
Alterations in Gene Pair Correlations as Potential Diagnostic Markers for Colon Cancer


Abstract
:1. Introduction
2. Results
2.1. Differentially Expressed Genes (DEGs)
2.2. Pathway Analysis
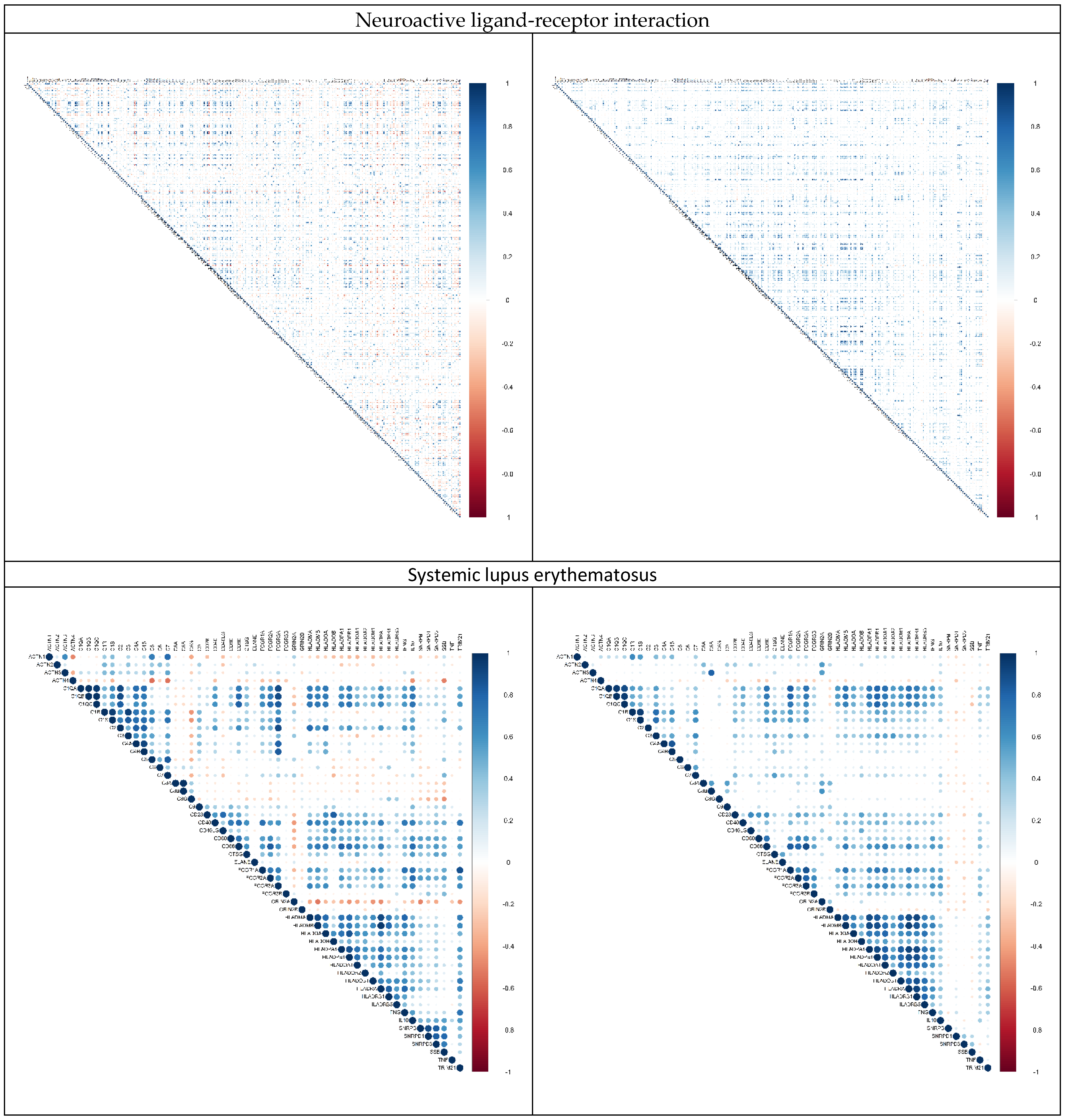
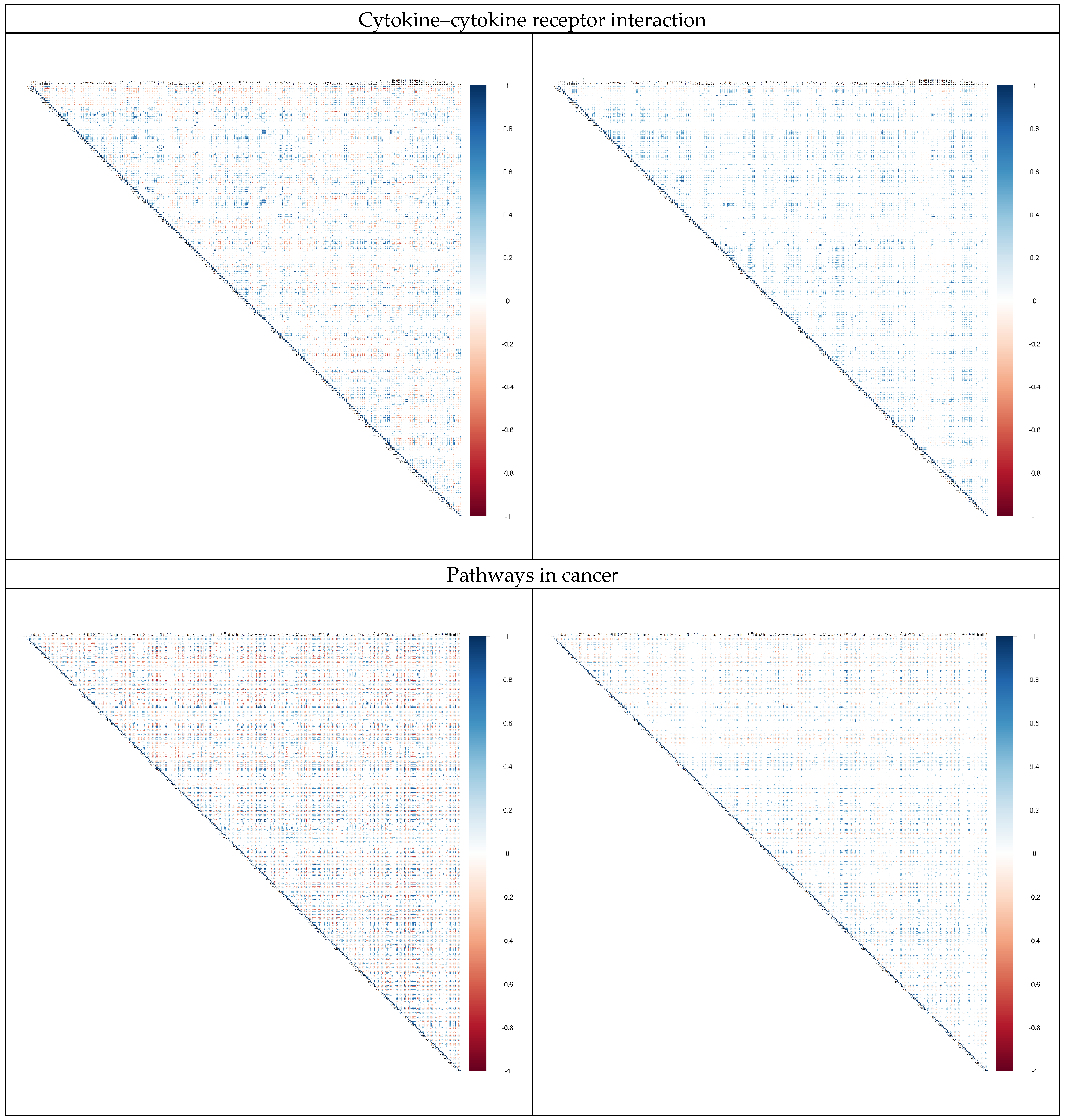
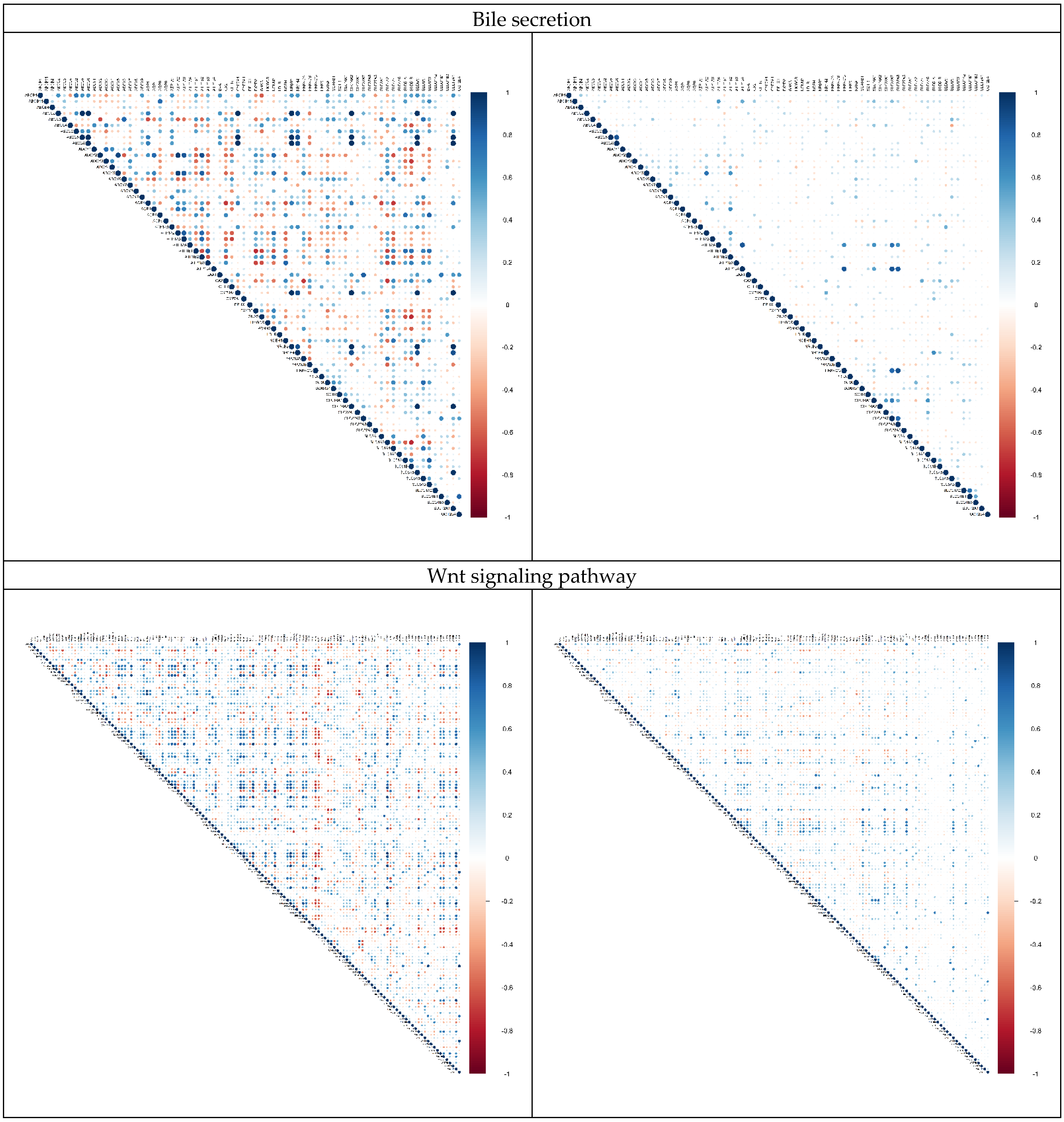
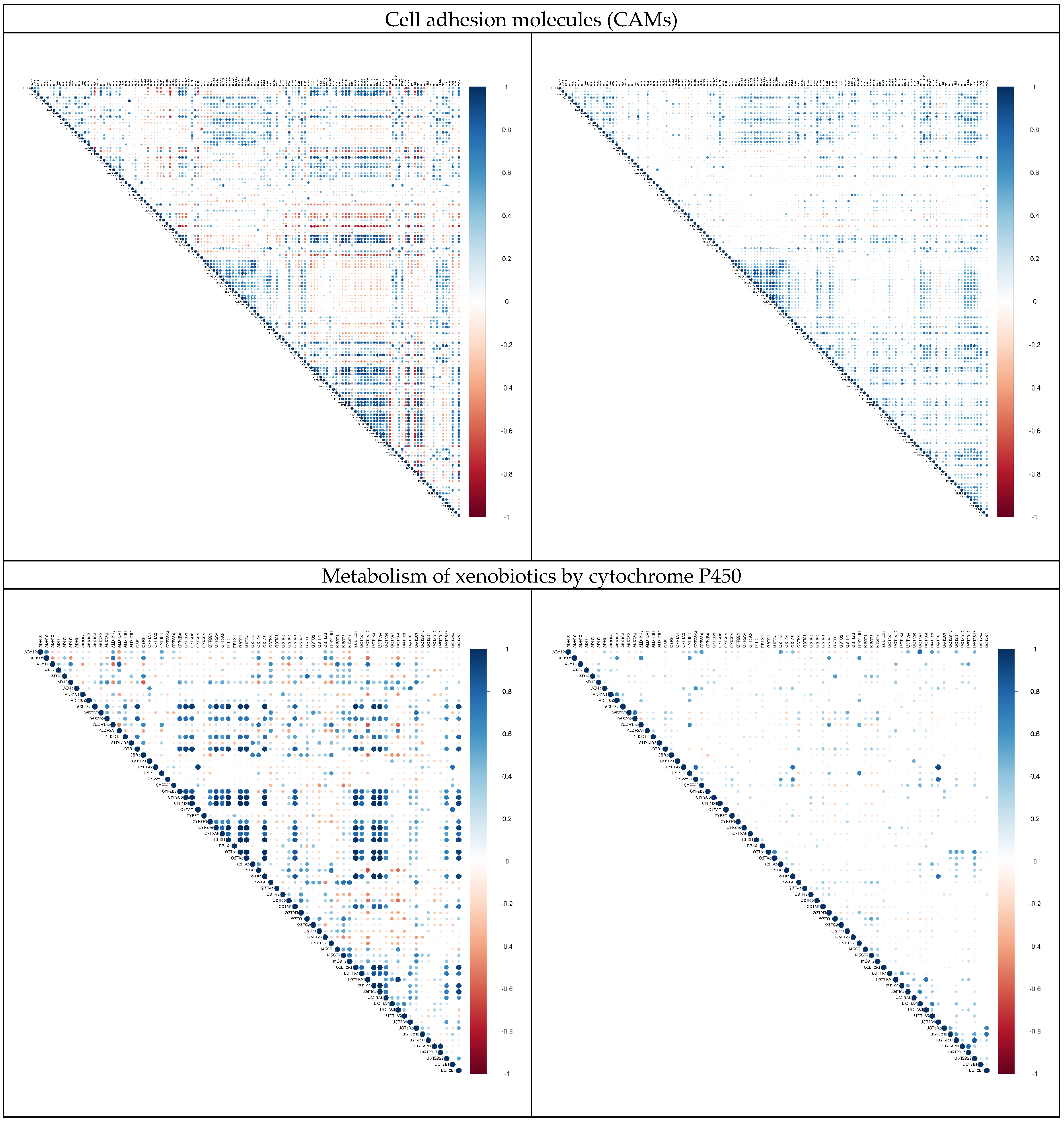
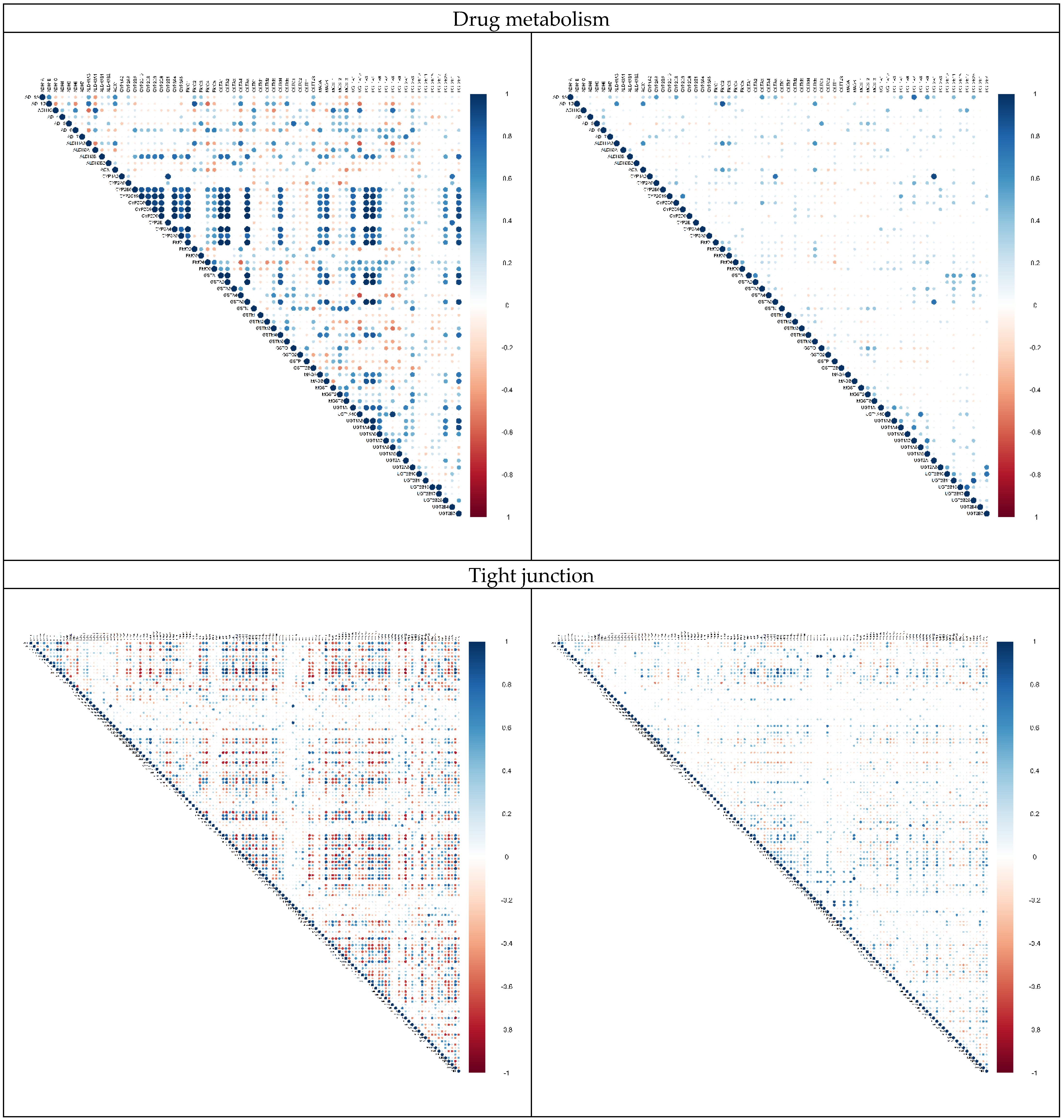
2.3. Alternation in Gene Pair Correlations
3. Discussion
3.1. Top 20 Upregulated DEGs
3.2. Top 20 Downregulated DEGs
3.3. Alternation in Gene Pair Correlations of the DEGs
3.4. Design an Assay for Potential Early Diagnosis of Colon Cancer
4. Materials and Methods
4.1. Data Acquisition
4.2. Identification and Visualization of Differentially Expressed Genes
4.3. Biological Pathway Analysis
4.4. Correlation Matrices
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Goding, S.A.; Fedewa, S.A.; Butterly, L.F.; Anderson, J.C.; Cercek, A.; Smith, R.A.; Jemal, A. Colorectal cancer statistics. CA Cancer J. Clin. 2020, 70, 145–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Available online: https://cancer.ca/en/cancer-information/cancer-types/colorectal/statistics (accessed on 24 December 2021).
- Sawicki, T.; Ruszkowska, M.; Danielewicz, A.; Niedźwiedzka, E.; Arłukowicz, T.; Przybyłowicz, K.E. A review of colorectal cancer in terms of epidemiology, risk factors, development, symptoms and diagnosis. Cancers 2021, 13, 2025. [Google Scholar] [CrossRef] [PubMed]
- Dekker, E.; Tanis, P.J.; Vleugels, J.L.A.; Kasi, P.M.; Wallace, M.B. Colorectal cancer. Lancet 2019, 394, 1467–1480. [Google Scholar] [CrossRef]
- Rawla, P.; Sunkara, T.; Barsouk, A. Epidemiology of colorectal cancer: Incidence, mortality, survival, and risk factors. Prz. Gastroenterol. 2019, 14, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.; Bernstein, C.N.; Samadder, J.N.; Ahmed, R. Screening rates for colorectal cancer in Canada: A cross-sectional study. CMAJ Open 2015, 3, E149–E157. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Läcke, E.; Stock, C.; Hoffmeister, M.; Brenner, H. Colonoscopy and sigmoidoscopy use among older adults in different countries: A systematic review. Prev. Med. 2017, 103, 33–42. [Google Scholar] [CrossRef]
- Brenner, D.R.; Heer, E.; Sutherland, R.L.; Ruan, Y.; Tinmouth, J.; Heitman, S.J.; Hilsden, R.J. National trends in colorectal cancer incidence among older and younger adults in Canada. JAMA Netw. Open 2019, 2, e198090. [Google Scholar] [CrossRef] [Green Version]
- Araghi, M.; Soerjomataram, I.; Bardot, A.; Ferlay, J.; Cabasag, C.J.; Morrison, D.S.; De, P.; Tervonen, H.; Walsh, P.M.; Bucher, O.; et al. Changes in colorectal cancer incidence in seven high-income countries: A population-based study. Lancet. Gastroenterol. Hepatol. 2019, 4, 511–518. [Google Scholar] [CrossRef]
- Siegel, R.; Fedewa, S.A.; Anderson, W.F.; Miller, K.D.; Ma, J.; Rosenberg, P.S.; Jemal, A. Colorectal cancer incidence patterns in the United States, 1974–2013. J. Natl. Cancer Inst. 2017, 109, djw322. [Google Scholar] [CrossRef] [Green Version]
- Bailey, C.E.; Hu, C.Y.; You, Y.N.; Bednarski, B.K.; Rodriguez-Bigas, M.A.; Skibber, J.M.; Cantor, S.B.; Chang, G.J. Increasing disparities in the age-related incidences of colon and rectal cancers in the United States, 1975–2010. JAMA Surg. 2015, 150, 17–22. [Google Scholar] [CrossRef]
- Prabhakaran, S.; Kong, J.C.; Williams, E.; Bell, S.; Warrier, S.; Simpson, P.; Chin, M.; Skinner, S.; Carne, P. Comparison of colorectal cancer outcomes in young adults and octogenarians. Am. J. Surg. 2021, 12, 951–956. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.H.; Chen, Y.X.; Fang, J.Y. Comprehensive review of targeted therapy for colorectal cancer. Sig. Transduct. Target. Ther. 2020, 5, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akond, Z.; Alam, M.; Mollah, M.N.H. Biomarker identification from RNA-Seq data using a robust statistical approach. Bioinformation 2018, 14, 153–163. [Google Scholar] [CrossRef] [Green Version]
- Conesa, A.; Madrigal, P.; Tarazona, S.; Gomez-Cabrero, D.; Cervera, A.; McPherson, A.; Szcześniak, M.W.; Gaffney, D.J.; Elo, L.L.; Zhang, X.; et al. A survey of best practices for RNA-seq data analysis. Genome Biol. 2016, 17, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.; Chan, Y.T.; Tan, H.Y.; Li, S.; Wang, N.; Feng, Y. Epigenetic regulation in human cancer: The potential role of epi-drug in cancer therapy. Mol. Cancer 2020, 19, 79. [Google Scholar] [CrossRef] [PubMed]
- Breuer, K.; Foroushani, A.K.; Laird, M.R.; Chen, C.; Sribnaia, A.; Lo, R.; Winsor, G.L.; Hancock, R.E.; Brinkman, F.S.; Lynn, D.J. InnateDB: Systems biology of innate immunity and beyond--recent updates and continuing curation. Nucleic Acids Res. 2013, 41, D1228–D1233. [Google Scholar] [CrossRef]
- Sakharkar, M.K.; Rajamanickam, K.; Ji, S.; Dhillon, S.K.; Yang, J. Pairwise correlation of genes involved in glucose metabolism: A potential diagnostic marker of cancer. Genes Cancer 2021, 12, 69–76. [Google Scholar] [CrossRef]
- Sakharkar, M.K.; Dhillon, S.K.; Rajamanickam, K.; Heng, B.; Braidy, N.; Guillemin, G.J.; Yang, J. Alteration in gene pair correlations in tryptophan metabolism as a hallmark in cancer diagnosis. Int. J. Tryptophan Res. 2020, 13, 1178646920977013. [Google Scholar] [CrossRef]
- Sakharkar, M.K.; Singh, S.K.K.; Chidambaram, S.B.; Essa, M.M.; Yang, J. Gene pair correlation coefficients in sphingolipid metabolic pathway as a potential diagnostic and prognostic biomarker for breast cancer. Cancers 2020, 12, 1747. [Google Scholar] [CrossRef]
- Poojary, M.; Jishnu, P.V.; Kabekkodu, S.P. Prognostic Value of Melanoma-Associated Antigen-A (MAGE-A) Gene Expression in Various Human Cancers: A Systematic Review and Meta-analysis of 7428 Patients and 44 Studies. Mol. Diagn. Ther. 2020, 24, 537–555. [Google Scholar] [CrossRef]
- Zajac, P.; Schultz-Thater, E.; Tornillo, L.; Sadowski, C.; Trella, E.; Mengus, C.; Iezzi, G.; Spagnoli, G.C. MAGE-A antigens and cancer immunotherapy. MAGE-A Antigens and Cancer Immunotherapy. Front. Med. 2017, 4, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcar, L.; Maclaine, N.J.; Hupp, T.R.; Meek, D.W. Mage-A cancer/testis antigens inhibit p53 function by blocking its interaction with chromatin. Cancer Res. 2010, 70, 10362–10370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monte, M.; Simonatto, M.; Peche, L.Y.; Bublik, D.R.; Gobessi, S.; Pierotti, M.A.; Rodolfo, M.; Schneider, C. MAGE-A tumor antigens target p53 transactivation function through histone deacetylase recruitment and confer resistance to chemotherapeutic agents. Proc. Natl. Acad. Sci. USA 2006, 103, 11160–11165. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; O’Herrin, S.M.; Wu, J.; Reagan-Shaw, S.; Ma, Y.; Bhat, K.M.; Gravekamp, C.; Setaluri, V.; Peters, N.; Hoffmann, F.M.; et al. MAGE-A, mMage-b, and MAGE-C proteins form complexes with KAP1 and suppress p53-dependent apoptosis in MAGE-positive cell lines. Cancer Res. 2007, 67, 9954–9962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Zhao, Y.; Xu, Y.; Sang, M.; Zhao, R.; Gu, L.; Shan, B. MAGE-A genes as predictors of the outcome of laryngeal squamous cell carcinoma. Oncol. Lett. 2020, 20, 59. [Google Scholar] [CrossRef] [PubMed]
- Wei, E.K.; Ma, J.; Pollak, M.N.; Rifai, N.; Fuchs, C.S.; Hankinson, S.E.; Giovannucci, E. A prospective study of C-peptide, insulin-like growth factor-I, insulin-like growth factor binding protein-1, and the risk of colorectal cancer in women. Cancer Epidemiol. Biomark. Prev. 2005, 14, 850–855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.; Pollak, M.N.; Giovannucci, E.; Chan, J.M.; Tao, Y.; Hennekens, C.H.; Stampfer, M.J. Prospective study of colorectal cancer risk in men and plasma levels of insulin-like growth factor (IGF)-I and IGF-binding protein-3. J. Natl. Cancer Inst. 1999, 91, 620–625. [Google Scholar] [CrossRef] [Green Version]
- Vakrakou, A.; Devetzi, M.; Papachristopoulou, G.; Malachias, A.; Scorilas, A.; Xynopoulos, D.; Talieri, M. Kallikrein-related peptidase 6 (KLK6) expression in the progression of colon adenoma to carcinoma. Biol. Chem. 2014, 395, 1105–1117. [Google Scholar] [CrossRef]
- Alexopoulou, D.K.; Kontos, C.K.; Christodoulou, S.; Papadopoulos, I.N.; Scorilas, A. KLK11 mRNA expression predicts poor disease-free and overall survival in colorectal adenocarcinoma patients. Biomark. Med. 2014, 8, 671–685. [Google Scholar] [CrossRef]
- Tailor, P.D.; Kodeboyina, S.K.; Bai, S.; Patel, N.; Sharma, S.; Ratnani, A.; Copland, J.A.; She, J.X.; Sharma, A. Diagnostic and prognostic biomarker potential of kallikrein family genes in different cancer types. Oncotarget 2018, 9, 17876–17888. [Google Scholar] [CrossRef]
- Ho, M.L.; Kuo, W.K.; Chu, L.J.; Yeh, I.H.; Fan, C.W.; Chang, H.I.; Yuan, C.L.; Chou, T.Y.; Chen, H.Y.; Yang, S.W.; et al. N-acetylgalactosamine-6-sulfatase (GALNS), Similar to Glycodelin, Is a Potential General Biomarker for Multiple Malignancies. Anticancer Res. 2019, 39, 6317–6324. [Google Scholar] [CrossRef] [PubMed]
- Govindrajan, R.; Parthasarathy, S. Glycodelin: A possible new biological marker in colorectal cancer. J. Clin. Oncol. 2006, 24, 20081. [Google Scholar] [CrossRef]
- Yoon, J.H.; Kim, D.; Kim, J.; Lee, H.; Ghim, J.; Kang, B.J.; Song, P.; Suh, P.G.; Ryu, S.H.; Lee, T.G. NOTUM Is involved in the progression of colorectal cancer. Cancer Genom. Proteom. 2018, 15, 485–497. [Google Scholar] [CrossRef] [Green Version]
- Nozaki, I.; Lunz, J.G.; Specht, S.; Stolz, D.B.; Taguchi, K.; Subbotin, V.M.; Murase, N.; Demetris, A.J. Small proline-rich proteins 2 are noncoordinately upregulated by IL-6/STAT3 signaling after bile duct ligation. Lab. Investig. 2005, 85, 109–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, Y.; Zheng, X.; Zhang, Y.; Xu, M.; Ye, C.; Lin, M.; Pan, J.; Xu, Z.; Lu, X.; Chi, P. High SPRR1A expression is associated with poor survival in patients with colon cancer. Oncol. Lett. 2020, 19, 3417–3424. [Google Scholar] [CrossRef]
- Pandey, R.; Zhou, M.; Chen, Y.; Darmoul, D.; Kisiel, C.C.; Nfonsam, V.N.; Ignatenko, N.A. Molecular Pathways Associated with Kallikrein 6 Overexpression in Colorectal Cancer. Genes 2021, 12, 749. [Google Scholar] [CrossRef]
- Ghasemzadeh, M.; Kaplan, Z.S.; Alwis, I.; Schoenwaelder, S.M.; Ashworth, K.J.; Westein, E.; Hosseini, E.; Salem, H.H.; Slattery, R.; McColl, S.R.; et al. The CXCR1/2 ligand NAP-2 promotes directed intravascular leukocyte migration through platelet thrombi. Blood 2013, 121, 4555–4566. [Google Scholar] [CrossRef] [Green Version]
- Sreeramkumar, V.; Adrover, J.M.; Ballesteros, I.; Cuartero, M.I.; Rossaint, J.; Bilbao, I.; Nácher, M.; Pitaval, C.; Radovanovic, I.; Fukui, Y.; et al. Neutrophils scan for activated platelets to initiate inflammation. Science 2014, 346, 1234–1238. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.; Lai, M.; Wang, S.; Zheng, K.; Xie, S.X.; Wang, X. Construction of a CXC chemokine-based prediction model for the prognosis of colon cancer. BioMed. Res. Int. 2020, 2020, 1–12. [Google Scholar] [CrossRef]
- Amelung, F.J.; Borstlap, W.A.A.; Consten, E.C.J.; Veld, J.V.; van Halsema, E.E.; Bemelman, W.A.; Siersema, P.D.; Ter Borg, F.; van Hooft, J.E.; Tanis, P.J. Dutch Snapshot Research Group. Propensity score-matched analysis of oncological outcome between stent as bridge to surgery and emergency resection in patients with malignant left-sided colonic obstruction. Br. J. Surg. 2019, 106, 1075–1086. [Google Scholar] [CrossRef]
- Kinouchi, T.; Uemura, M.; Wang, C.; Ishizuya, Y.; Yamamoto, Y.; Hayashi, T.; Matsuzaki, K.; Nakata, W.; Yoshida, T.; Jingushi, K.; et al. Expression level of CXCL7 in peripheral blood cells is a potential biomarker for the diagnosis of renal cell carcinoma. Cancer Sci. 2017, 108, 2495–2502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Q.; Jian, Z.X.; Jia, B.Q.; Chang, L. CXCL7 promotes proliferation and invasion of cholangiocarcinoma cells. Oncol. Rep. 2017, 37, 1114–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, Q.; Li, E.; Liu, Y.; Xie, W.; Huang, C.; Song, J.; Zhang, W.; Zheng, Y.; Wang, H.; Wang, Q. CTAPIII/CXCL7: A novel biomarker for early diagnosis of lung cancer. Cancer Med. 2018, 7, 325–335. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Zhang, L.; Zhang, T.; Qi, X.; Cheng, G.; Xia, L. Serum Chemokine CXCL7 as a potential novel biomarker for obstructive colorectal cancer. Front. Oncol. 2021, 10, 599363. [Google Scholar] [CrossRef] [PubMed]
- Pendás-Franco, N.; García, J.M.; Peña, C.; Valle, N.; Pálmer, H.G.; Heinäniemi, M.; Carlberg, C.; Jiménez, B.; Bonilla, F.; Muñoz, A.; et al. DICKKOPF-4 is induced by TCF/beta-catenin and upregulated in human colon cancer, promotes tumour cell invasion and angiogenesis and is repressed by 1alpha,25-dihydroxyvitamin D3. Oncogene 2008, 24, 4467–4477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsui, A.; Yamaguchi, T.; Maekawa, S.; Miyazaki, C.; Takano, S.; Uetake, T.; Inoue, T.; Otaka, M.; Otsuka, H.; Sato, T.; et al. DICKKOPF-4 and -2 genes are upregulated in human colorectal cancer. Cancer Sci. 2009, 100, 1923–1930. [Google Scholar] [CrossRef]
- Maimaiti, A.; Aizezi, A.; Anniwaer, J.; Ali, B.; Dilixiati, M. Zinc finger of the cerebellum 5 promotes colorectal cancer cell proliferation and cell cycle progression through enhanced CDK1/CDC25c signaling. Arch. Med. Sci. 2019, 17, 449–461. [Google Scholar] [CrossRef]
- Jiang, J.; Liu, H.; Tao, L.; Lin, X.; Yang, Y.; Tan, S.; Wu, B. Let-7d inhibits colorectal cancer cell proliferation through the CST1/p65 pathway. Int. J. Oncol. 2018, 53, 781–790. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Li, T.; Ye, G.; Zhao, L.; Zhang, Z.; Mo, D.; Wang, Y.; Zhang, C.; Deng, H.; Li, G.; et al. High expression of COL10A1 is associated with poor prognosis in colorectal cancer. Onco Targets Ther. 2018, 11, 1571–1581. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Liu, W.; Li, H.; Deng, Q.; Yang, M.; Li, X.; Liang, Z. Identification of key genes and pathways associated with cholangiocarcinoma development based on weighted gene correlation network analysis. Peer J. 2019, 7, e7968. [Google Scholar] [CrossRef]
- Ren, L.; Yi, J.; Li, W.; Zheng, X.; Liu, J.; Wang, J.; Du, G. Apolipoproteins and cancer. Cancer Med. 2019, 8, 7032–7043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roula, D.; Theiler, A.; Luschnig, P.; Sturm, G.J.; Tomazic, P.V.; Marsche, G.; Heinemann, A.; Sturm, E.M. Apolipoprotein A-IV acts as an endogenous anti-inflammatory protein and is reduced in treatment-naïve allergic patients and allergen-challenged mice. Allergy 2020, 75, 392–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.S.; Chang, J.H.; Yu, H.K.; Ahn, J.H.; Yum, J.S.; Lee, S.K. Inhibition of angiogenesis and angiogenesis-dependent tumor growth by the cryptic kringle fragments of human apolipoprotein(a). J. Biol. Chem. 2003, 278, 29000–29008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro, M.A.; Carpintero, R.; Acin, S.; Arbones-Mainar, J.M.; Calleja, L.; Carnicer, R. Immune-regulation of the apolipoprotein A-I/C-III/A-IV gene cluster in experimental inflammation. Cytokine 2005, 31, 52–63. [Google Scholar] [CrossRef] [PubMed]
- Zamanian-Daryoush, M.; Lindner, D.; Tallant, T.C.; Wang, Z.; Buffa, J.; Klipfell, E. The cardioprotective protein apolipoprotein A1 promotes potent anti-tumorigenic effects. J. Biol. Chem. 2013, 288, 21237–21252. [Google Scholar] [CrossRef] [Green Version]
- Feng, W.; Gao, X.; McClung, G.; Zhu, S.; Ishihara, K.; Brash, J.L. Methacrylate polymer layers bearing poly(ethylene oxide) and phosphorylcholine side chains as non-fouling surfaces: In vitro interactions with plasma proteins and platelets. Acta Biomater. 2011, 7, 3692–3699. [Google Scholar] [CrossRef]
- Yui, Y.; Aoyama, T.; Morishita, H.; Takahashi, M.; Takatsu, Y.; Kawai, C. Serum prostacyclin stabilizing factor is identical to apolipoprotein A-I (Apo A-I). A novel function of Apo A-I. J. Clin. Investig. 1988, 82, 803–807. [Google Scholar] [CrossRef]
- Xu, X.; Chen, D.; Feng, X.; Hu, J.; Ge, J.; Yan, C.; Zhang, D.; Ling, Z.; Chen, J.; Wu, J. Apolipoprotein B Is associated with the microenvironment of cholangiocarcinoma. Front. Oncol. 2021, 11, 654689. [Google Scholar] [CrossRef]
- Available online: https://www.news-medical.net/whitepaper/20180123/Role-of-Apolipoprotein-C3-in-lipid-metabolism.aspx (accessed on 24 December 2021).
- Qu, H.; Su, Y.; Yu, L.; Zhao, H.; Xin, C. Wild-type p53 regulates OTOP2 transcription through DNA loop alteration of the promoter in colorectal cancer. FEBS Open Bio. 2018, 9, 26–34. [Google Scholar] [CrossRef] [Green Version]
- Guo, S.; Sun, Y. OTOP2, Inversely Modulated by miR-3148, Inhibits CRC Cell Migration, Proliferation and Epithelial-Mesenchymal Transition: Evidence from Bioinformatics Data Mining and Experimental Verification. Cancer Manag Res. 2022, 14, 1371–1384. [Google Scholar] [CrossRef]
- Yang, H.; Liu, H.; Lin, H.C.; Gan, D.; Jin, W.; Cui, C.; Yan, Y.; Qian, Y.; Han, C.; Wang, Z. Association of a novel seven-gene expression signature with the disease prognosis in colon cancer patients. Aging 2019, 11, 8710–8727. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.T.; Ung, T.T.; Kim, N.H.; Jung, Y.D. Role of bile acids in colon carcinogenesis. World J. Clin. Cases 2018, 6, 577–588. [Google Scholar] [CrossRef]
- Poupon, R. ASBT inhibitors in cholangiopathies-Good for mice, good for men? J. Hepatol. 2016, 64, 537–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, L.; Liu, P.; Zhu, T. Long noncoding RNA SLC30A10 promotes colorectal tumor proliferation and migration via miR-21c/APC axis. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 6682–6691. [Google Scholar] [PubMed]
- Ahmad, T.R.; Higuchi, S.; Bertaggia, E.; Hung, A.; Shanmugarajah, N.; Guilz, N.C.; Gamarra, J.R.; Haeusler, R.A. Bile acid composition regulates the manganese transporter Slc30a10 in intestine. J. Biol. Chem. 2020, 295, 12545–12558. [Google Scholar] [CrossRef]
- Wu, Q.; Yang, Z.F.; Wang, K.J.; Feng, X.Y.; Lv, Z.J.; Li, Y.; Jian, Z.X. AQP8 inhibits colorectal cancer growth and metastasis by down-regulating PI3K/AKT signaling and PCDH7 expression. Am. J. Cancer Res. 2018, 8, 266–279. [Google Scholar]
- Liu, H.; Zhang, C. Identification of differentially expressed genes and their upstream regulators in colorectal cancer. Cancer Gene Ther. 2017, 24, 244–250. [Google Scholar] [CrossRef]
- Yang, X.; Wei, W.; Tan, S.; Guo, L.; Qiao, S.; Yao, B.; Wang, Z. Identification and verification of HCAR3 and INSL5 as new potential therapeutic targets of colorectal cancer. World J. Surg. Oncol. 2021, 19, 248. [Google Scholar] [CrossRef]
- Belgi, A.; Hossain, M.A.; Shabanpoor, F.; Chan, L.; Zhang, S.; Bathgate, R.A.; Tregear, G.W.; Wade, J.D. Structure and function relationship of murine insulin-like peptide 5 (INSL5): Free C-terminus is essential for RXFP4 receptor binding and activation. Biochemistry 2011, 50, 8352–8361. [Google Scholar] [CrossRef]
- De La Cena, K.O.C.; Ho, R.X.; Amraei, R.; Woolf, N.; Tashjian, J.Y.; Zhao, Q.; Richards, S.; Walker, J.; Huang, J.; Chitalia, V.C.; et al. Transmembrane and immunoglobulin domain containing 1, a putative tumor suppressor, induces G2/M cell cycle checkpoint arrest in colon cancer cells. Am. J. Pathol. 2021, 191, 157–167. [Google Scholar] [CrossRef]
- Zhang, H.; Du, Y.; Wang, Z.; Lou, R.; Wu, J.; Feng, J. Integrated analysis of oncogenic networks in colorectal cancer identifies GUCA2A as a molecular marker. Biochem. Res. Int. 2019, 2019, 6469420. [Google Scholar] [CrossRef] [PubMed]
- Fang, S.; Fang, X. Advances in glucose metabolism research in colorectal cancer. Biomed. Rep. 2016, 5, 289–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Guerrero, M.C.; Garcia-Pardo, J.; Berenguer, E.; Fernandez-Alvarez, R.; Barfi, G.B.; Lyons, P.J.; Aviles, F.X.; Huber, R.; Lorenzo, J.; Reverter, D. Crystal structure and mechanism of human carboxypeptidase O: Insights into its specific activity for acidic residues. Proc. Natl. Acad. Sci. USA 2018, 115, E3932–E3939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.; Li, Y.; Cui, Y.; Guo, Y. Identifications of candidate genes significantly associated with rectal cancer by integrated bioinformatics analysis. Technol. Cancer Res. Treat. 2020, 19, 1533033820973270. [Google Scholar] [CrossRef]
- Tseng, W.W.; Liu, C.D. Peptide YY and cancer: Current findings and potential clinical applications. Peptides 2002, 23, 389–395. [Google Scholar] [CrossRef]
- Pan, F.; Chen, T.; Sun, X.; Li, K.; Jiang, X.; Försti, A.; Zhu, Y.; Lai, M. Prognosis prediction of colorectal cancer using gene expression profiles. Front. Oncol. 2019, 9, 252. [Google Scholar] [CrossRef] [Green Version]
- Cheng, B.; Rong, A.; Zhou, Q.; Li, W. CLDN8 promotes colorectal cancer cell proliferation, migration, and invasion by activating MAPK/ERK signaling. Cancer Manag. Res. 2019, 11, 3741–3751. [Google Scholar] [CrossRef] [Green Version]
- Reilly, C.M.; Regna, N.; Mishra, N. HDAC inhibition in lupus models. Mol. Med. 2011, 17, 417–425. [Google Scholar] [CrossRef]
- Ozawa, T.; Hashiguchi, Y.; Yagi, T.; Fukushima, Y.; Shimada, R.; Hayama, T.; Tsuchiya, T.; Nozawa, K.; Iinuma, H.; Ishihara, S.; et al. Angiotensin I-converting enzyme inhibitors/angiotensin II receptor blockers may reduce tumor recurrence in left-sided and early colorectal cancers. Int. J. Colorectal Dis. 2019, 34, 1731–1739. [Google Scholar] [CrossRef]
- Makar, G.A.; Holmes, J.H.; Yang, Y.X. Angiotensin-converting enzyme inhibitor therapy and colorectal cancer risk. J. Natl. Cancer Inst. 2014, 106, djt374. [Google Scholar] [CrossRef] [Green Version]
- Azoulay, L.; Assimes, T.L.; Yin, H.; Bartels, D.B.; Schiffrin, E.L.; Suissa, S. Long-term use of angiotensin receptor blockers and the risk of cancer. PLoS ONE 2012, 7, e50893. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Upregulated | Downregulated | ||||
|---|---|---|---|---|---|
| Gene Name | Log2FC ≥ 2.50 | p-Value | Gene Name | Log2FC ≤ 2.50 | p-Value |
| MAGEA3 | 11.81 | 0 | APOA4 | −8.12 | 0 |
| MAGEA6 | 10.71 | 0 | OTOP2 | −7.95 | 0 |
| IGFL1 | 10.60 | 0 | APOC3 | −7.84 | 0 |
| PRSS56 | 10.55 | 0 | SLC10A2 | −7.16 | 0 |
| MAGEA12 | 10.16 | 0 | APOA1 | −7.10 | 0 |
| KLK6 | 9.74 | 0 | MS4A10 | −7.07 | 0 |
| PAEP | 9.67 | 0 | AQP8 | −6.86 | 0 |
| NOTUM | 9.65 | 0 | CA1 | −6.58 | 0 |
| PRR9 | 9.51 | 0 | APOB | −6.41 | 0 |
| KLK8 | 9.28 | 0 | INSL5 | −6.14 | 0 |
| SPRR2E | 9.21 | 0 | TMIGD1 | −6.07 | 0 |
| PPBP | 9.21 | 0 | GUCA2B | −6.06 | 0 |
| SPRR1A | 9.10 | 0 | G6PC | −6.02 | 0 |
| MAGEA1 | 9.07 | 1.58 × 10−10 | CPO | −5.83 | 0 |
| FEZF1 | 8.99 | 0 | KRTAP13-2 | −5.75 | 0 |
| DKK4 | 8.93 | 0 | PYY | −5.74 | 0 |
| ZIC5 | 8.91 | 3.71 × 10−13 | OTOP3 | −5.73 | 0 |
| KLK7 | 8.87 | 0 | BEST4 | −5.70 | 0 |
| CST1 | 8.58 | 0 | SLC30A10 | −5.57 | 0 |
| SPRR2D | 8.52 | 0 | CLDN8 | −5.55 | 0 |
| Pathway Name | Uploaded Gene Count | Total Number of Genes | |
|---|---|---|---|
| 1 | Neuroactive ligand–receptor interaction | 29 | 275 |
| 2 | Systemic lupus erythematosus | 28 | 126 |
| 3 | Cytokine–cytokine receptor interaction | 27 | 258 |
| 4 | Pathways in cancer | 19 | 329 |
| 5 | Bile secretion | 18 | 72 |
| 6 | Wnt signaling pathway | 17 | 140 |
| 7 | Cell adhesion molecules | 17 | 144 |
| 8 | Metabolism of xenobiotics by cytochrome P450 | 16 | 73 |
| 9 | Drug metabolism | 14 | 67 |
| 19 | Tight junction | 14 | 135 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, B.Y.; Sakharkar, M.K. Alterations in Gene Pair Correlations as Potential Diagnostic Markers for Colon Cancer. Int. J. Mol. Sci. 2022, 23, 12463. https://doi.org/10.3390/ijms232012463
Yang BY, Sakharkar MK. Alterations in Gene Pair Correlations as Potential Diagnostic Markers for Colon Cancer. International Journal of Molecular Sciences. 2022; 23(20):12463. https://doi.org/10.3390/ijms232012463
Chicago/Turabian StyleYang, Bonnie Yang, and Meena Kishore Sakharkar. 2022. "Alterations in Gene Pair Correlations as Potential Diagnostic Markers for Colon Cancer" International Journal of Molecular Sciences 23, no. 20: 12463. https://doi.org/10.3390/ijms232012463
APA StyleYang, B. Y., & Sakharkar, M. K. (2022). Alterations in Gene Pair Correlations as Potential Diagnostic Markers for Colon Cancer. International Journal of Molecular Sciences, 23(20), 12463. https://doi.org/10.3390/ijms232012463