarction in the era of primary percutaneous coronary intervention: Mechanisms, incidence and identification of patients at risk. World J. Cardiol. 2017, 9, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Swirski, F.K.; Nahrendorf, M. Leukocyte behavior in atherosclerosis, myocardial infarction, and heart failure. Science 2013, 339, 161–166. [Google Scholar] [CrossRef] [Green Version]
- Saxena, A.; Russo, I.; Frangogiannis, N.G. Inflammation as a therapeutic target in myocardial infarction: Learning from past failures to meet future challenges. Transl. Res. J. Lab. Clin. Med. 2016, 167, 152–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puhl, S.-L.; Steffens, S. Neutrophils in Post-myocardial Infarction Inflammation: Damage vs. Resolution? Front. Cardiovasc. Med. 2019, 6, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nahrendorf, M.; Swirski, F.K.; Aikawa, E.; Stangenberg, L.; Wurdinger, T.; Figueiredo, J.L.; Libby, P.; Weissleder, R.; Pittet, M.J. The healing myocardium sequentially mobilizes two monocyte subsets with divergent and complementary functions. J. Exp. Med. 2007, 204, 3037–3047. [Google Scholar] [CrossRef] [Green Version]
- Hilgendorf, I.; Gerhardt, L.M.; Tan, T.C.; Winter, C.; Holderried, T.A.; Chousterman, B.G.; Iwamoto, Y.; Liao, R.; Zirlik, A.; Scherer-Crosbie, M.; et al. Ly-6Chigh monocytes depend on Nr4a1 to balance both inflammatory and reparative phases in the infarcted myocardium. Circ. Res. 2014, 114, 1611–1622. [Google Scholar] [CrossRef] [Green Version]
- Stylianidis, V.; Hermans, K.C.M.; Blankesteijn, W.M. Wnt Signaling in Cardiac Remodeling and Heart Failure. Handb. Exp. Pharmacol. 2017, 243, 371–393. [Google Scholar] [CrossRef]
- Aisagbonhi, O.; Rai, M.; Ryzhov, S.; Atria, N.; Feoktistov, I.; Hatzopoulos, A.K. Experimental myocardial infarction triggers canonical Wnt signaling and endothelial-to-mesenchymal transition. Dis. Models Mech. 2011, 4, 469–483. [Google Scholar] [CrossRef]
- Niehrs, C. The complex world of WNT receptor signalling. Nat. Rev. Mol. Cell Biol. 2012, 13, 767. [Google Scholar] [CrossRef] [PubMed]
- Meyer, I.S.; Jungmann, A.; Dieterich, C.; Zhang, M.; Lasitschka, F.; Werkmeister, S.; Haas, J.; Müller, O.J.; Boutros, M.; Nahrendorf, M.; et al. The cardiac microenvironment uses non-canonical WNT signaling to activate monocytes after myocardial infarction. EMBO Mol. Med. 2017, 9, 1279–1293. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Pan, S.; Hsieh, M.H.; Ng, N.; Sun, F.; Wang, T.; Kasibhatla, S.; Schuller, A.G.; Li, A.G.; Cheng, D.; et al. Targeting Wnt-driven cancer through the inhibition of Porcupine by LGK974. Proc. Natl. Acad. Sci. USA 2013, 110, 20224–20229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ridker, P.M.; Everett, B.M.; Thuren, T.; MacFadyen, J.G.; Chang, W.H.; Ballantyne, C.; Fonseca, F.; Nicolau, J.; Koenig, W.; Anker, S.D.; et al. Antiinflammatory Therapy with Canakinumab for Atherosclerotic Disease. N. Engl. J. Med. 2017, 377, 1119–1131. [Google Scholar] [CrossRef] [PubMed]
- Ong, S.B.; Hernandez-Resendiz, S.; Crespo-Avilan, G.E.; Mukhametshina, R.T.; Kwek, X.Y.; Cabrera-Fuentes, H.A.; Hausenloy, D.J. Inflammation following acute myocardial infarction: Multiple players, dynamic roles, and novel therapeutic opportunities. Pharmacol. Ther. 2018, 186, 73–87. [Google Scholar] [CrossRef]
- Leuschner, F.; Rauch, P.J.; Ueno, T.; Gorbatov, R.; Marinelli, B.; Lee, W.W.; Dutta, P.; Wei, Y.; Robbins, C.; Iwamoto, Y.; et al. Rapid monocyte kinetics in acute myocardial infarction are sustained by extramedullary monocytopoiesis. J. Exp. Med. 2012, 209, 123–137. [Google Scholar] [CrossRef]
- Frangogiannis, N.G. The inflammatory response in myocardial injury, repair, and remodelling. Nat. Rev. Cardiol. 2014, 11, 255–265. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, K.; Sano, S.; Fuster, J.J.; Kikuchi, R.; Shimizu, I.; Ohshima, K.; Katanasaka, Y.; Ouchi, N.; Walsh, K. Secreted Frizzled-related Protein 5 Diminishes Cardiac Inflammation and Protects the Heart from Ischemia/Reperfusion Injury. J. Biol. Chem. 2016, 291, 2566–2575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laeremans, H.; Rensen, S.S.; Ottenheijm, H.C.; Smits, J.F.; Blankesteijn, W.M. Wnt/frizzled signalling modulates the migration and differentiation of immortalized cardiac fibroblasts. Cardiovasc. Res. 2010, 87, 514–523. [Google Scholar] [CrossRef] [Green Version]
- Moon, J.; Zhou, H.; Zhang, L.-s.; Tan, W.; Liu, Y.; Zhang, S.; Morlock, L.K.; Bao, X.; Palecek, S.P.; Feng, J.Q.; et al. Blockade to pathological remodeling of infarcted heart tissue using a porcupine antagonist. Proc. Natl. Acad. Sci. USA 2017, 114, 1649–1654. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.C.; Kuo, W.-W.; Baskaran, R.; Chen, M.-C.; Ho, T.-J.; Chen, R.-J.; Chen, Y.-F.; Padma, V.V.; Lay, I.-S.; Huang, C.-Y. Enhancement of beta-catenin in cardiomyocytes suppresses survival protein expression but promotes apoptosis and fibrosis. Enhanc. Beta-Catenin Cardiomyocytes Suppresses Surviv. Protein Expr. But Promot. Apoptosis Fibros. 2017, 24, 195–205. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.C.; Chang, R.L.; Chen, Y.F.; Yang, J.J.; Baskaran, R.; Chung, L.C.; Chen, R.J.; Day, C.H.; Vijaya Padma, V.; Huang, C.Y. beta-Catenin overexpression causes an increase in inflammatory cytokines and NF-kappaB activation in cardiomyocytes. Cell. Mol. Biol. 2016, 63, 17–22. [Google Scholar] [CrossRef]
- Assmus, B.; Iwasaki, M.; Schachinger, V.; Roexe, T.; Koyanagi, M.; Iekushi, K.; Xu, Q.; Tonn, T.; Seifried, E.; Liebner, S.; et al. Acute myocardial infarction activates progenitor cells and increases Wnt signalling in the bone marrow. Eur. Heart J. 2012, 33, 1911–1919. [Google Scholar] [CrossRef] [Green Version]
- Barandon, L.; Casassus, F.; Leroux, L.; Moreau, C.; Allieres, C.; Lamaziere, J.M.; Dufourcq, P.; Couffinhal, T.; Duplaa, C. Secreted frizzled-related protein-1 improves postinfarction scar formation through a modulation of inflammatory response. Arterioscler. Thromb. Vasc. Biol. 2011, 31, e80–e87. [Google Scholar] [CrossRef] [Green Version]
- Meyer, I.S.; Leuschner, F. The role of Wnt signaling in the healing myocardium: A focus on cell specificity. Basic Res. Cardiol. 2018, 113, 44. [Google Scholar] [CrossRef]
- Palevski, D.; Levin-Kotler, L.P.; Kain, D.; Naftali-Shani, N.; Landa, N.; Ben-Mordechai, T.; Konfino, T.; Holbova, R.; Molotski, N.; Rosin-Arbesfeld, R.; et al. Loss of Macrophage Wnt Secretion Improves Remodeling and Function After Myocardial Infarction in Mice. J. Am. Heart Assoc. 2017, 6, e004387. [Google Scholar] [CrossRef] [Green Version]
- Blyszczuk, P.; Muller-Edenborn, B.; Valenta, T.; Osto, E.; Stellato, M.; Behnke, S.; Glatz, K.; Basler, K.; Luscher, T.F.; Distler, O.; et al. Transforming growth factor-beta-dependent Wnt secretion controls myofibroblast formation and myocardial fibrosis progression in experimental autoimmune myocarditis. Eur. Heart J. 2017, 38, 1413–1425. [Google Scholar] [CrossRef] [Green Version]
- Daskalopoulos, E.P.; Blankesteijn, W.M. Effect of Interventions in WNT Signaling on Healing of Cardiac Injury: A Systematic Review. Cells 2021, 10, 207. [Google Scholar] [CrossRef]
- Kim, Y.S.; Jeong, H.Y.; Kim, A.R.; Kim, W.H.; Cho, H.; Um, J.; Seo, Y.; Kang, W.S.; Jin, S.W.; Kim, M.C.; et al. Natural product derivative BIO promotes recovery after myocardial infarction via unique modulation of the cardiac microenvironment. Sci. Rep. 2016, 6, 30726. [Google Scholar] [CrossRef] [Green Version]
- Jung, Y.-S.; Park, J.-I. Wnt signaling in cancer: Therapeutic targeting of Wnt signaling beyond β-catenin and the destruction complex. Exp. Mol. Med. 2020, 52, 183–191. [Google Scholar] [CrossRef]
- Foulquier, S.; Daskalopoulos, E.P.; Lluri, G.; Hermans, K.C.M.; Deb, A.; Blankesteijn, W.M. WNT Signaling in Cardiac and Vascular Disease. Pharmacol. Rev. 2018, 70, 68–141. [Google Scholar] [CrossRef] [Green Version]
- Gatica-Andrades, M.; Vagenas, D.; Kling, J.; Nguyen, T.T.K.; Benham, H.; Thomas, R.; Körner, H.; Venkatesh, B.; Cohen, J.; Blumenthal, A. WNT ligands contribute to the immune response during septic shock and amplify endotoxemia-driven inflammation in mice. Blood Adv. 2017, 1, 1274–1286. [Google Scholar] [CrossRef] [Green Version]
- Houschyar, K.S.; Chelliah, M.P.; Rein, S.; Maan, Z.N.; Weissenberg, K.; Duscher, D.; Branski, L.K.; Siemers, F. Role of Wnt signaling during inflammation and sepsis: A review of the literature. Int. J. Artif. Organs 2018, 41, 247–253. [Google Scholar] [CrossRef]
- Sharma, A.; Yang, W.-L.; Ochani, M.; Wang, P. Mitigation of sepsis-induced inflammatory responses and organ injury through targeting Wnt/β-catenin signaling. Sci. Rep. 2017, 7, 9235. [Google Scholar] [CrossRef]
- Mo, Y.; Wang, Y.; Zhang, L.; Yang, L.; Zhou, M.; Li, X.; Li, Y.; Li, G.; Zeng, Z.; Xiong, W.; et al. The role of Wnt signaling pathway in tumor metabolic reprogramming. J. Cancer 2019, 10, 3789–3797. [Google Scholar] [CrossRef] [Green Version]
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Near-optimal probabilistic RNA-seq quantification. Nat. Biotechnol. 2016, 34, 525–527. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Fabregat, A.; Jupe, S.; Matthews, L.; Sidiropoulos, K.; Gillespie, M.; Garapati, P.; Haw, R.; Jassal, B.; Korninger, F.; May, B.; et al. The Reactome Pathway Knowledgebase. Nucleic Acids Res. 2018, 46, D649–D655. [Google Scholar] [CrossRef]
- Trappe, K.; Thomas, D.; Bikou, O.; Kelemen, K.; Lugenbiel, P.; Voss, F.; Becker, R.; Katus, H.A.; Bauer, A. Suppression of persistent atrial fibrillation by genetic knockdown of caspase 3: A pre-clinical pilot study. Eur. Heart J. 2011, 34, 147–157. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Set Name | Description | Genes | FDR |
|---|---|---|---|
| IMMUNE_SYSTEM | Genes involved in immune system | 25 | 6.41 × 10−6 |
| ADAPTIVE_IMMUNE_SYSTEM | Genes involved in adaptive immune system | 19 | 3.02 × 10−5 |
| DEVELOPMENTAL_BIOLOGY | Genes involved in developmental biology | 18 | 2.84 × 10−6 |
| AXON_GUIDANCE | Genes involved in axon guidance | 15 | 2.68 × 10−6 |
| EXTRACELLULAR_MATRIX_ORGANIZATION | Genes involved in extracellular matrix organization | 6 | 6.8 × 10−3 |
| PLATELET_HOMEOSTASIS | Genes involved in platelet homeostasis | 6 | 4.74 × 10−3 |
| COLLAGEN_FORMATION | Genes involved in collagen formation | 6 | 1.00 × 10−3 |
| SIGNALING_BY_ROBO_RECEPTOR | Genes involved in signaling by Robo receptor | 5 | 6.26 × 10−4 |
| TCR_SIGNALING | Genes involved in TCR signaling | 5 | 6.8 × 10−3 |
| SEMAPHORIN_INTERACTIONS | Genes involved in semaphorin interactions | 5 | 1.77 × 10−2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meyer, I.S.; Li, X.; Meyer, C.; Voloshanenko, O.; Pohl, S.; Boutros, M.; Katus, H.A.; Frey, N.; Leuschner, F. Blockade of Wnt Secretion Attenuates Myocardial Ischemia–Reperfusion Injury by Modulating the Inflammatory Response. Int. J. Mol. Sci. 2022, 23, 12252. https://doi.org/10.3390/ijms232012252
Meyer IS, Li X, Meyer C, Voloshanenko O, Pohl S, Boutros M, Katus HA, Frey N, Leuschner F. Blockade of Wnt Secretion Attenuates Myocardial Ischemia–Reperfusion Injury by Modulating the Inflammatory Response. International Journal of Molecular Sciences. 2022; 23(20):12252. https://doi.org/10.3390/ijms232012252
Chicago/Turabian StyleMeyer, Ingmar Sören, Xue Li, Carina Meyer, Oksana Voloshanenko, Susann Pohl, Michael Boutros, Hugo Albert Katus, Norbert Frey, and Florian Leuschner. 2022. "Blockade of Wnt Secretion Attenuates Myocardial Ischemia–Reperfusion Injury by Modulating the Inflammatory Response" International Journal of Molecular Sciences 23, no. 20: 12252. https://doi.org/10.3390/ijms232012252
APA StyleMeyer, I. S., Li, X., Meyer, C., Voloshanenko, O., Pohl, S., Boutros, M., Katus, H. A., Frey, N., & Leuschner, F. (2022). Blockade of Wnt Secretion Attenuates Myocardial Ischemia–Reperfusion Injury by Modulating the Inflammatory Response. International Journal of Molecular Sciences, 23(20), 12252. https://doi.org/10.3390/ijms232012252