

A Xanthohumol-Rich Hop Extract Diminishes Endotoxin-Induced Activation of TLR4 Signaling in Human Peripheral Blood Mononuclear Cells: A Study in Healthy Women
 , ,
, , 
Abstract
:1. Introduction
2. Results
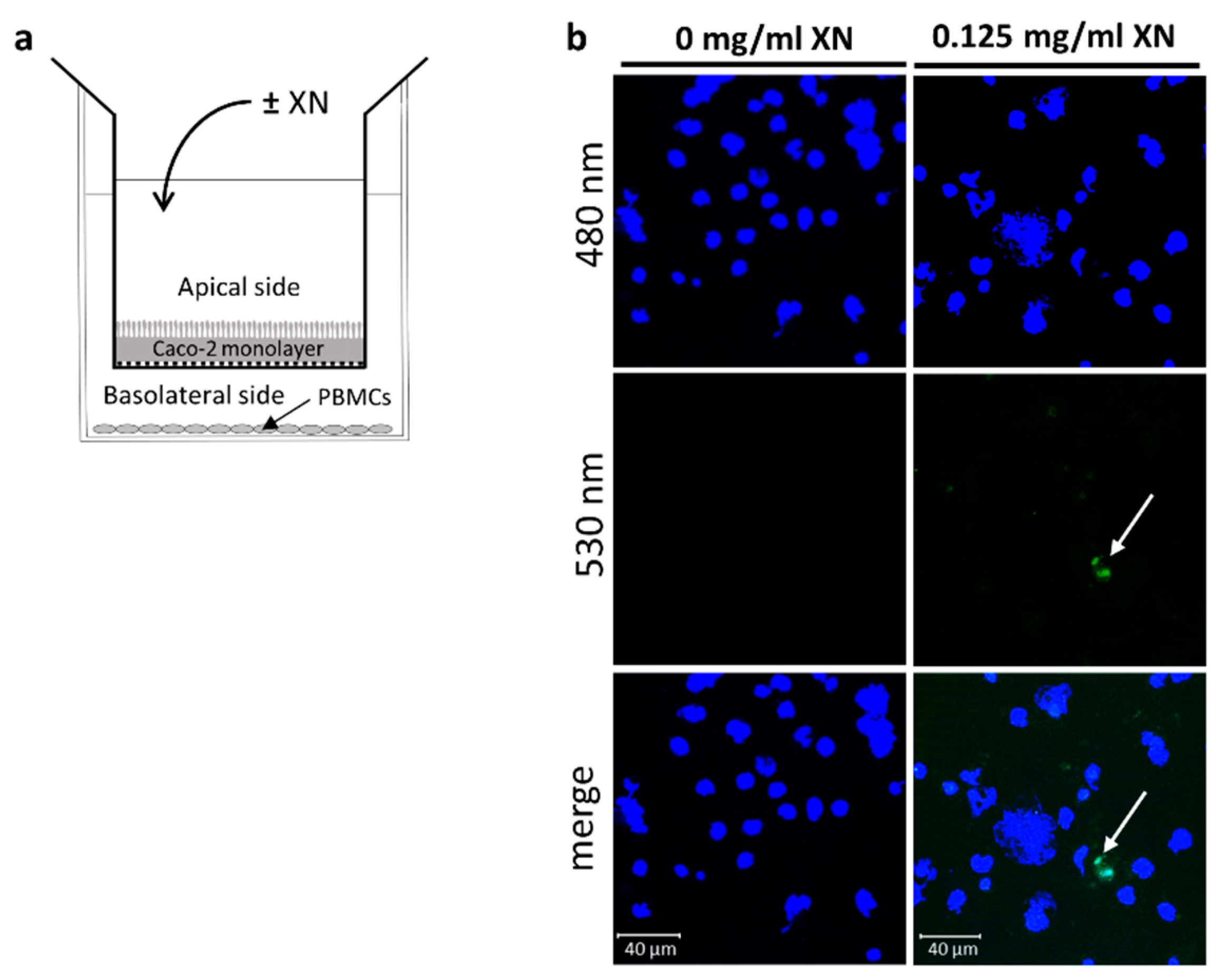
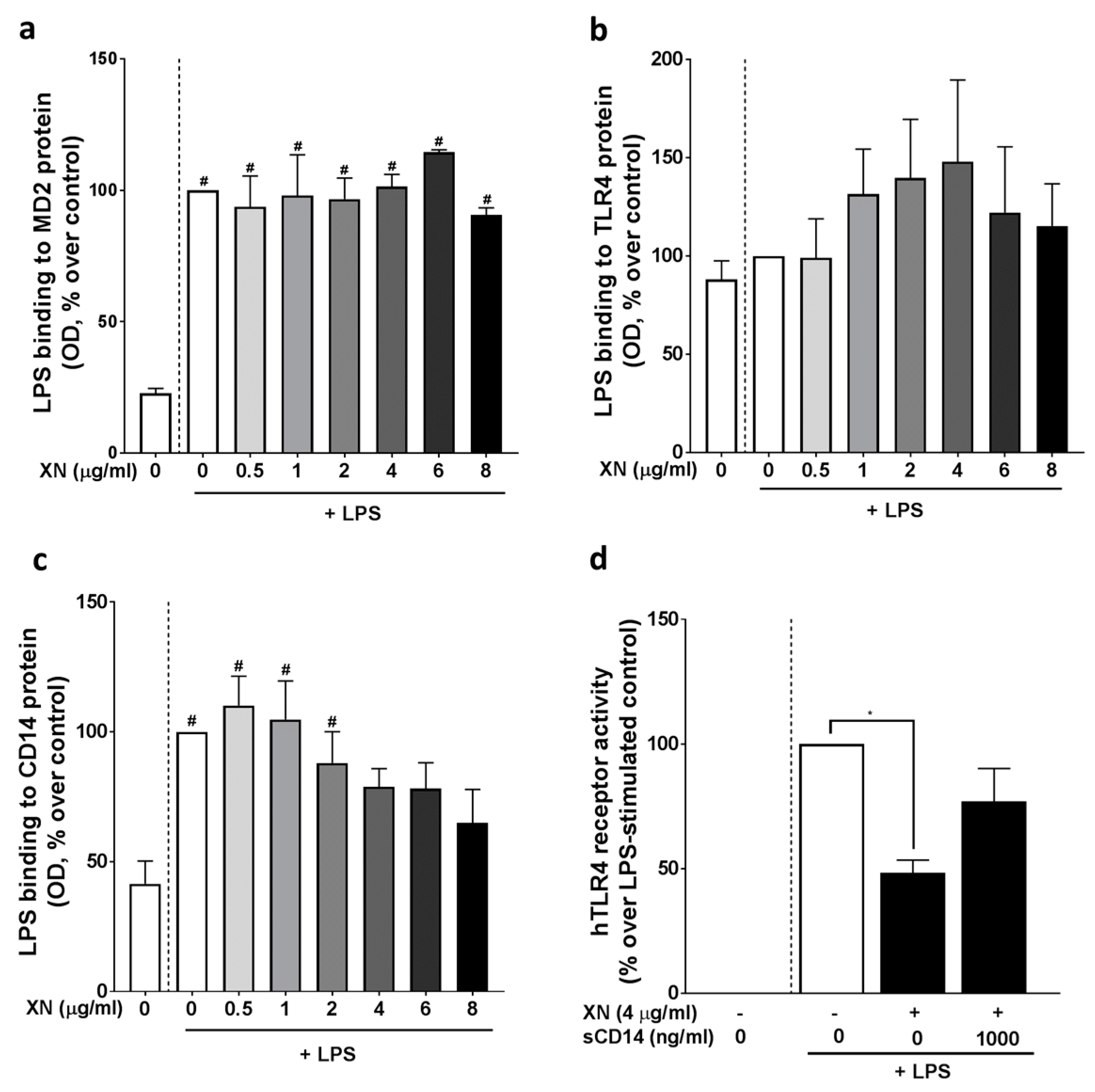
2.1. Co-Culture Cell Assay
2.2. Effect of the Oral Intake of XN on LPS-Induced Inflammation
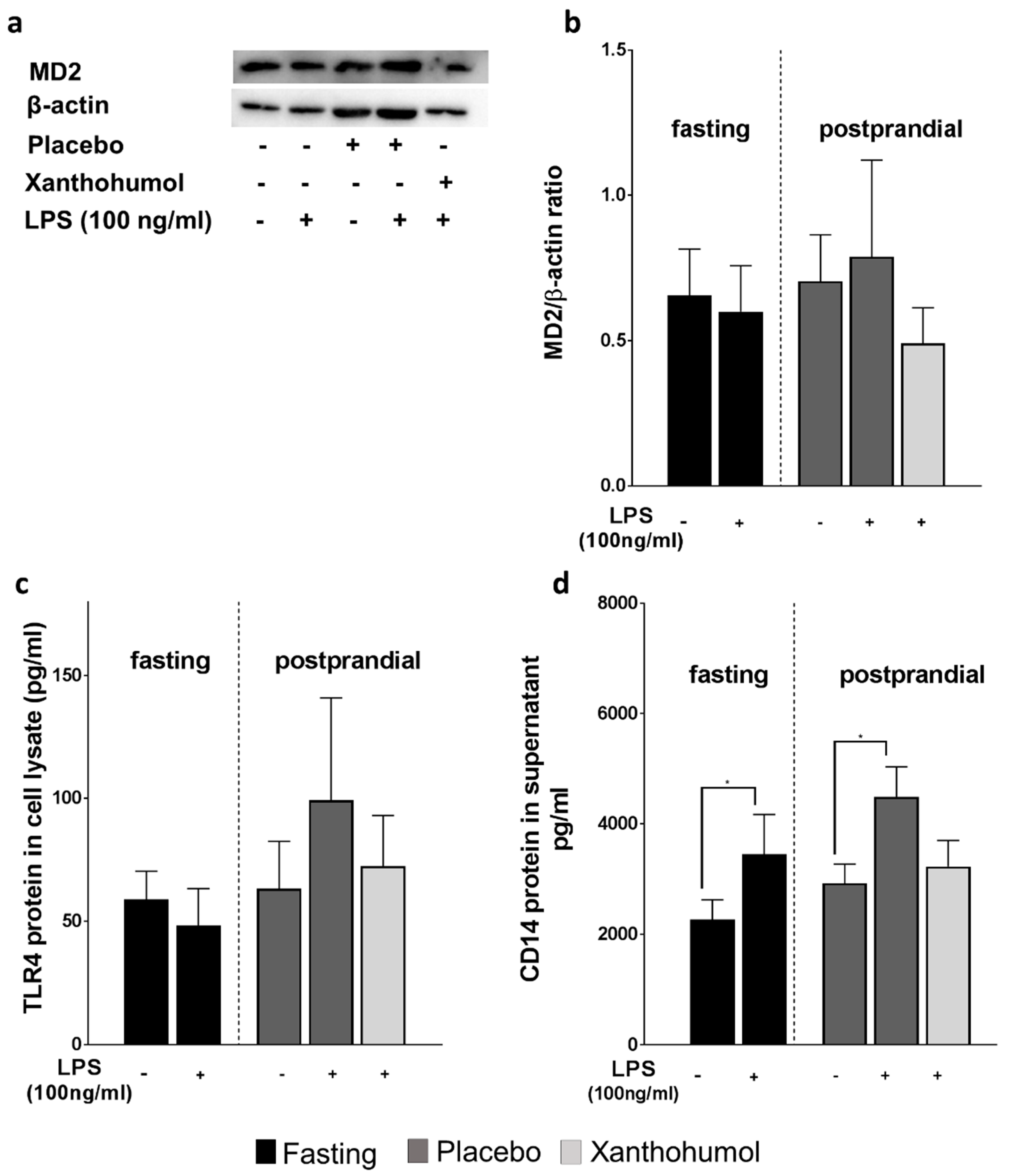
2.3. Effect of the Oral Intake of XN on MD-2, TLR4 and sCD14 Protein in PBMCs
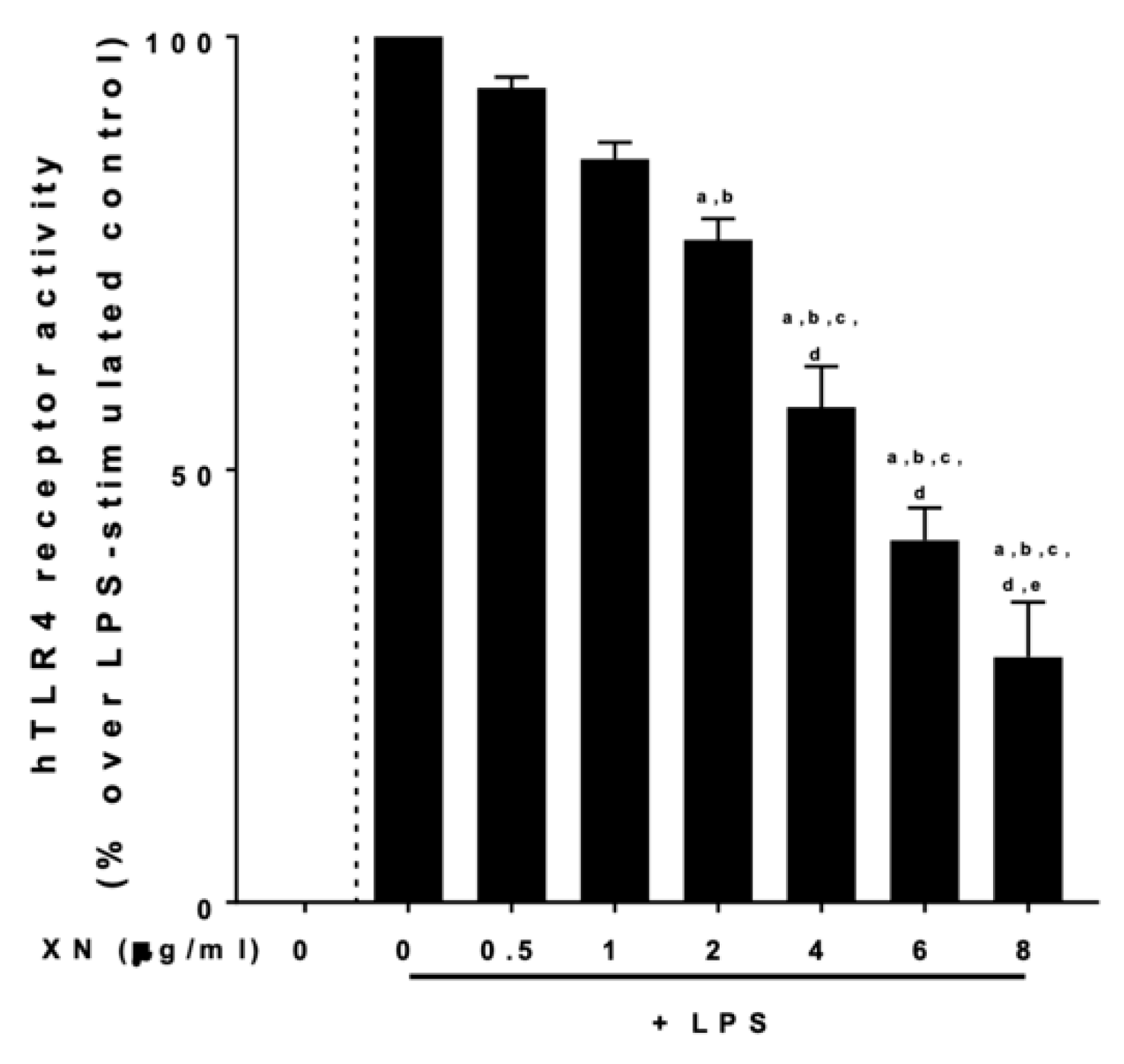
2.4. Effect of XN on the LPS-Dependent Activation of TLR4 and the Effect of sCD14 Herein
3. Discussion
Limitations
4. Materials and Methods
4.1. Co-Culture Cell Assay
4.2. Study Participants
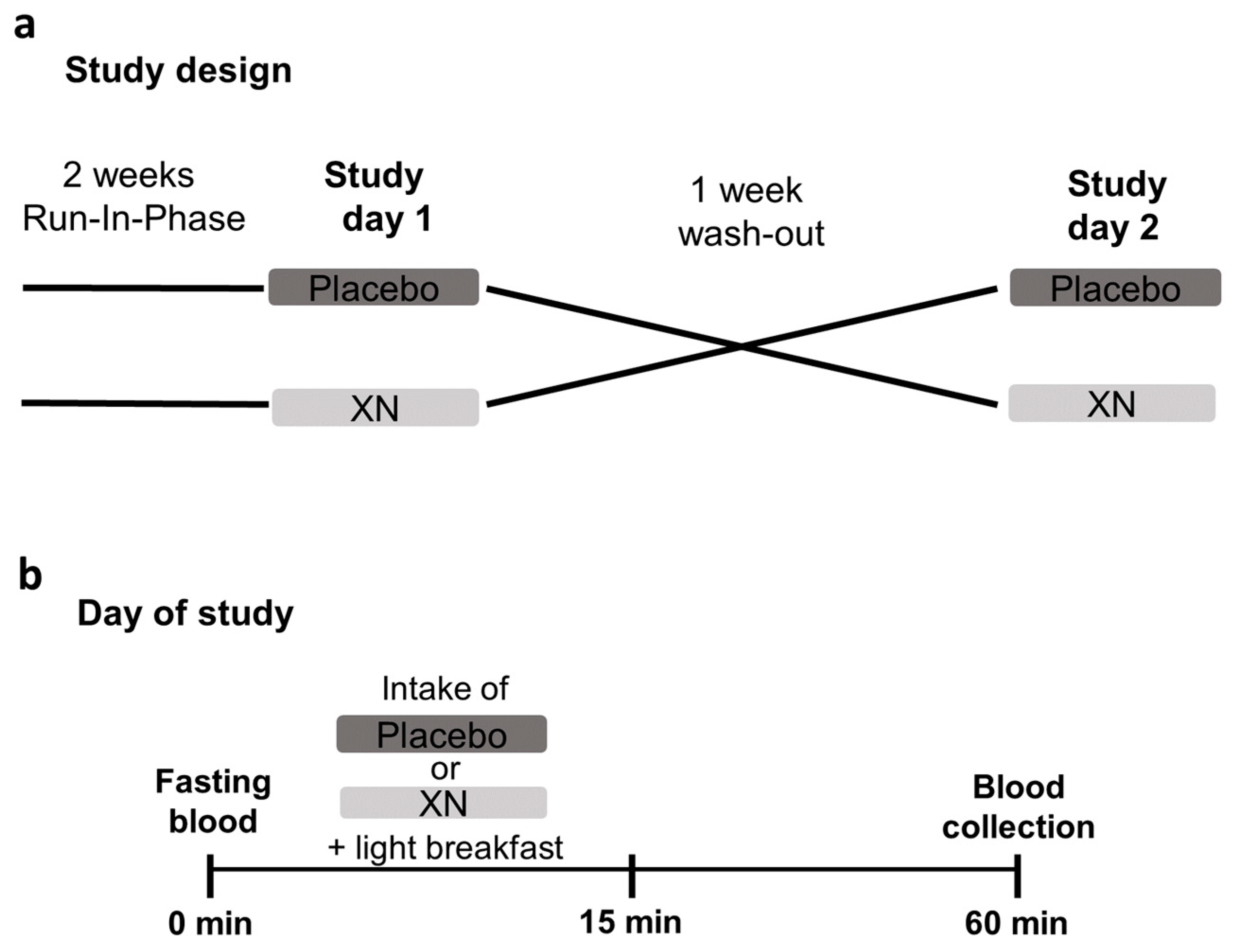
4.3. Intervention Study
4.4. Isolation and Culture of PBMCs
4.5. Cell Culture Experiments with hTLR4 Transfected HEK Cells Response
4.6. Western Blot
4.7. Enzyme-Linked Immunosorbent Assays (ELISA’s)
4.8. LPS Binding Assays
4.9. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Antimicrobial Resistance Collaborators. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- Massier, L.; Blüher, M.; Kovacs, P.; Chakaroun, R.M. Impaired Intestinal Barrier and Tissue Bacteria: Pathomechanisms for Metabolic Diseases. Front. Endocrinol. 2021, 12, 616506. [Google Scholar] [CrossRef] [PubMed]
- Hritz, I.; Mandrekar, P.; Velayudham, A.; Catalano, D.; Dolganiuc, A.; Kodys, K.; Kurt-Jones, E.; Szabo, G. The critical role of toll-like receptor (TLR) 4 in alcoholic liver disease is independent of the common TLR adapter MyD88. Hepatology 2008, 48, 1224–1231. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Zhang, X.; Li, Y.; Lopes-Virella, M.F.; Huanga, Y. TLR4 antagonist attenuates atherogenesis in LDL receptor-deficient mice with diet-induced type 2 diabetes. Immunobiology 2015, 220, 1246–1254. [Google Scholar] [CrossRef] [Green Version]
- Spruss, A.; Kanuri, G.; Wagnerberger, S.; Haub, S.; Bischoff, S.C.; Bergheim, I. Toll-like receptor 4 is involved in the development of fructose-induced hepatic steatosis in mice. Hepatology 2009, 50, 1094–1104. [Google Scholar] [CrossRef]
- Lu, Y.C.; Yeh, W.C.; Ohashi, P.S. LPS/TLR4 signal transduction pathway. Cytokine 2008, 42, 145–151. [Google Scholar] [CrossRef]
- Hege, M.; Jung, F.; Sellmann, C.; Jin, C.; Ziegenhardt, D.; Hellerbrand, C.; Bergheim, I. An iso-alpha-acid-rich extract from hops (Humulus lupulus) attenuates acute alcohol-induced liver steatosis in mice. Nutrition 2018, 45, 68–75. [Google Scholar] [CrossRef]
- Hsu, H.-Y.; Hua, K.-F.; Lin, C.-C.; Lin, C.-H.; Hsu, J.; Wong, C.-H. Extract of Reishi polysaccharides induces cytokine expression via TLR4-modulated protein kinase signaling pathways. J. Immunol. 2004, 173, 5989–5999. [Google Scholar] [CrossRef] [Green Version]
- Iniguez, A.B.; Zhu, M.J. Hop bioactive compounds in prevention of nutrition-related noncommunicable diseases. Crit. Rev. Food Sci. Nutr. 2020, 61, 1900–1913. [Google Scholar] [CrossRef]
- Legette, L.; Karnpracha, C.; Reed, R.L.; Choi, J.; Bobe, G.; Christensen, J.M.; Rodriguez-Proteau, R.; Purnell, J.Q.; Stevens, J.F. Human pharmacokinetics of xanthohumol, an antihyperglycemic flavonoid from hops. Mol. Nutr. Food Res. 2014, 58, 248–255. [Google Scholar] [CrossRef]
- Dorn, C.; Kraus, B.; Motyl, M.; Weiss, T.S.; Gehrig, M.; Schölmerich, J.; Heilmann, J.; Hellerbrand, C. Xanthohumol, a chalcon derived from hops, inhibits hepatic inflammation and fibrosis. Mol. Nutr. Food Res. 2010, 54 (Suppl. S2), S205–S213. [Google Scholar] [CrossRef]
- Miranda, C.L.; Elias, V.D.; Hay, J.J.; Choi, J.; Reed, R.L.; Stevens, J. Xanthohumol improves dysfunctional glucose and lipid metabolism in diet-induced obese C57BL/6J mice. Arch. Biochem. Biophys. 2016, 599, 22–30. [Google Scholar] [CrossRef] [Green Version]
- Cho, Y.-C.; Kim, H.J.; Kim, Y.-J.; Lee, K.Y.; Choi, H.J.; Lee, I.-S.; Kang, B.Y. Differential anti-inflammatory pathway by xanthohumol in IFN-gamma and LPS-activated macrophages. Int. Immunopharmacol. 2008, 8, 567–573. [Google Scholar] [CrossRef]
- Jung, F.; Staltner, R.; Tahir, A.; Baumann, A.; Burger, K.; Halilbasic, E.; Hellerbrand, C.; Bergheim, I. Oral intake of xanthohumol attenuates lipoteichoic acid-induced inflammatory response in human PBMCs. Eur. J. Nutr. 2022, 61. [Google Scholar] [CrossRef]
- Peluso, M.R.; Miranda, C.L.; Hobbs, D.J.; Proteau, R.R.; Stevens, J.F. Xanthohumol and related prenylated flavonoids inhibit inflammatory cytokine production in LPS-activated THP-1 monocytes: Structure-activity relationships and in silico binding to myeloid differentiation protein-2 (MD-2). Planta Med. 2010, 76, 1536–1543. [Google Scholar] [CrossRef]
- Chen, G.; Xiao, B.; Chen, L.; Bai, B.; Zhang, Y.; Xu, Z.; Fu, L.; Liu, Z.; Li, X.; Zhao, Y.; et al. Discovery of new MD2-targeted anti-inflammatory compounds for the treatment of sepsis and acute lung injury. Eur. J. Med. Chem. 2017, 139, 726–740. [Google Scholar] [CrossRef]
- Langley, B.O.; Ryan, J.J.; Hanes, D.; Phipps, J.; Stack, E.; Metz, T.O.; Stevens, J.F.; Bradley, R. Xanthohumol Microbiome and Signature in Healthy Adults (the XMaS Trial): Safety and Tolerability Results of a Phase I Triple-Masked, Placebo-Controlled Clinical Trial. Mol. Nutr. Food Res. 2021, 65, e2001170. [Google Scholar] [CrossRef]
- Pichler, C.; Ferk, F.; Al-Serori, H.; Huber, W.; Jäger, W.; Waldherr, M.; Mišík, M.; Kundi, M.; Nersesyan, A.; Herbacek, I.; et al. Xanthohumol Prevents DNA Damage by Dietary Carcinogens: Results of a Human Intervention Trial. Cancer Prev. Res. 2017, 10, 153–160. [Google Scholar] [CrossRef] [Green Version]
- Stevens, J.F.; Page, J.E. Xanthohumol and related prenylflavonoids from hops and beer: To your good health! Phytochemistry 2004, 65, 1317–1330. [Google Scholar] [CrossRef]
- Stevens, J.F.; Taylor, A.W.; Clawson, J.E.; Deinzer, M.L. Fate of xanthohumol and related prenylflavonoids from hops to beer. J. Agric. Food Chem. 1999, 4, 2421–2428. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, Y.; Zhang, W.; Liu, H.; Sun, C.; Zhang, B.; Bai, B.; Wu, D.; Xiao, Z.; Lum, H.; et al. Inhibition of myeloid differentiation factor 2 by baicalein protects against acute lung injury. Phytomedicine 2019, 63, 152997. [Google Scholar] [CrossRef]
- Ventola, C.L. The antibiotic resistance crisis: Part 1: Causes and threats. Pharmacol. Ther. 2015, 40, 277–283. [Google Scholar]
- Ferk, F.; Mišík, M.; Nersesyan, A.; Pichler, C.; Jäger, W.; Szekeres, T.; Marculescu, R.; Poulsen, H.E.; Henriksen, T.; Bono, R.; et al. Impact of xanthohumol (a prenylated flavonoid from hops) on DNA stability and other health-related biochemical parameters: Results of human intervention trials. Mol. Nutr. Food Res. 2016, 60, 773–786. [Google Scholar] [CrossRef]
- Chen, X.; Li, Z.; Hong, H.; Wang, N.; Chen, J.; Lu, S.; Zhang, H.; Zhang, X.; Bei, C. Xanthohumol suppresses inflammation in chondrocytes and ameliorates osteoarthritis in mice. Biomed. Pharmacother. 2021, 137, 111238. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhang, R.; Zheng, T.; Chen, Z.; Ji, G.; Peng, F.; Wang, W. Xanthohumol Attenuated Inflammation and ECM Degradation by Mediating HO-1/C/EBPbeta Pathway in Osteoarthritis Chondrocytes. Front. Pharmacol. 2021, 12, 680585. [Google Scholar] [CrossRef] [PubMed]
- Bouchard-Boivin, F.; Désy, O.; Béland, S.; Houde, I.; De Serres, S.A. TNF-alpha Production by Monocytes Stimulated with Epstein-Barr Virus-Peptides as a Marker of Immunosuppression-Related Adverse Events in Kidney Transplant Recipients. Kidney Int. Rep. 2019, 4, 1446–1453. [Google Scholar] [CrossRef] [Green Version]
- Ciesielska, A.; Matyjek, M.; Kwiatkowska, K. TLR4 and CD14 trafficking and its influence on LPS-induced pro-inflammatory signaling. Cell Mol. Life Sci. 2021, 78, 1233–1261. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Correia, J.; Soldau, K.; Christen, U.; Tobias, P.S.; Ulevitch, R.J. Lipopolysaccharide is in close proximity to each of the proteins in its membrane receptor complex. transfer from CD14 to TLR4 and MD-2. J. Biol. Chem. 2001, 276, 21129–21135. [Google Scholar] [CrossRef] [Green Version]
- Da Silva Correia, J.; Ulevitch, R.J. MD-2 and TLR4 N-linked glycosylations are important for a functional lipopolysaccharide receptor. J. Biol. Chem. 2002, 277, 1845–1854. [Google Scholar] [CrossRef] [Green Version]
- Bazil, V.; Strominger, J.L. Shedding as a mechanism of down-modulation of CD14 on stimulated human monocytes. J. Immunol. 1991, 147, 1567–1574. [Google Scholar]
- Rokita, E.; Menzel, E.J. Characteristics of CD14 shedding from human monocytes. Evidence for the competition of soluble CD14 (sCD14) with CD14 receptors for lipopolysaccharide (LPS) binding. APMIS 1997, 105, 510–518. [Google Scholar] [CrossRef] [PubMed]
- Dresel, M.; Dunkel, A.; Hofmann, T. Sensomics analysis of key bitter compounds in the hard resin of hops (Humulus lupulus L.) and their contribution to the bitter profile of Pilsner-type beer. J. Agric. Food Chem. 2015, 63, 3402–3418. [Google Scholar] [CrossRef] [PubMed]
- Menck, K.; Behme, D.; Pantke, M.; Reiling, N.; Binder, C.; Pukrop, T.; Klemm, F. Isolation of human monocytes by double gradient centrifugation and their differentiation to macrophages in teflon-coated cell culture bags. J. Vis. Exp. 2014, 91, e51554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motyl, M.; Kraus, B.; Heilmann, J. Pitfalls in cell culture work with xanthohumol. Pharmazie 2012, 67, 91–94. [Google Scholar]
- Wolff, H.; Motyl, M.; Hellerbrand, C.; Heilmann, J.; Kraus, B. Xanthohumol uptake and intracellular kinetics in hepatocytes, hepatic stellate cells, and intestinal cells. J. Agric. Food Chem. 2011, 59, 12893–12901. [Google Scholar] [CrossRef]
- Capó, X.; Martorell, M.; Sureda, A.; Batle, J.M.; Tur, J.A.; Pons, A. Docosahexaenoic diet supplementation, exercise and temperature affect cytokine production by lipopolysaccharide-stimulated mononuclear cells. J. Physiol. Biochem. 2016, 72, 421–434. [Google Scholar] [CrossRef]
- Brandt, A.; Jin, C.J.; Nolte, K.; Sellmann, C.; Engstler, A.J.; Bergheim, I. Short-Term Intake of a Fructose-, Fat- and Cholesterol-Rich Diet Causes Hepatic Steatosis in Mice: Effect of Antibiotic Treatment. Nutrients 2017, 9, 1013. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Wu, J.; Ying, S.; Chen, G.; Wu, B.; Xu, T.; Liang, G. Discovery of new MD2 inhibitor from chalcone derivatives with anti-inflammatory effects in LPS-induced acute lung injury. Sci. Rep. 2016, 6, 25130. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Healthy Participants |
|---|---|
| Sex (m/f) | 0/12 |
| Age (years) | 26.1 ± 1.1 |
| Body weight (kg) | 60.9 ± 2.1 |
| Height (m) | 1.66 ± 0.02 |
| BMI (kg/m2) | 22.1 ± 0.5 |
| Blood pressure | |
| Systolic (mmHg) | 126.6 ± 7.57 |
| Diastolic (mmHg) | 78.3 ± 3.6 |
| Fasting glucose (mg/dL) | 90.1 ± 2.2 |
| Uric acid (mg/dL) | 4.1 ± 0.2 |
| AST (U/L) | 29.8 ± 8.7 |
| ALT (U/L) | 33.7 ± 16.6 |
| γ-GT (U/L) | 12.2 ± 1.2 |
| Cholesterol (mg/dL) | 171.8 ± 8.4 |
| HDL cholesterol (mg/dL) | 66.4 ± 4.8 |
| LDL cholesterol (mg/dL) | 91.6 ± 6.1 |
| Triglycerides (mg/dL) | 77.6 ± 9.2 |
| CRP (mg/dL) | 0.03 ± 0.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jung, F.; Staltner, R.; Baumann, A.; Burger, K.; Halilbasic, E.; Hellerbrand, C.; Bergheim, I. A Xanthohumol-Rich Hop Extract Diminishes Endotoxin-Induced Activation of TLR4 Signaling in Human Peripheral Blood Mononuclear Cells: A Study in Healthy Women. Int. J. Mol. Sci. 2022, 23, 12702. https://doi.org/10.3390/ijms232012702
Jung F, Staltner R, Baumann A, Burger K, Halilbasic E, Hellerbrand C, Bergheim I. A Xanthohumol-Rich Hop Extract Diminishes Endotoxin-Induced Activation of TLR4 Signaling in Human Peripheral Blood Mononuclear Cells: A Study in Healthy Women. International Journal of Molecular Sciences. 2022; 23(20):12702. https://doi.org/10.3390/ijms232012702
Chicago/Turabian StyleJung, Finn, Raphaela Staltner, Anja Baumann, Katharina Burger, Emina Halilbasic, Claus Hellerbrand, and Ina Bergheim. 2022. "A Xanthohumol-Rich Hop Extract Diminishes Endotoxin-Induced Activation of TLR4 Signaling in Human Peripheral Blood Mononuclear Cells: A Study in Healthy Women" International Journal of Molecular Sciences 23, no. 20: 12702. https://doi.org/10.3390/ijms232012702
APA StyleJung, F., Staltner, R., Baumann, A., Burger, K., Halilbasic, E., Hellerbrand, C., & Bergheim, I. (2022). A Xanthohumol-Rich Hop Extract Diminishes Endotoxin-Induced Activation of TLR4 Signaling in Human Peripheral Blood Mononuclear Cells: A Study in Healthy Women. International Journal of Molecular Sciences, 23(20), 12702. https://doi.org/10.3390/ijms232012702