
Identifying Hub Genes Associated with Neoadjuvant Chemotherapy Resistance in Breast Cancer and Potential Drug Repurposing for the Development of Precision Medicine



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Differential Gene Expression Analysis of Breast Cancer Patients Sensitive and Resistant to Chemotherapy
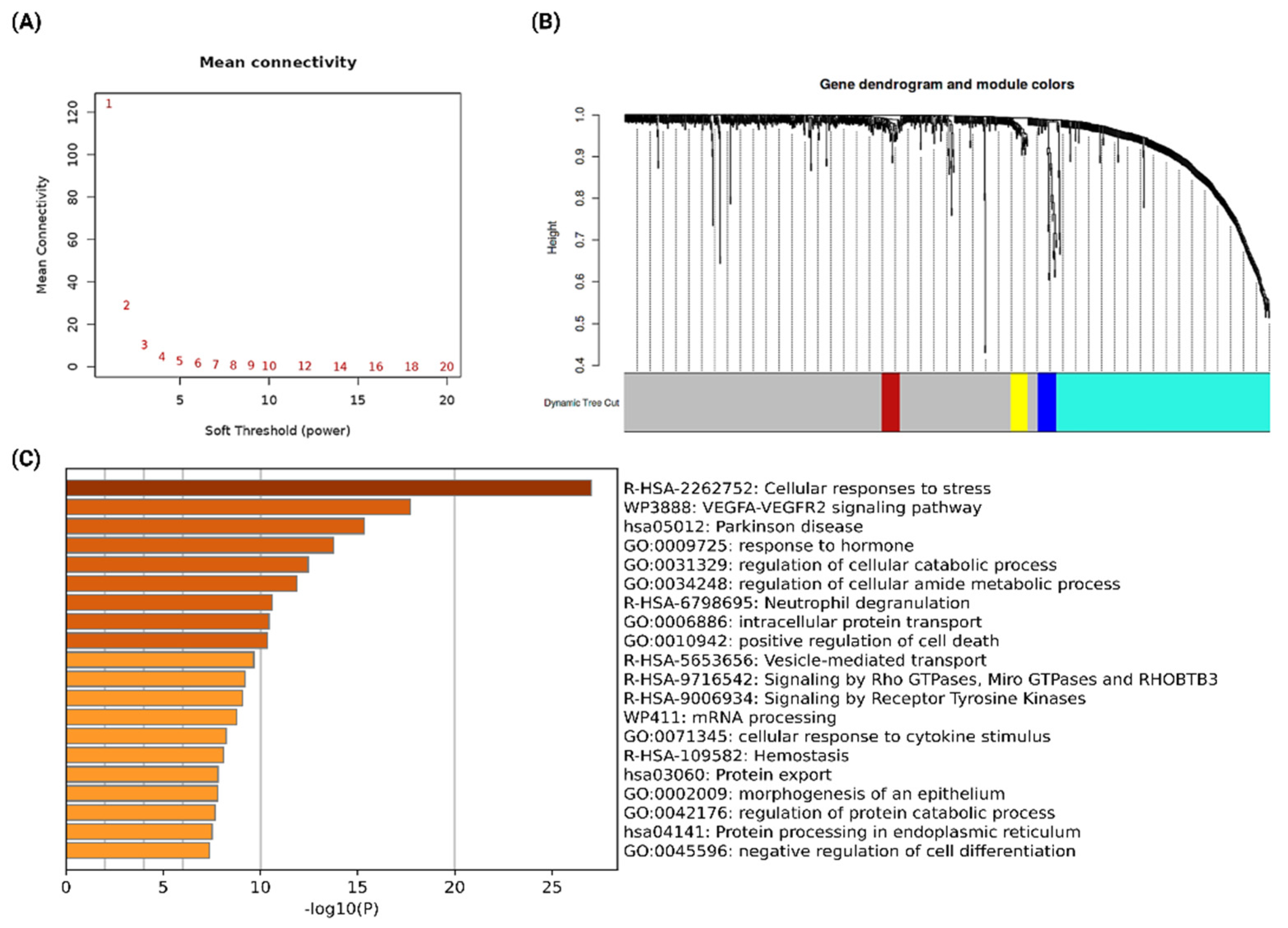
2.2. Co-Expression Network Analysis to Identify the Functionally Significant Co-Expressed Module
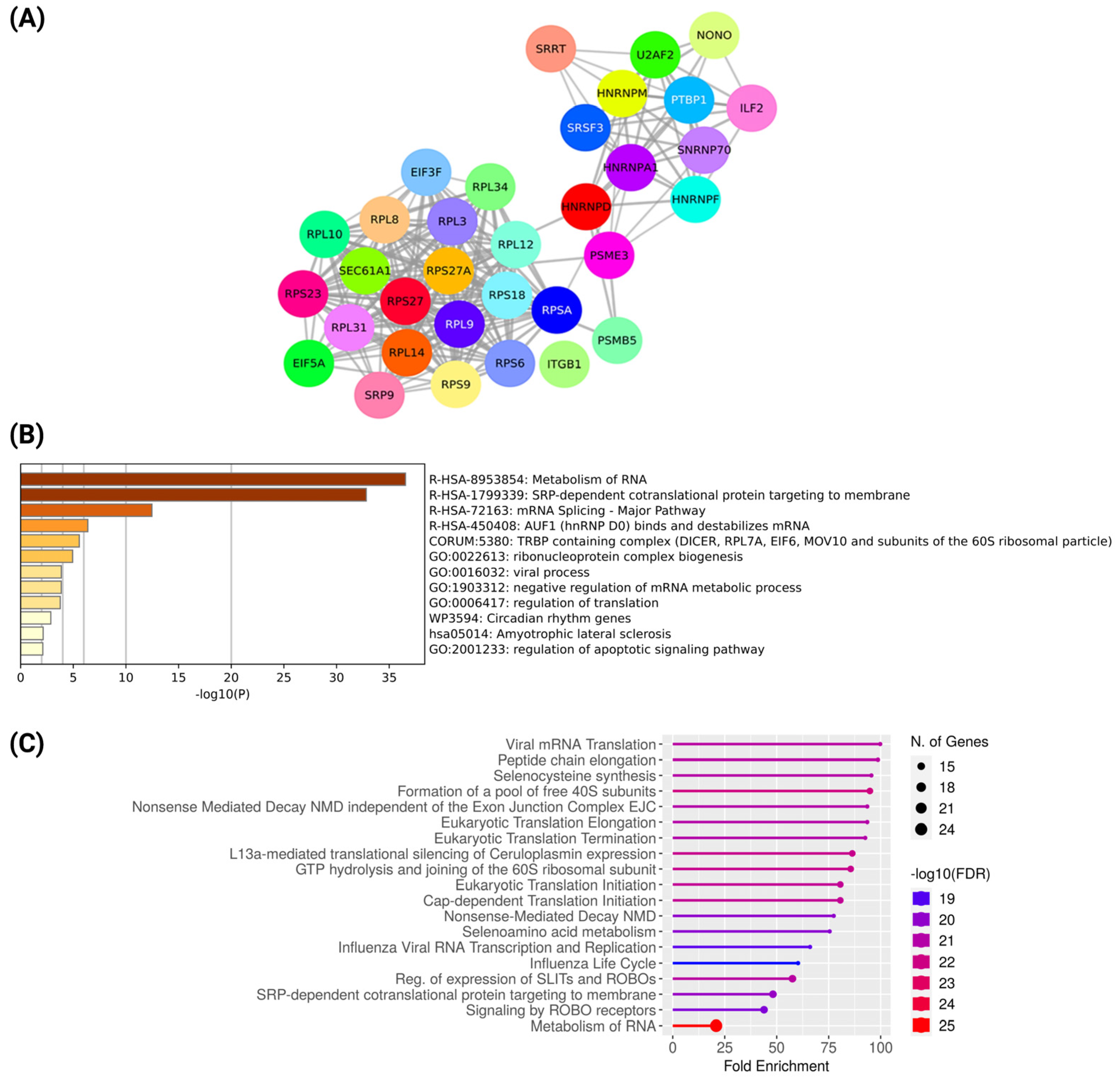
2.3. Network Analysis of Co-Expressed Modules, Identification of Hub Genes from the PPI Network, and Functional Enrichment Analysis
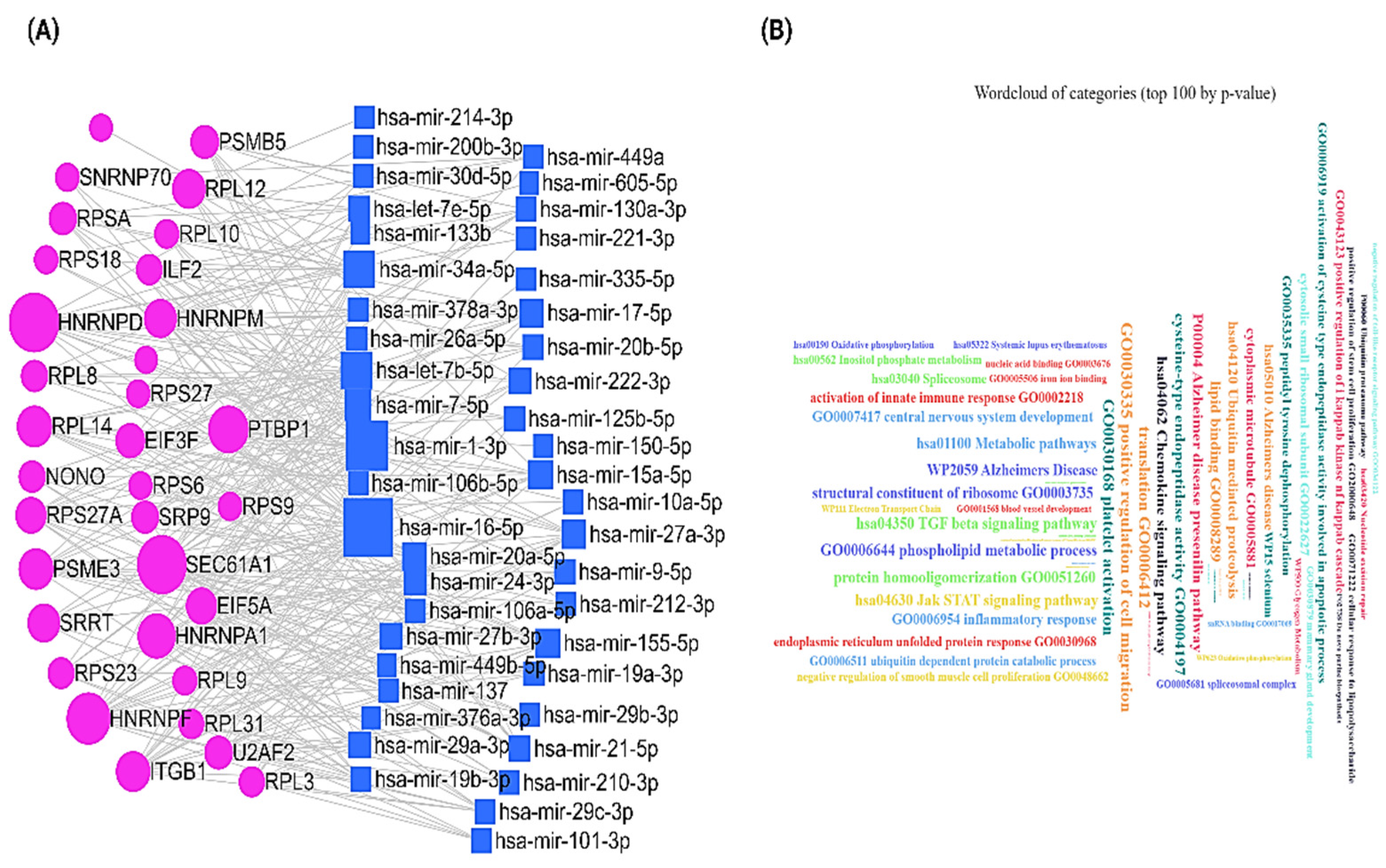
2.4. mRNA-miRNA Network Analysis Reveals the Potentiality of the Hub Genes in the Alternation of Regulatory Network in NAC-Resistant Breast Cancer
2.5. Analysis of the Hub Genes in the Single-Cell RNA-seq Data of Breast Cancer Patients
2.6. Drug Repurposing Using Drug-Gene Interaction Analysis of Hub Genes
2.7. Survival Analysis of the Hub Genes
3. Discussion
4. Materials and Methods
4.1. Acquisition of Transcriptome Dataset
4.2. Read Mapping and Differential Gene Expression Analysis
4.3. Weighted Gene Co-Expression Network Analysis
4.4. Construction of PPI Network and Hub Gene Analysis
4.5. mRNA-miRNA Network Analysis
4.6. Functional Enrichment Analysis of Hub Genes and miRNAs
4.7. Survival Analysis of Hub Genes
4.8. Drug-hub Gene Interaction and Drug Repositioning
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vasan, N.; Baselga, J.; Hyman, D.M. A View on Drug Resistance in Cancer. Nature 2019, 575, 299–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swanton, C. Intratumour Heterogeneity: Evolution through Space and Time. Cancer Res. 2012, 72, 4875–4882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holohan, C.; Van Schaeybroeck, S.; Longley, D.B.; Johnston, P.G. Cancer Drug Resistance: An Evolving Paradigm. Nat. Rev. Cancer 2013, 13, 714–726. [Google Scholar] [CrossRef] [PubMed]
- da Silva-Diz, V.; Lorenzo-Sanz, L.; Bernat-Peguera, A.; Lopez-Cerda, M.; Muñoz, P. Cancer Cell Plasticity: Impact on Tumor Progression and Therapy Response. Semin. Cancer Biol. 2018, 53, 48–58. [Google Scholar] [CrossRef]
- Mukherjee, S.; Heng, H.H.; Frenkel-Morgenstern, M. Emerging Role of Chimeric RNAs in Cell Plasticity and Adaptive Evolution of Cancer Cells. Cancers 2021, 13, 4328. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Frenkel-Morgenstern, M. Evolutionary Impact of Chimeric RNAs on Generating Phenotypic Plasticity in Human Cells. Trends Genet. 2022, 38, 4–7. [Google Scholar] [CrossRef] [PubMed]
- Boumahdi, S.; de Sauvage, F.J. The Great Escape: Tumour Cell Plasticity in Resistance to Targeted Therapy. Nat. Rev. Drug Discov. 2019, 19, 39–56. [Google Scholar] [CrossRef] [PubMed]
- Cosentino, G.; Plantamura, I.; Tagliabue, E.; Iorio, M.V.; Cataldo, A. Breast Cancer Drug Resistance: Overcoming the Challenge by Capitalizing on MicroRNA and Tumor Microenvironment Interplay. Cancers 2021, 13, 3691. [Google Scholar] [CrossRef] [PubMed]
- Saha, T.; Solomon, J.; Samson, A.O.; Gil-Henn, H. Invasion and Metastasis as a Central Hallmark of Breast Cancer. J. Clin. Med. 2021, 10, 3498. [Google Scholar] [CrossRef] [PubMed]
- Cancer Facts & Figures 2022|Enhanced Reader. Available online: https://www.cancer.org/research/cancer-facts-statistics/all-cancer-facts-figures/cancer-facts-figures-2022.html (accessed on 7 October 2022).
- Ji, X.; Lu, Y.; Tian, H.; Meng, X.; Wei, M.; Cho, W.C. Chemoresistance Mechanisms of Breast Cancer and Their Countermeasures. Biomed. Pharmacother. 2019, 114, 108800. [Google Scholar] [CrossRef] [PubMed]
- Nabholtz, J.M.; Gligorov, J. The Role of Taxanes in the Treatment of Breast Cancer. Expert. Opin. Pharmacother. 2005, 6, 1073–1094. [Google Scholar] [CrossRef] [PubMed]
- Sunters, A.; Madureira, P.A.; Pomeranz, K.M.; Aubert, M.; Brosens, J.J.; Cook, S.J.; Burgering, B.M.T.; Coombes, R.C.; Lam, E.W.F. Paclitaxel-Induced Nuclear Translocation of FOXO3a in Breast Cancer Cells Is Mediated by c-Jun NH2-Terminal Kinase and Akt. Cancer Res. 2006, 66, 212–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- KJ, P. Preclinical Mechanisms of Action of Docetaxel and Docetaxel Combinations in Prostate Cancer. Semin. Oncol. 2001, 28, 3–7. [Google Scholar] [CrossRef]
- Tian, H.; Ma, D.; Tan, X.; Yan, W.; Wu, X.; He, C.; Zhong, L.; Zhang, Y.; Yu, B.; Zhang, Y.; et al. Platinum and Taxane Based Adjuvant and Neoadjuvant Chemotherapy in Early Triple-Negative Breast Cancer: A Narrative Review. Front. Pharmacol. 2021, 12, 770663. [Google Scholar] [CrossRef] [PubMed]
- Abu Samaan, T.M.; Samec, M.; Liskova, A.; Kubatka, P.; Büsselberg, D. Paclitaxel’s Mechanistic and Clinical Effects on Breast Cancer. Biomolecules 2019, 9, 789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikuła-Pietrasik, J.; Witucka, A.; Pakuła, M.; Uruski, P.; Begier-Krasińska, B.; Niklas, A.; Tykarski, A.; Książek, K. Comprehensive Review on How Platinum- and Taxane-Based Chemotherapy of Ovarian Cancer Affects Biology of Normal Cells. Cell. Mol. Life Sci. 2018, 76, 681–697. [Google Scholar] [CrossRef] [Green Version]
- Orr, G.A.; Verdier-Pinard, P.; McDaid, H.; Horwitz, S.B. Mechanisms of Taxol Resistance Related to Microtubules. Oncogene 2003, 22, 7280–7295. [Google Scholar] [CrossRef] [Green Version]
- Nikolaou, M.; Pavlopoulou, A.; Georgakilas, A.G.; Kyrodimos, E. The Challenge of Drug Resistance in Cancer Treatment: A Current Overview. Clin. Exp. Metastasis 2018, 35, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, S.; Miyoshi, Y.; Egawa, C.; Ishitobi, M.; Taguchi, T.; Tamaki, Y.; Monden, M.; Noguchi, S. Prediction of Response to Docetaxel by Quantitative Analysis of Class I and III β-Tubulin Isotype MRNA Expression in Human Breast Cancers. Clin. Cancer Res. 2003, 9, 2992–2997. [Google Scholar] [PubMed]
- Maloney, S.M.; Hoover, C.A.; Morejon-Lasso, L.V.; Prosperi, J.R. Mechanisms of Taxane Resistance. Cancers 2020, 12, 3323. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Gupta, P.; Srivastava, S.K. Penfluridol Overcomes Paclitaxel Resistance in Metastatic Breast Cancer. Sci. Rep. 2019, 9, 5066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, A.; Yu, Z.; Song, Y.; Zhang, E. Silencing of Glutaminase 1 Resensitizes Taxol-Resistant Breast Cancer Cells to Taxol. Mol. Med. Rep. 2015, 11, 4727–4733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, L.; Zhou, Y.; Li, X.; Lin, S.; Tan, Z.; Guan, F. Integrating Transcriptomics, Proteomics, Glycomics and Glycoproteomics to Characterize Paclitaxel Resistance in Breast Cancer Cells. J. Proteomics 2021, 243, 104266. [Google Scholar] [CrossRef]
- Chi, Y.; Xue, J.; Huang, S.; Xiu, B.; Su, Y.; Wang, W.; Guo, R.; Wang, L.; Li, L.; Shao, Z.; et al. CapG Promotes Resistance to Paclitaxel in Breast Cancer through Transactivation of PIK3R1/P50. Theranostics 2019, 9, 6840–6855. [Google Scholar] [CrossRef] [PubMed]
- Lai, D.; Ho, K.C.; Hao, Y.; Yang, X. Taxol Resistance in Breast Cancer Cells Is Mediated by the Hippo Pathway Component TAZ and Its Downstream Transcriptional Targets Cyr61 and CTGF. Cancer Res. 2011, 71, 2728–2738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, I.; Shalli, K.; McDonald, S.L.; Moir, S.E.; Hutcheon, A.W.; Heys, S.D.; Schofield, A.C. Reduced Expression of P27 Is a Novel Mechanism of Docetaxel Resistance in Breast Cancer Cells. Breast Cancer Res. 2004, 6, R601–R607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De, S.; Cipriano, R.; Jackson, M.W.; Stark, G.R. Overexpression of Kinesins Mediates Docetaxel Resistance in Breast Cancer Cells. Cancer Res. 2009, 69, 8035–8042. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Han, L.; Yuan, Y.; Li, J.; Hei, N.; Liang, H. Gene Co-Expression Network Analysis Reveals Common System-Level Properties of Prognostic Genes across Cancer Types. Nat. Commun. 2014, 5, 3231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, S.; Banerjee, B.; Karasik, D.; Frenkel-Morgenstern, M. MRNA-LncRNA Co-Expression Network Analysis Reveals the Role of LncRNAs in Immune Dysfunction during Severe SARS-CoV-2 Infection. Viruses 2021, 13, 402. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, R.Q.; Raschka, T.; Kodamullil, A.T.; Hofmann-Apitius, M.; Mubeen, S.; Domingo-Fernández, D. Towards a Global Investigation of Transcriptomic Signatures through Co-Expression Networks and Pathway Knowledge for the Identification of Disease Mechanisms. Nucleic Acids Res. 2021, 49, 7939–7953. [Google Scholar] [CrossRef]
- Wang, X.; Bove, A.M.; Simone, G.; Ma, B. Molecular Bases of VEGFR-2-Mediated Physiological Function and Pathological Role. Front. Cell Dev. Biol. 2020, 8, 599281. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.; Lovly, C.M. Mechanisms of Receptor Tyrosine Kinase Activation in Cancer. Mol. Cancer 2018, 17, 58. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Morris, J.H.; Cook, H.; Kuhn, M.; Wyder, S.; Simonovic, M.; Santos, A.; Doncheva, N.T.; Roth, A.; Bork, P.; et al. The STRING Database in 2017: Quality-Controlled Protein-Protein Association Networks, Made Broadly Accessible. Nucleic Acids Res. 2017, 45, D362–D368. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yang, D. The Regulation of RNA Metabolism in Hormone Signaling and Breast Cancer. Mol. Cell. Endocrinol. 2021, 529, 111221. [Google Scholar] [CrossRef] [PubMed]
- Fabbri, L.; Chakraborty, A.; Robert, C.; Vagner, S. The Plasticity of MRNA Translation during Cancer Progression and Therapy Resistance. Nat. Rev. Cancer 2021, 21, 558–577. [Google Scholar] [CrossRef] [PubMed]
- Gara, R.K.; Kumari, S.; Ganju, A.; Yallapu, M.M.; Jaggi, M.; Chauhan, S.C. Slit/Robo Pathway: A Promising Therapeutic Target for Cancer. Drug Discov. Today 2015, 20, 156–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasad, A.; Fernandis, A.Z.; Rao, Y.; Ganju, R.K. Slit Protein-Mediated Inhibition of CXCR4-Induced Chemotactic and Chemoinvasive Signaling Pathways in Breast Cancer Cells. J. Biol. Chem. 2004, 279, 9115–9124. [Google Scholar] [CrossRef] [Green Version]
- Yuasa-Kawada, J.; Kinoshita-Kawada, M.; Rao, Y.; Wu, J.Y. Deubiquitinating Enzyme USP33/VDU1 Is Required for Slit Signaling in Inhibiting Breast Cancer Cell Migration. Proc. Natl. Acad. Sci. USA 2009, 106, 14530–14535. [Google Scholar] [CrossRef] [Green Version]
- Kalimuthu, K.; Keerthana, C.K.; Mohan, M.; Arivalagan, J.; Christyraj, J.R.S.S.; Firer, M.A.; Choudry, M.H.A.; Anto, R.J.; Lee, Y.J. The Emerging Role of Selenium Metabolic Pathways in Cancer: New Therapeutic Targets for Cancer. J. Cell. Biochem. 2022, 123, 532–542. [Google Scholar] [CrossRef]
- Voorde, J.V.; Ackermann, T.; Pfetzer, N.; Sumpton, D.; Mackay, G.; Kalna, G.; Nixon, C.; Blyth, K.; Gottlieb, E.; Tardito, S. Improving the Metabolic Fidelity of Cancer Models with a Physiological Cell Culture Medium. Sci. Adv. 2019, 5, eaau7314. [Google Scholar] [CrossRef]
- Song, X.; Wang, X.; Liu, Z.; Yu, Z. Role of GPX4-Mediated Ferroptosis in the Sensitivity of Triple Negative Breast Cancer Cells to Gefitinib. Front. Oncol. 2020, 10, 597434. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhao, M. Gene Dosage Analysis on the Single-Cell Transcriptomes Linking Cotranslational Protein Targeting to Metastatic Triple-Negative Breast Cancer. Pharmaceuticals 2021, 14, 918. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.; Zhou, G.; Soufan, O.; Xia, J. MiRNet 2.0: Network-Based Visual Analytics for MiRNA Functional Analysis and Systems Biology. Nucleic Acids Res. 2020, 48, W244–W251. [Google Scholar] [CrossRef]
- Kern, F.; Fehlmann, T.; Solomon, J.; Schwed, L.; Grammes, N.; Backes, C.; van Keuren-Jensen, K.; Craig, D.W.; Meese, E.; Keller, A. MiEAA 2.0: Integrating Multi-Species MicroRNA Enrichment Analysis and Workflow Management Systems. Nucleic Acids Res. 2020, 48, W521–W528. [Google Scholar] [CrossRef] [PubMed]
- Chung, W.; Eum, H.H.; Lee, H.O.; Lee, K.M.; Lee, H.B.; Kim, K.T.; Ryu, H.S.; Kim, S.; Lee, J.E.; Park, Y.H.; et al. Single-Cell RNA-Seq Enables Comprehensive Tumour and Immune Cell Profiling in Primary Breast Cancer. Nat. Commun. 2017, 8, 15081. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.Z.; Al-Eryani, G.; Roden, D.L.; Junankar, S.; Harvey, K.; Andersson, A.; Thennavan, A.; Wang, C.; Torpy, J.R.; Bartonicek, N.; et al. A Single-Cell and Spatially Resolved Atlas of Human Breast Cancers. Nat. Genet. 2021, 53, 1334–1347. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Yan, M.; Zhang, G.; Liu, W.; Deng, C.; Liao, G.; Xu, L.; Luo, T.; Yan, H.; Long, Z.; et al. CancerSEA: A Cancer Single-Cell State Atlas. Nucleic Acids Res. 2019, 47, D900–D908. [Google Scholar] [CrossRef] [Green Version]
- Braune, E.B.; Tsoi, Y.L.; Phoon, Y.P.; Landor, S.; Cascales, H.S.; Ramsköld, D.; Deng, Q.; Lindqvist, A.; Lian, X.; Sahlgren, C.; et al. Loss of CSL unlocks a hypoxic response and enhanced tumor growth potential in breast cancer cells. B. Loss of CSL unlocks a hypoxic response and enhanced tumor growth potential in breast cancer cells. Stem Cell Rep. 2016, 6, 643–651. [Google Scholar]
- Jordan, N.V.; Bardia, A.; Wittner, B.S.; Benes, C.; Ligorio, M.; Zheng, Y.; Yu, M.; Sundaresan, T.K.; Licausi, J.A.; Desai, R. HER2 expression identifies dynamic functional states within circulating breast cancer cells. Nature 2016, 537, 102–106. [Google Scholar]
- Aceto, N.; Bardia, A.; Wittner, B.S.; Donaldson, M.C.; O'Keefe, R.; Engstrom, A.; Bersani, F.; Zheng, Y.; Comaills, V.; Niederhoffer, K.; et al. AR Expression in Breast Cancer CTCs Associates with Bone MetastasesA Role for AR in Breast Cancer Bone Metastasis. Mol. Cancer Res. 2018, 16, 720–727. [Google Scholar]
- Li, L.-Y.; Guan, Y.-d.; Chen, X.-S.; Yang, J.-M.; Cheng, Y. DNA Repair Pathways in Cancer Therapy and Resistance. Front. Pharmacol. 2021, 11, 629266. [Google Scholar] [CrossRef]
- Győrffy, B. Survival Analysis across the Entire Transcriptome Identifies Biomarkers with the Highest Prognostic Power in Breast Cancer. Comput. Struct. Biotechnol. J. 2021, 19, 4101–4109. [Google Scholar] [CrossRef]
- Lüönd, F.; Tiede, S.; Christofori, G. Breast Cancer as an Example of Tumour Heterogeneity and Tumour Cell Plasticity during Malignant Progression. Br. J. Cancer 2021, 125, 164–175. [Google Scholar] [CrossRef]
- Asselain, B.; Barlow, W.; Bartlett, J.; Bergh, J.; Bergsten-Nordström, E.; Bliss, J.; Boccardo, F.; Boddington, C.; Bogaerts, J.; Bonadonna, G.; et al. Long-Term Outcomes for Neoadjuvant versus Adjuvant Chemotherapy in Early Breast Cancer: Meta-Analysis of Individual Patient Data from Ten Randomised Trials. Lancet Oncol. 2018, 19, 27–39. [Google Scholar] [CrossRef] [Green Version]
- Mougalian, S.S.; Soulos, P.R.; Killelea, B.K.; Lannin, D.R.; Abu-Khalaf, M.M.; Digiovanna, M.P.; Sanft, T.B.; Pusztai, L.; Gross, C.P.; Chagpar, A.B. Use of Neoadjuvant Chemotherapy for Patients with Stage I to III Breast Cancer in the United States. Cancer 2015, 121, 2544–2552. [Google Scholar] [CrossRef] [PubMed]
- Rudrapal, M.; Khairnar, S.J.; Jadhav, A.G.; Rudrapal, M.; Khairnar, S.J.; Jadhav, A.G. Drug Repurposing (DR): An Emerging Approach in Drug Discovery. In Drug Repurposing: Hypothesis, Molecular Aspects and Therapeutic Applications; IntechOpen: London, UK, 2020. [Google Scholar] [CrossRef]
- Park, J.E.; Park, J.; Jun, Y.; Oh, Y.; Ryoo, G.; Jeong, Y.S.; Gadalla, H.H.; Min, J.S.; Jo, J.H.; Song, M.G.; et al. Expanding Therapeutic Utility of Carfilzomib for Breast Cancer Therapy by Novel Albumin-Coated Nanocrystal Formulation. J. Control. Release 2019, 302, 148–159. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.H.; Gonzalez-Angulo, A.M.; Reuben, J.M.; Booser, D.J.; Pusztai, L.; Krishnamurthy, S.; Esseltine, D.; Stec, J.; Broglio, K.R.; Islam, R.; et al. Bortezomib (VELCADE) in Metastatic Breast Cancer: Pharmacodynamics, Biological Effects, and Prediction of Clinical Benefits. Ann. Oncol. 2006, 17, 813–817. [Google Scholar] [CrossRef] [PubMed]
- Landys, K.; Borgstrom, S.; Andersson, T.; Noppa, H. Mitoxantrone as a First-Line Treatment of Advanced Breast Cancer. Investig. New Drugs 1985, 3, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Yakhni, M.; Briat, A.; Guerrab, A.E.; Furtado, L.; Kwiatkowski, F.; Miot-Noirault, E.; Cachin, F.; Penault-Llorca, F.; Radosevic-Robin, N. Homoharringtonine, an Approved Anti-Leukemia Drug, Suppresses Triple Negative Breast Cancer Growth through a Rapid Reduction of Anti-Apoptotic Protein Abundance. Am. J. Cancer Res. 2019, 9, 1043–1060. [Google Scholar]
- Palmer, A.C.; Sorger, P.K. Combination Cancer Therapy Can Confer Benefit via Patient-to-Patient Variability without Drug Additivity or Synergy. Cell 2017, 171, 1678–1691.e13. [Google Scholar] [CrossRef] [Green Version]
- Detroja, T.S.; Gil-Henn, H.; Samson, A.O. Text-Mining Approach to Identify Hub Genes of Cancer Metastasis and Potential Drug Repurposing to Target Them. J. Clin. Med. 2022, 11, 2130. [Google Scholar] [CrossRef]
- Jaaks, P.; Coker, E.A.; Vis, D.J.; Edwards, O.; Carpenter, E.F.; Leto, S.M.; Dwane, L.; Sassi, F.; Lightfoot, H.; Barthorpe, S.; et al. Effective Drug Combinations in Breast, Colon and Pancreatic Cancer Cells. Nature 2022, 603, 166–173. [Google Scholar] [CrossRef]
- Sun, L.; Liu, L.; Liu, X.; Wang, Y.; Li, M.; Yao, L.; Yang, J.; Ji, G.; Guo, C.; Pan, Y.; et al. MGr1-Ag/37LRP Induces Cell Adhesion-Mediated Drug Resistance through FAK/PI3K and MAPK Pathway in Gastric Cancer. Cancer Sci. 2014, 105, 651–659. [Google Scholar] [CrossRef]
- Chetty, C.J.; Ferreira, E.; Jovanovic, K.; Weiss, S.F.T. Knockdown of LRP/LR Induces Apoptosis in Pancreatic Cancer and Neuroblastoma Cells through Activation of Caspases. Exp. Cell Res. 2017, 360, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Khumalo, T.; Ferreira, E.; Jovanovic, K.; Veale, R.B.; Weiss, S.F.T. Knockdown of LRP/LR Induces Apoptosis in Breast and Oesophageal Cancer Cells. PLoS ONE 2015, 10, e0139584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Tan, X.; Liu, P.; Yang, Y.; Huang, Y.; Liu, X.; Meng, X.; Yu, B.; Wu, M.; Jin, H. ITGA6 and RPSA Synergistically Promote Pancreatic Cancer Invasion and Metastasis via PI3K and MAPK Signaling Pathways. Exp. Cell Res. 2019, 379, 30–47. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Gu, B.; Wang, Y.; Shen, S.; Huang, W. E2F1-Mediated MNX1-AS1-MiR-218-5p-SEC61A1 Feedback Loop Contributes to the Progression of Colon Adenocarcinoma. J. Cell. Biochem. 2019, 120, 6145–6153. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Zhao, L.; Guo, C.; Liu, C.; Liu, Y. Identification of a Novel DNA Repair-Related Prognostic Signature Predicting Survival of Patients with Hepatocellular Carcinoma. Cancer Manag. Res. 2019, 11, 7473–7484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Zeng, J.; Li, H.; Li, F.; Jiang, B.; Zhao, M.; Liu, Z.; Li, R.; Ma, T. A Risk Model Based on Sorafenib-Response Target Genes Predicts the Prognosis of Patients with HCC. J. Oncol. 2022, 2022, 7257738. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Hu, N.; Song, X.; Yang, J. Hsa_Circ_0007841 Enhances Multiple Myeloma Chemotherapy Resistance Through Upregulating ABCG2. Technol. Cancer Res. Treat. 2020, 19, 1533033820928371. [Google Scholar] [CrossRef]
- Wang, Y.; Lin, Q.; Song, C.; Ma, R.; Li, X. Depletion of Circ_0007841 Inhibits Multiple Myeloma Development and BTZ Resistance via MiR-129-5p/JAG1 Axis. Cell. Cycle 2020, 19, 3289–3302. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; Zhou, S.; Hu, J. Long Noncoding RNA MAGI2-AS3/MiR-218-5p/GDPD5/SEC61A1 Axis Drives Cellular Proliferation and Migration and Confers Cisplatin Resistance in Nasopharyngeal Carcinoma. Int. Forum. Allergy Rhinol. 2020, 10, 1012–1023. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, S.; Gao, S.; Ma, Y.; Tan, X.; Kang, Y.; Ren, W. HIF-1α, TWIST-1 and ITGB-1, Associated with Tumor Stiffness, as Novel Predictive Markers for the Pathological Response to Neoadjuvant Chemotherapy in Breast Cancer. Cancer Manag. Res. 2020, 12, 2209–2222. [Google Scholar] [CrossRef] [PubMed]
- Baltes, F.; Pfeifer, V.; Silbermann, K.; Caspers, J.; Wantoch von Rekowski, K.; Schlesinger, M.; Bendas, G. B1-Integrin Binding to Collagen Type 1 Transmits Breast Cancer Cells into Chemoresistance by Activating ABC Efflux Transporters. Biochim. Biophys. Acta Mol. Cell. Res. 2020, 1867, 118663. [Google Scholar] [CrossRef]
- Huang, L.; Hu, C.; Chao, H.; Wang, R.; Lu, H.; Li, H.; Chen, H. MiR-29c Regulates Resistance to Paclitaxel in Nasopharyngeal Cancer by Targeting ITGB1. Exp. Cell Res. 2019, 378, 1–10. [Google Scholar] [CrossRef]
- Zhu, T.; Zhu, T.; Chen, R.; Chen, R.; Wang, J.; Wang, J.; Yue, H.; Yue, H.; Lu, X.; Lu, X.; et al. The Prognostic Value of ITGA and ITGB Superfamily Members in Patients with High Grade Serous Ovarian Cancer. Cancer Cell Int. 2020, 20, 257. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Zhu, X.; Mei, D.; Ding, Z. Caveolin-1 Contributes to Anoikis Resistance in Human Gastric Cancer SGC-7901 Cells via Regulating Src-Dependent EGFR-ITGB1 Signaling. J. Biochem. Mol. Toxicol. 2018, 32, e22202. [Google Scholar] [CrossRef]
- Ueki, Y.; Saito, K.; Iioka, H.; Sakamoto, I.; Kanda, Y.; Sakaguchi, M.; Horii, A.; Kondo, E. PLOD2 Is Essential to Functional Activation of Integrin Β1 for Invasion/Metastasis in Head and Neck Squamous Cell Carcinomas. iScience 2020, 23, 100850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Zhang, S.; Liu, J.; Tian, Y.; Ma, B.; Xu, S.; Fu, Y.; Luo, Y. Secreted Pyruvate Kinase M2 Promotes Lung Cancer Metastasis through Activating the Integrin Beta1/FAK Signaling Pathway. Cell. Rep. 2020, 30, 1780–1797.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, W.; Zou, Y.; Jiang, Q.; Zhou, Z.; Ding, H.; Yan, L.; Yang, S. Psmb5 Is Associated with Proliferation and Drug Resistance in Triple-Negative Breast Cance. Int. J. Biol. Mark. 2018, 33, 102–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.-Y.; Li, C.-Y.; Hsu, H.-P.; Cho, C.-Y.; Yen, M.-C.; Weng, T.-Y.; Chen, W.-C.; Hung, Y.-H.; Lee, K.-T.; Hung, J.-H.; et al. PSMB5 Plays a Dual Role in Cancer Development and Immunosuppression. Am. J. Cancer Res. 2017, 7, 2103–2120. [Google Scholar] [PubMed]
- Lü, S.; Chen, Z.; Yang, J.; Chen, L.; Gong, S.; Zhou, H.; Guo, L.; Wang, J. Overexpression of the PSMB5 Gene Contributes to Bortezomib Resistance in T-Lymphoblastic Lymphoma/Leukemia Cells Derived from Jurkat Line. Exp. Hematol. 2008, 36, 1278–1284. [Google Scholar] [CrossRef] [PubMed]
- Ri, M.; Iida, S.; Nakashima, T.; Miyazaki, H.; Mori, F.; Ito, A.; Inagaki, A.; Kusumoto, S.; Ishida, T.; Komatsu, H.; et al. Bortezomib-Resistant Myeloma Cell Lines: A Role for Mutated PSMB5 in Preventing the Accumulation of Unfolded Proteins and Fatal ER Stress. Leukemia 2010, 24, 1506–1512. [Google Scholar] [CrossRef]
- Matsunaga, T.; Tsuchimura, S.; Azuma, N.; Endo, S.; Ichihara, K.; Ikari, A. Caffeic Acid Phenethyl Ester Potentiates Gastric Cancer Cell Sensitivity to Doxorubicin and Cisplatin by Decreasing Proteasome Function. Anti-Cancer Drugs 2019, 30, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Haupt, Y.; Maya, R.; Kazaz, A.; Oren, M. Mdm2 Promotes the Rapid Degradation of P53. Nature 1997, 387, 296–299. [Google Scholar] [CrossRef]
- Zhu, Q.; Wani, G.; Yao, J.; Patnaik, S.; Wang, Q.E.; El-Mahdy, M.A.; Prætorius-Ibba, M.; Wani, A.A. The Ubiquitin–Proteasome System Regulates P53-Mediated Transcription at P21waf1 Promoter. Oncogene 2007, 26, 4199–4208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez, N.; Gallagher, M.; Lao, N.; Gallagher, C.; Clarke, C.; Doolan, P.; Aherne, S.; Blanco, A.; Meleady, P.; Clynes, M.; et al. MiR-7 Triggers Cell Cycle Arrest at the G1/S Transition by Targeting Multiple Genes Including Skp2 and Psme3. PLoS ONE 2013, 8, e65671. [Google Scholar] [CrossRef] [PubMed]
- Mesrian Tanha, H.; Mojtabavi Naeini, M.; Rahgozar, S.; Moafi, A.; Honardoost, M.A. Integrative Computational In-Depth Analysis of Dysregulated MiRNA-MRNA Interactions in Drug-Resistant Pediatric Acute Lymphoblastic Leukemia Cells: An Attempt to Obtain New Potential Gene-MiRNA Pathways Involved in Response to Treatment. Tumor Biol. 2016, 37, 7861–7872. [Google Scholar] [CrossRef]
- More, D.A.; Kumar, A. SRSF3: Newly Discovered Functions and Roles in Human Health and Diseases. Eur. J. Cell. Biol. 2020, 99, 151099. [Google Scholar] [CrossRef]
- Sen, S.; Langiewicz, M.; Jumaa, H.; Webster, N.J.G. Deletion of Serine/Arginine-Rich Splicing Factor 3 in Hepatocytes Predisposes to Hepatocellular Carcinoma in Mice. Hepatology 2015, 61, 171–183. [Google Scholar] [CrossRef] [Green Version]
- Kumar, D.; Das, M.; Sauceda, C.; Ellies, L.G.; Kuo, K.; Parwal, P.; Kaur, M.; Jih, L.; Bandyopadhyay, G.K.; Burton, D.; et al. Degradation of Splicing Factor SRSF3 Contributes to Progressive Liver Disease. J. Clin. Investig. 2019, 129, 4477–4491. [Google Scholar] [CrossRef] [PubMed]
- Torres, S.; García-Palmero, I.; Marín-Vicente, C.; Bartolomé, R.A.; Calviño, E.; Fernández-Aceñero, M.J.; Casal, J.I. Proteomic Characterization of Transcription and Splicing Factors Associated with a Metastatic Phenotype in Colorectal Cancer. J. Proteome Res. 2018, 17, 252–264. [Google Scholar] [CrossRef] [PubMed]
- McCubrey, J.A.; Abrams, S.L.; Fitzgerald, T.L.; Cocco, L.; Martelli, A.M.; Montalto, G.; Cervello, M.; Scalisi, A.; Candido, S.; Libra, M.; et al. Roles of Signaling Pathways in Drug Resistance, Cancer Initiating Cells and Cancer Progression and Metastasis. Adv. Biol. Regul. 2015, 57, 75–101. [Google Scholar] [CrossRef] [PubMed]
- You, K.S.; Yi, Y.W.; Kwak, S.J.; Seong, Y.S. Inhibition of RPTOR Overcomes Resistance to EGFR Inhibition in Triple-Negative Breast Cancer Cells. Int. J. Oncol. 2018, 52, 828–840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campos-Parra, A.D.; Padua-Bracho, A.; Pedroza-Torres, A.; Figueroa-González, G.; Fernández-Retana, J.; Millan-Catalan, O.; Peralta-Zaragoza, O.; Cantú de León, D.; Herrera, L.A.; Pérez-Plasencia, C. Comprehensive Transcriptome Analysis Identifies Pathways with Therapeutic Potential in Locally Advanced Cervical Cancer. Gynecol. Oncol. 2016, 143, 406–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Liang, S.; Yang, X.; Ji, Z.; Zhao, W.; Ye, X.; Rui, J. RNAi-Mediated RPL34 Knockdown Suppresses the Growth of Human Gastric Cancer Cells. Oncol. Rep. 2015, 34, 2267–2272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, S.; Zhao, J.; Fowdur, M.; Wang, K.; Jiang, T.; He, M. Highly Expressed Ribosomal Protein L34 Indicates Poor Prognosis in Osteosarcoma and Its Knockdown Suppresses Osteosarcoma Proliferation Probably through Translational Control. Sci. Rep. 2016, 6, 37690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.; Cui, J.; Yang, Y.; Liu, Z.; Yan, H.; Tang, C.; Wang, H.; Qin, H.; Li, X.; Li, J.; et al. Over-Expressed RPL34 Promotes Malignant Proliferation of Non-Small Cell Lung Cancer Cells. Gene 2016, 576, 421–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, H.; Li, J.; Jia, Y.; Wu, J.; Yuan, L.; Li, M.; Wei, J.; Xu, B. Silencing of Ribosomal Protein L34 (RPL34) Inhibits the Proliferation and Invasion of Esophageal Cancer Cells. Oncol. Res. 2017, 25, 1061–1068. [Google Scholar] [CrossRef] [PubMed]
- Rho, J.H.; Qin, S.; Wang, J.; Roehrl, M.H.A. Proteomic Expression Analysis of Surgical Human Colorectal Cancer Tissues: Up-Regulation of PSB7, PRDX1, and SRP9 and Hypoxic Adaptation in Cancer. J. Proteome Res. 2008, 7, 2959–2972. [Google Scholar] [CrossRef] [Green Version]
- Erdoğan, G.; Trabulus, D.C.; Talu, C.K.; Güven, M. Investigation of SRP9 Protein Expression in Breast Cancer. Mol. Biol. Rep. 2022, 49, 531–537. [Google Scholar] [CrossRef]
- Hsu, M.C.; Pan, M.R.; Chu, P.Y.; Tsai, Y.L.; Tsai, C.H.; Shan, Y.S.; Chen, L.T.; Hung, W.C. Protein Arginine Methyltransferase 3 Enhances Chemoresistance in Pancreatic Cancer by Methylating HnRNPA1 to Increase ABCG2 Expression. Cancers 2018, 11, 8. [Google Scholar] [CrossRef] [Green Version]
- Roy, R.; Durie, D.; Li, H.; Liu, B.Q.; Skehel, J.M.; Mauri, F.; Cuorvo, L.V.; Barbareschi, M.; Guo, L.; Holcik, M.; et al. HnRNPA1 Couples Nuclear Export and Translation of Specific MRNAs Downstream of FGF-2/S6K2 Signalling. Nucleic Acids Res. 2014, 42, 12483–12497. [Google Scholar] [CrossRef]
- Xu, Y.; Gao, X.D.; Lee, J.H.; Huang, H.; Tan, H.; Ahn, J.; Reinke, L.M.; Peter, M.E.; Feng, Y.; Gius, D.; et al. Cell Type-Restricted Activity of HnRNPM Promotes Breast Cancer Metastasis via Regulating Alternative Splicing. Genes Dev. 2014, 28, 1191–1203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harvey, S.E.; Xu, Y.; Lin, X.; Gao, X.D.; Qiu, Y.; Ahn, J.; Xiao, X.; Cheng, C. Coregulation of Alternative Splicing by HnRNPM and ESRP1 during EMT. RNA 2018, 24, 1326–1338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reviejo, M.; Soto, M.; Lozano, E.; Asensio, M.; Martínez-Augustin, O.; Sánchez de Medina, F.; Marin, J.J.G. Impact of Alternative Splicing on Mechanisms of Resistance to Anticancer Drugs. Biochem. Pharmacol. 2021, 193, 114810. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Xu, S.; Chen, Y.; Gao, Z.; Li, Y.; Hu, J.; Huang, X.; Zhang, Y.; Jiang, X.; Li, L.; et al. Depletion of Ars2 Inhibits Cell Proliferation and Leukemogenesis in Acute Myeloid Leukemia by Modulating the MiR-6734-3p/P27 Axis. Leukemia 2018, 33, 1090–1101. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Hu, X.; Li, Y.; Zhang, H.; Fu, R.; Liu, Y.; Hu, J.; Deng, Q.; Luo, Q.; Zhang, D.; et al. Ars2 Promotes Cell Proliferation and Tumorigenicity in Glioblastoma through Regulating MiR-6798-3p. Sci. Rep. 2018, 8, 15602. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Chen, C.; Xu, Z.; Scuoppo, C.; Rillahan, C.D.; Gao, J.; Spitzer, B.; Bosbach, B.; Kastenhuber, E.R.; Baslan, T.; et al. Deletions Linked to TP53 Loss Drive Cancer through P53-Independent Mechanisms. Nature 2016, 531, 471–475. [Google Scholar] [CrossRef] [Green Version]
- Xie, R.; Chen, X.; Cheng, L.; Huang, M.; Zhou, Q.; Zhang, J.; Chen, Y.; Peng, S.; Chen, Z.; Dong, W.; et al. NONO Inhibits Lymphatic Metastasis of Bladder Cancer via Alternative Splicing of SETMAR. Mol. Ther. 2021, 29, 291–307. [Google Scholar] [CrossRef]
- Cui, J.; Placzek, W.J. PTBP1 Modulation of MCL1 Expression Regulates Cellular Apoptosis Induced by Antitubulin Chemotherapeutics. Cell. Death Differ. 2016, 23, 1681–1690. [Google Scholar] [CrossRef] [Green Version]
- Barrón-Gallardo, C.A.; Garcia-Chagollán, M.; Morán-Mendoza, A.J.; Delgadillo-Cristerna, R.; Martínez-Silva, M.G.; Aguilar-Lemarroy, A.; Jave-Suárez, L.F. Transcriptomic Analysis of Breast Cancer Patients Sensitive and Resistant to Chemotherapy: Looking for Overall Survival and Drug Resistance Biomarkers. Technol. Cancer Res. Treat. 2022, 21, 15330338211068965. [Google Scholar] [CrossRef]
- RNA Expression Classifiers from a Model of Breast Epithelial Cell Organization to Predict Pathological Complete Response in Triple Negative Breast Cancer. Available online: https://www.fortunejournals.com/articles/rna-expression-classifiers-from-a-model-of-breast-epithelial-cell-organization-to-predict-pathological-complete-response-in-triple.html (accessed on 21 July 2022).
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-Seq Aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. FeatureCounts: An Efficient General Purpose Program for Assigning Sequence Reads to Genomic Features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langfelder, P.; Horvath, S. WGCNA: An R Package for Weighted Correlation Network Analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, I. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Chin, C.-H.; Chen, S.-H.; Wu, H.-H.; Ho, C.-W.; Ko, M.-T.; Lin, C.-Y. CytoHubba: Identifying Hub Objects and Sub-Networks from Complex Interactome. BMC Syst. Biol. 2014, 8 (Suppl 4), S11. [Google Scholar] [CrossRef] [Green Version]
- Ge, S.X.; Jung, D.; Jung, D.; Yao, R. ShinyGO: A Graphical Gene-Set Enrichment Tool for Animals and Plants. Bioinformatics 2020, 36, 2628–2629. [Google Scholar] [CrossRef]
- Freshour, S.L.; Kiwala, S.; Cotto, K.C.; Coffman, A.C.; McMichael, J.F.; Song, J.J.; Griffith, M.; Griffith, O.L.; Wagner, A.H. Integration of the Drug–Gene Interaction Database (DGIdb 4.0) with Open Crowdsource Efforts. Nucleic Acids Res. 2021, 49, D1144–D1151. [Google Scholar] [CrossRef]
- Caroli, J.; Sorrentino, G.; Forcato, M.; Del Sal, G.; Bicciato, S. GDA, a Web-Based Tool for Genomics and Drugs Integrated Analysis. Nucleic Acids Res. 2018, 46, W148–W156. [Google Scholar] [CrossRef] [Green Version]
- Davis, A.P.; Grondin, C.J.; Johnson, R.J.; Sciaky, D.; Wiegers, J.; Wiegers, T.C.; Mattingly, C.J. Comparative Toxicogenomics Database (CTD): Update 2021. Nucleic Acids Res. 2021, 49, D1138–D1143. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saha Detroja, T.; Detroja, R.; Mukherjee, S.; Samson, A.O. Identifying Hub Genes Associated with Neoadjuvant Chemotherapy Resistance in Breast Cancer and Potential Drug Repurposing for the Development of Precision Medicine. Int. J. Mol. Sci. 2022, 23, 12628. https://doi.org/10.3390/ijms232012628
Saha Detroja T, Detroja R, Mukherjee S, Samson AO. Identifying Hub Genes Associated with Neoadjuvant Chemotherapy Resistance in Breast Cancer and Potential Drug Repurposing for the Development of Precision Medicine. International Journal of Molecular Sciences. 2022; 23(20):12628. https://doi.org/10.3390/ijms232012628
Chicago/Turabian StyleSaha Detroja, Trishna, Rajesh Detroja, Sumit Mukherjee, and Abraham O. Samson. 2022. "Identifying Hub Genes Associated with Neoadjuvant Chemotherapy Resistance in Breast Cancer and Potential Drug Repurposing for the Development of Precision Medicine" International Journal of Molecular Sciences 23, no. 20: 12628. https://doi.org/10.3390/ijms232012628
APA StyleSaha Detroja, T., Detroja, R., Mukherjee, S., & Samson, A. O. (2022). Identifying Hub Genes Associated with Neoadjuvant Chemotherapy Resistance in Breast Cancer and Potential Drug Repurposing for the Development of Precision Medicine. International Journal of Molecular Sciences, 23(20), 12628. https://doi.org/10.3390/ijms232012628