


On the Need to Distinguish between Insulin-Normal and Insulin-Resistant Patients in Testosterone Therapy

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
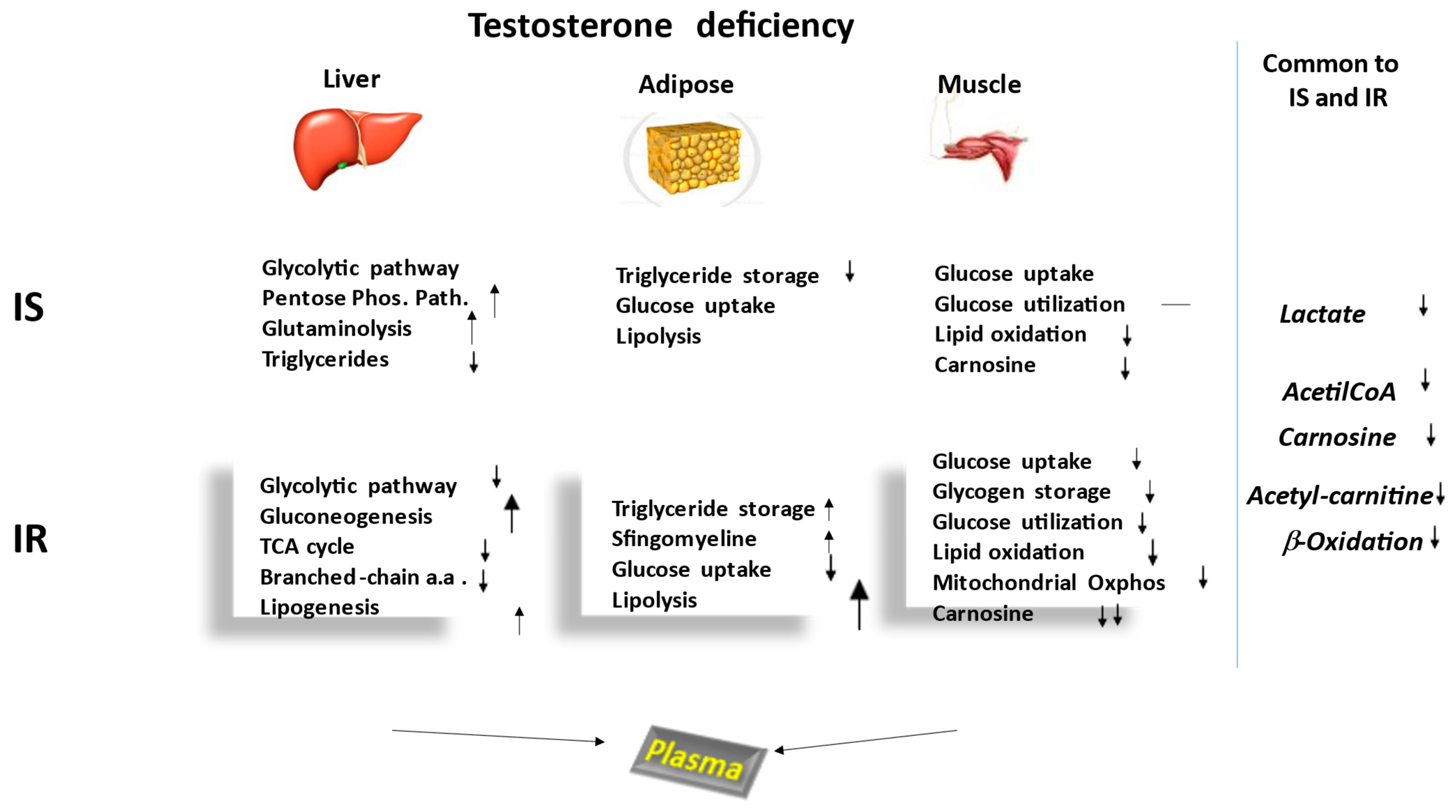
2. Metabolomic Comparison between IS and IR Hypogonadism
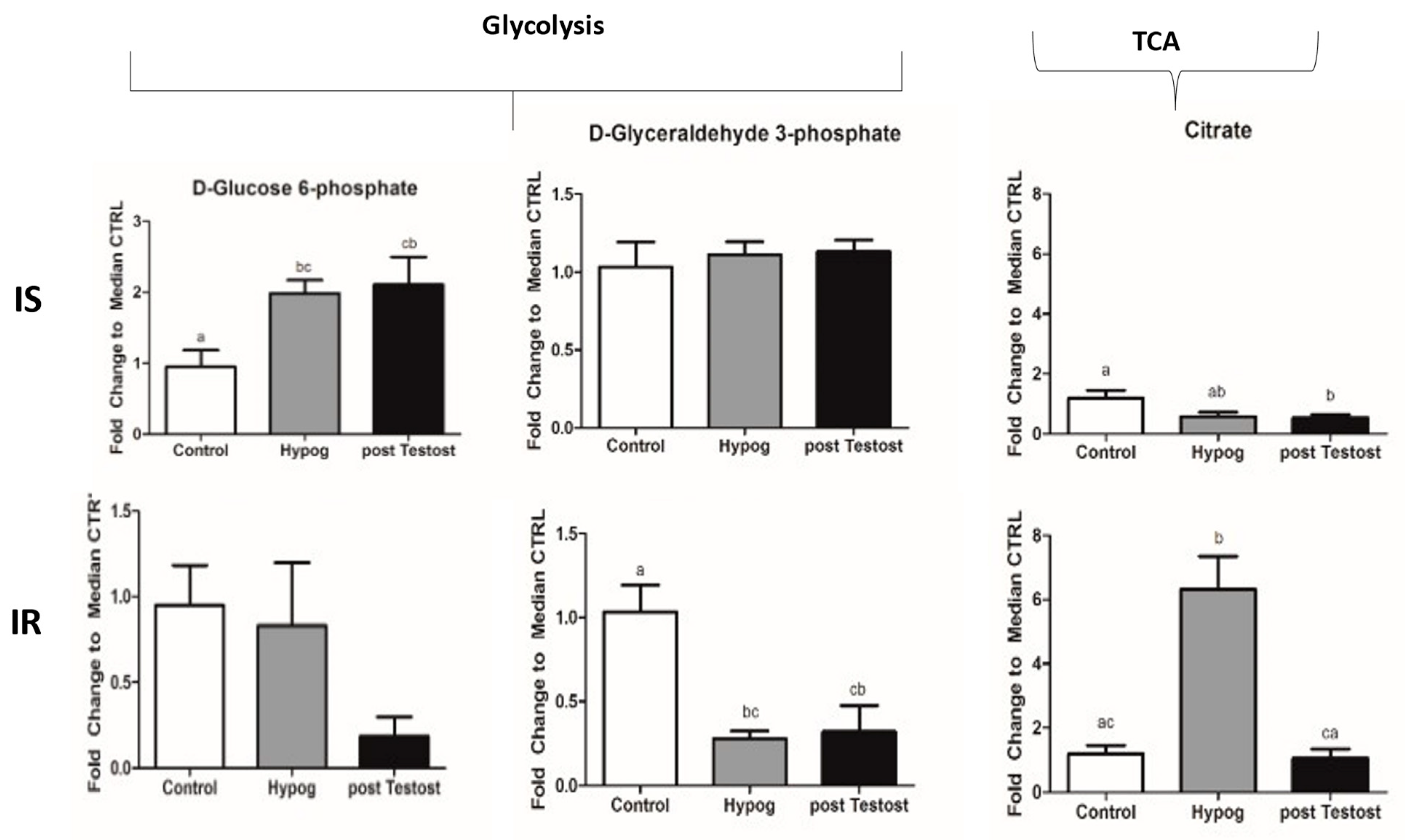
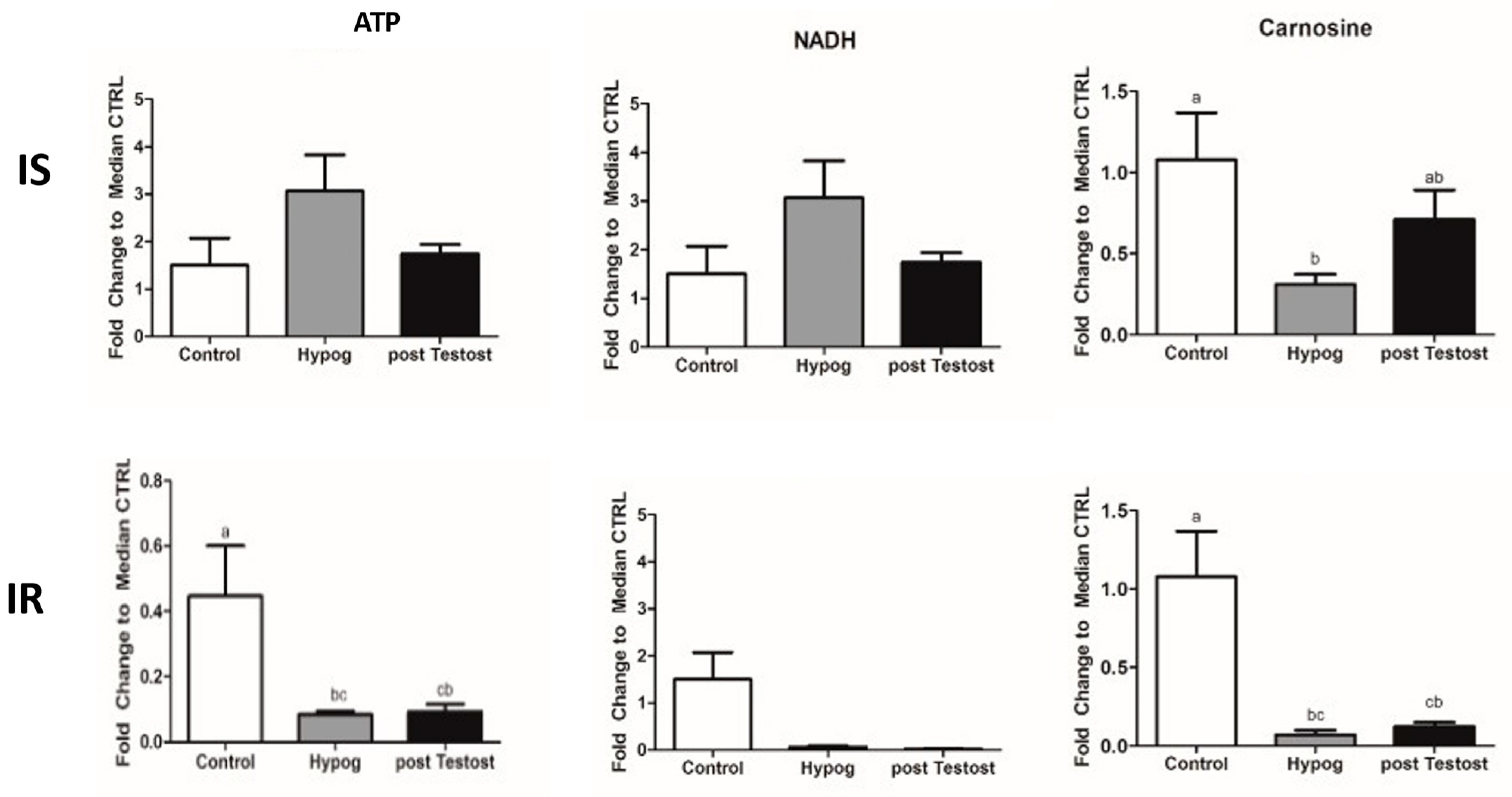
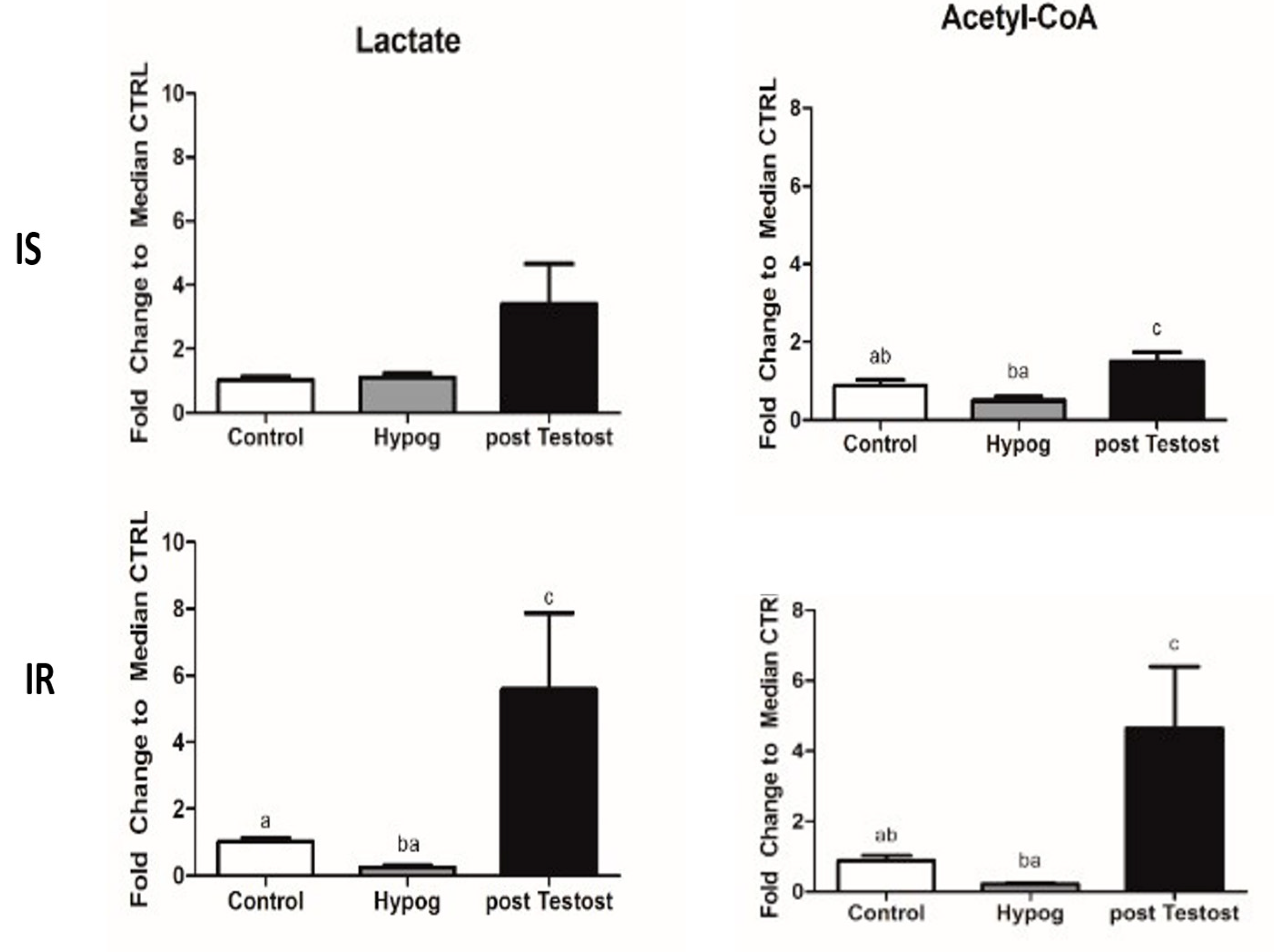
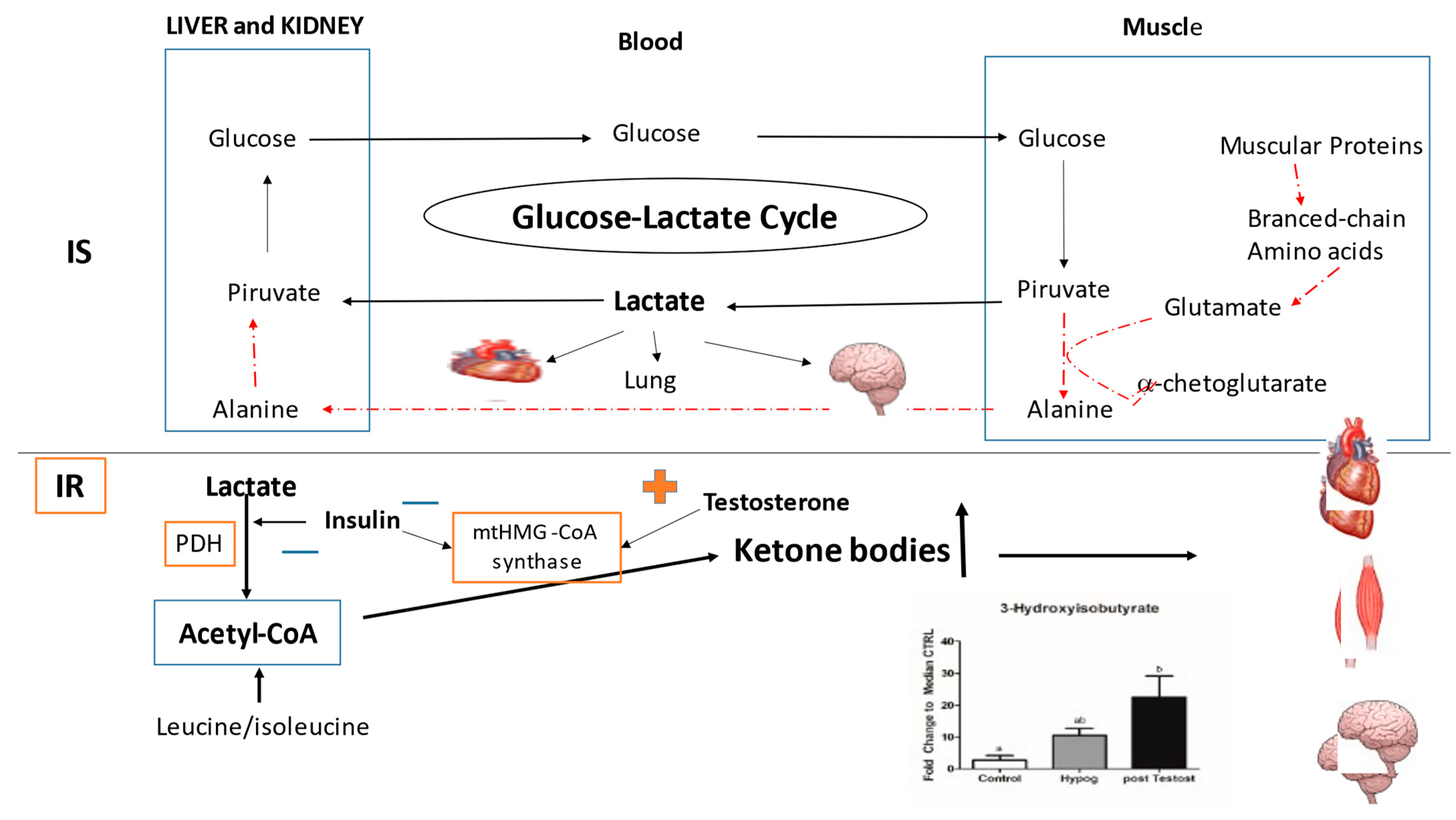
2.1. Carbohydrate Metabolism
2.2. Lipid Metabolism
2.3. Amino Acid Metabolism
2.4. Other Metabolisms
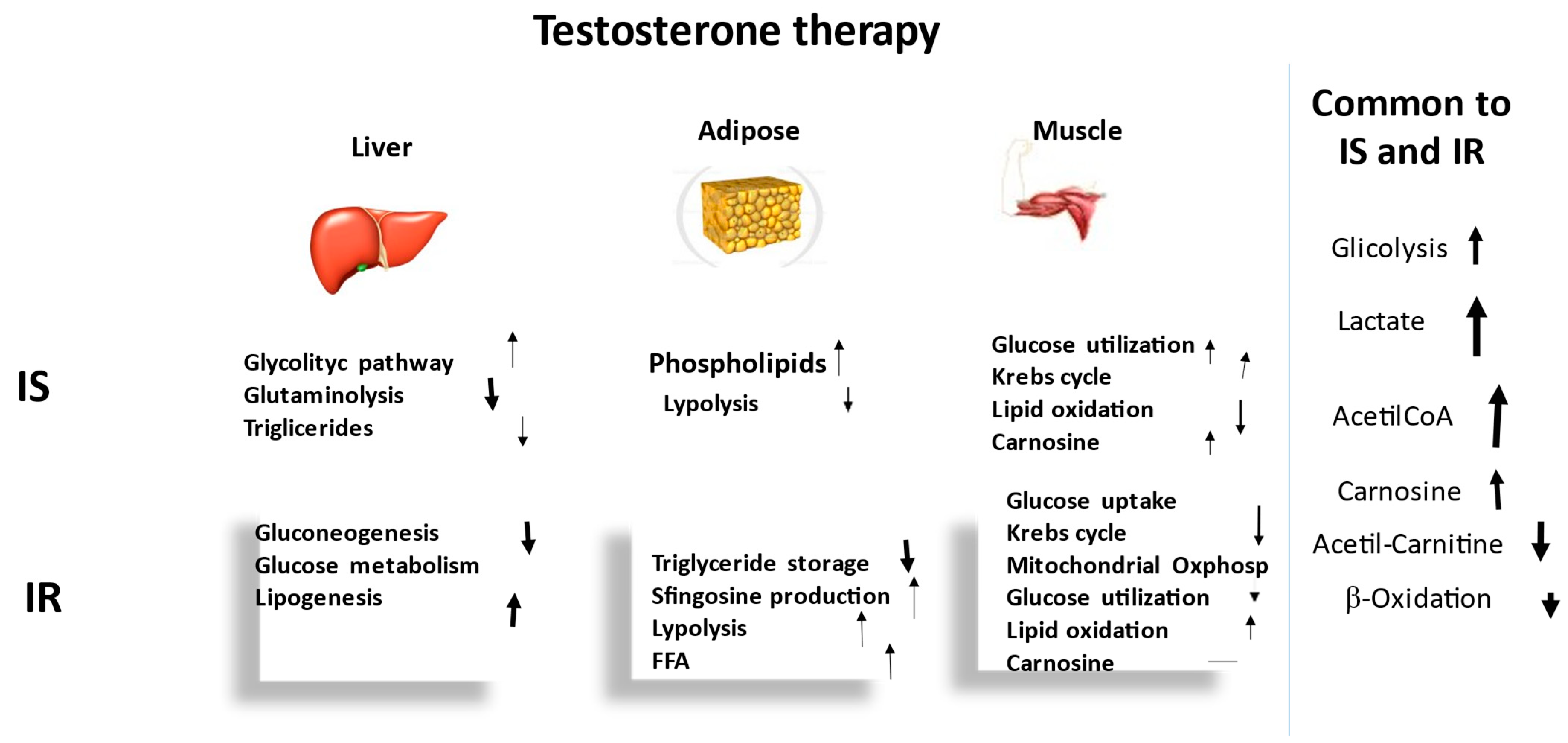
3. Metabolomic Comparison between IS and IR Hypogonadism after TRT
3.1. Carbohydrate Metabolism
3.2. Lipid Metabolism
3.3. Amino Acid Metabolism
4. Discussion
5. Conclusions
6. Future Directions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vakkat, M.; Jones, T.H. Testosterone and the metabolic syndrome. Ther. Adv. Endocrinol. Metab. 2010, 5, 207–223. [Google Scholar]
- Araujo, A.B.; O’Donnell, A.B.; Brambilla, D.J.; Simpson, W.B.; Longcope, C.; Matsumoto, A.M.; Ckinlay, J.B.M. Prevalence and incidence of androgen deficiency in middle-aged and older men: Estimates from the Massachusetts Male Aging Study. J. Clin. Endocrinol. Metab. 2004, 89, 5920–5926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lunenfeld, B. Testosterone deficiency and the metabolic syndrome. Aging Male 2007, 10, 53–56. [Google Scholar] [CrossRef]
- Mulligan, T.; Frick, M.F.; Zuraw, Q.C.; Stemhagen, A.; Mcwhirter, C. Prevalence of hypogonadism in males aged at least 45 years: The HIM study. Int. J. Clin. Pract. 2006, 60, 762–769. [Google Scholar] [CrossRef] [Green Version]
- Morgentaler, A.; Miner, M.M.; Caliber, M.; Guay, A.T.; Khera, M.; Traish, A.M. Testosterone Therapy and Cardiovascular Risk: Advances and Controversies. Mayo Clin. Proc. 2015, 90, 224–251. [Google Scholar] [CrossRef] [Green Version]
- Behre, H.M.; Tammela, T.L.J.; Arver, S.; Tolrá, J.R.; Bonifacio, V.; Lamche, M.; Kelly, J.; Hiemeyer, F.; Giltay, E.J.; Gooren, L.J.; et al. A randomized, double-blind, placebo-controlled trial of testosterone gel on body composition and health-related quality-of-life in men with hypogonadal to low-normal levels of serum testosterone and symptoms of androgen deficiency over 6 months with 12 months open-label follow-up. Aging Male 2012, 15, 198–207. [Google Scholar] [CrossRef]
- Yassin, A.A.; Saad, F.; Gooren, L.J. Metabolic syndrome, testosterone deficiency and erectile dysfunction never come alone. Andrologia 2008, 40, 259–264. [Google Scholar] [CrossRef]
- Hamilton, E.J.; Gianatti, E.; Strauss, B.J.; Wentworth, J.; Lim-Joon, D.; Bolton, D.; Zajac, J.D.; Grossmann, M. Increase in visceral and subcutaneous abdominal fat in men with prostate cancer treated with androgen deprivation therapy. Clin. Endocrinol. 2011, 74, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Gianatti, E.J.; Dupuis, P.; Hoermann, R.; Strauss, B.J.; Wentworth, J.M.; Zajac, J.D.; Grossmann, M. Effect of Testosterone Treatment on Glucose Metabolism in Men with Type 2 Diabetes: A Randomized Controlled Trial. Diabetes Care 2014, 37, 2098–2107. [Google Scholar] [CrossRef] [Green Version]
- Jones, T.H.; Arver, S.; Behre, H.M.; Buvat, J.; Meuleman, E.; Moncada, I.; Morales, A.M.; Volterrani, M.; Yellowlees, A.; Howell, J.D.; et al. Testosterone Replacement in Hypogonadal Men with Type 2 Diabetes and/or Metabolic Syndrome (the TIMES2 Study). Diabetes Care 2011, 34, 828–837. [Google Scholar] [CrossRef] [Green Version]
- Hackett, G.; Cole, N.; Bhartia, M.; Kennedy, D.; Raju, J.; Wilkinson, P.; The BLAST Study Group. Testosterone Replacement Therapy Improves Metabolic Parameters in Hypogonadal Men with Type 2 Diabetes but Not in Men with Coexisting Depression: The BLAST Study. J. Sex. Med. 2014, 11, 840–856. [Google Scholar] [CrossRef] [PubMed]
- Tsai, E.C.; Matsumoto, A.M.; Fujimoto, W.Y.; Boyko, E.J. Association of bioavailable, free, and total testosterone with insulin resistance influence of sex hormone-binding globulin and body fat. Diabetes Care 2004, 27, 861–868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhindsa, S.; Ghanim, H.; Batra, M.; Kuhadiya, N.D.; Abuaysheh, S.; Sandhu, S.; Green, K.; Makdissi, A.; Hejna, J.; Chaudhuri, A.; et al. Insulin Resistance and Inflammation in Hypogonadotropic Hypogonadism and Their Reduction after Testosterone Replacement in Men with Type 2 Diabetes. Diabetes Care 2016, 39, 82–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitteloud, N.; Hardin, M.; Dwyer, A.A.; Valassi, E.; Yialamas, M.; Elahi, D.; Hayes, F.J. Increasing Insulin Resistance Is Associated with a Decrease in Leydig Cell Testosterone Secretion in Men. J. Clin. Endocrinol. Metab. 2005, 90, 2636–2641. [Google Scholar] [CrossRef] [Green Version]
- Corona, G.; Mannucci, E.; Petrone, L.; Ricca, V.; Balercia, G.; Mansani, R.; Chiarini, V.; Giommi, R.; Forti, G.; Maggi, M. Association of hypogonadism and type II diabetes in men attending an outpatient erectile dysfunction clinic. Int. J. Impot. Res. 2005, 18, 190–197. [Google Scholar] [CrossRef] [Green Version]
- Stellato, R.K.; A Feldman, H.; Hamdy, O.; Horton, E.S.; McKinlay, J.B. Testosterone, sex hormone-binding globulin, and the development of type 2 diabetes in middle-aged men: Prospective results from the Massachusetts male aging study. Diabetes Care 2000, 23, 490–494. [Google Scholar] [CrossRef] [Green Version]
- Haffner, S.M.; Laakso, M.; Miettinen, H.; Mykkänen, L.; Karhapää, P.; Rainwater, D.L. Low levels of sex hormone-binding globulin and testosterone are associated with smaller, denser low density lipoprotein in normoglycemic men. J. Clin. Endocrinol. Metab. 1996, 81, 3697–3701. [Google Scholar] [CrossRef] [Green Version]
- Ottarsdottir, K.; Nilsson, A.G.; Hellgren, M.; Lindblad, U.; Daka, B. The association between serum testosterone and insulin resistance: A longitudinal study. Endocr. Connect. 2018, 7, 1491–1500. [Google Scholar] [CrossRef]
- Laaksonen, D.E.; Niskanen, L.; Punnonen, K.; Nyyssönen, K.; Tuomainen, T.P.; Valkonen, V.P.; Salonen, R.; Salonen, J.T. Testosterone and Sex Hormone–Binding Globulin Predict the Metabolic Syndrome and Diabetes in Middle-Aged Men. Diabetes Care 2004, 27, 1036–1041. [Google Scholar] [CrossRef] [Green Version]
- Selvin, E.; Feinleib, M.; Zhang, L.; Rohrmann, S.; Rifai, N.; Nelson, W.G.; Dobs, A.; Basaria, S.; Golden, S.H.; Platz, E.A. Androgens and Diabetes in Men: Results from the Third National Health and Nutrition Examination Survey (NHANES III). Diabetes Care 2007, 30, 234–238. [Google Scholar] [CrossRef] [Green Version]
- Kupelian, V.; Page, S.T.; Araujo, A.B.; Travison, T.G.; Bremner, W.J.; McKinlay, J.B. Low Sex Hormone-Binding Globulin, Total Testosterone, and Symptomatic Androgen Deficiency Are Associated with Development of the Metabolic Syndrome in Nonobese Men. J. Clin. Endocrinol. Metab. 2006, 91, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Haider, K.S.; Haider, A.; Saad, F.; Doros, G.; Hanefeld, M.; Dhindsa, S.; Dandona, P.; Traish, A. Remission of type 2 diabetes following long-term treatment with injectable testosterone undecanoate in patients with hypogonadism and type 2 diabetes: 11-year data from a real-world registry study. Diabetes Obes. Metab. 2020, 22, 2055–2068. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, D.; Goodwin, E.; Channer, K.S.; Jones, T.H. Testosterone replacement therapy improves insulin resistance, glycaemic control, visceral adiposity and hypercholesterolaemia in hypogonadal men with type 2 diabetes. Eur. J. Endocrinol. 2006, 154, 899–906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grossmann, M. Testosterone and glucose metabolism in men: Current concepts and controversies. J. Endocrinol. 2013, 220, R37–R55. [Google Scholar] [CrossRef] [Green Version]
- Naharci, M.I.; Pinar, M.; Bolu, E.; Olgun, A. Effect of Testosterone on Insulin Sensitivity in Men with Idiopathic Hypogonadotropic Hypogonadism. Endocr. Pract. 2007, 13, 629–635. [Google Scholar] [CrossRef]
- Lenzi, A.; Lombardi, G.; Martino, E. Endocrinologia e Attività Motorie 160; Elsevier Srl.: Rome, Italy, 2008. [Google Scholar]
- Isidori, A.; Buvat, J.; Corona, G.; Goldstein, I.; Jannini, E.A.; Lenzi, A.; Porst, H.; Salonia, A.; Traish, A.M.; Maggi, M. A Critical Analysis of the Role of Testosterone in Erectile Function: From Pathophysiology to Treatment—A Systematic Review. Eur. Urol. 2014, 65, 99–112. [Google Scholar] [CrossRef]
- Fanelli, G.; Gevi, F.; Belardo, A.; Zolla, L. Metabolic patterns in insulin-sensitive male hypogonadism. Cell Death Dis. 2018, 9, 653. [Google Scholar] [CrossRef] [Green Version]
- Gevi, F.; Fanelli, G.; Zolla, L. Metabolic patterns in insulin-resistant male hypogonadism. Cell Death Dis. 2018, 9, 671. [Google Scholar] [CrossRef] [Green Version]
- Nicholson, J.K.; Wilson, I.D. Opinion: Understanding ‘global’ systems biology: Metabonomics and the continuum of metabolism. Nat. Rev. Drug. Discov. 2003, 2, 668–676. [Google Scholar] [CrossRef]
- Bogan, J.S.; Kandror, K.V. Biogenesis and regulation of insulin-responsive vesicles containing GLUT4. Curr. Opin. Cell Biol. 2010, 22, 506–512. [Google Scholar] [CrossRef] [Green Version]
- Kelly, D.M.; Akhtar, S.; Sellers, D.J.; Muraleedharan, V.; Channer, K.S.; Jones, T.H. Testosterone differentially regulates targets of lipid and glucose metabolism in liver, muscle and adipose tissues of the testicular feminised mouse. Endocrine 2016, 54, 504–515. [Google Scholar] [CrossRef] [PubMed]
- Ondrusova, M.; Spanikova, B.; Sevcikova, K.; Ondrus, D. Testosterone Deficiency and Bone Metabolism Damage in Testicular Cancer Survivors. Am. J. Men’s Health 2016, 12, 628–633. [Google Scholar] [CrossRef] [PubMed]
- Muthusamy, T.; Murugesan, P.; Balasubramanian, K. Sex steroids deficiency impairs glucose transporter 4 expression and its translocation through defective Akt phosphorylation in target tissues of adult male rat. Metabolism 2009, 58, 1581–1592. [Google Scholar] [CrossRef]
- Olorunshola, K.V.; Aliyu, O.F.; Achie, L.N. Testosterone and orchidectomy modulates intestinal fluid and glucose transport in albino wistar rat. Eur. J. Sci. Res. 2012, 76, 281–287. [Google Scholar]
- Sears, D.D.; Hsiao, G.; Hsiao, A.; Yu, J.G.; Courtney, C.H.; Ofrecio, J.M.; Chapman, J.; Subramaniam, S. Mechanisms of human insulin resistance and thiazolidinedione-mediated insulin sensitisation. Proc. Natl. Acad. Sci. USA 2009, 106, 18745–18750. [Google Scholar] [CrossRef] [Green Version]
- Martin, P.L.; Yin, J.-J.; Seng, V.; Casey, O.; Corey, E.; Morrissey, C.; Simpson, R.M.; Kelly, K. Androgen deprivation leads to increased carbohydrate metabolism and hexokinase 2-mediated survival in Pten/Tp53-deficient prostate cancer. Oncogene 2016, 36, 525–533. [Google Scholar] [CrossRef]
- Haymana, C.; Aydoğdu, A.; Soykut, B.; Erdem, O.; Ibrahimov, T.; Dinc, M.; Meric, C.; Basaran, Y.; Sonmez, A.; Azal, O. Oxidative stress status in congenital hypogonadism: An appraisal. Toxicol. Mech. Methods 2017, 27, 451–457. [Google Scholar] [CrossRef]
- Chen, C.-C.; Chen, C.-W.; Lin, P.-H.; Chou, J.-C.; Weng, T.-C.; Jian, C.-Y.; Hu, S.; Lai, W.-H.; Lieu, F.-K.; Wang, S.-W.; et al. Interactive effect of corticosterone and lactate on regulation of testosterone production in rat Leydig cells. J. Cell. Physiol. 2017, 232, 2135–2144. [Google Scholar] [CrossRef]
- Newgard, C.B.; An, J.; Bain, J.R.; Muehlbauer, M.J.; Stevens, R.D.; Lien, L.F.; Haqq, A.M.; Shah, S.H.; Arlotto, M.; Slentz, C.A.; et al. A Branched-Chain Amino Acid-Related Metabolic Signature that Differentiates Obese and Lean Humans and Contributes to Insulin Resistance. Cell Metab. 2009, 9, 311–326. [Google Scholar] [CrossRef] [Green Version]
- D’Alessandro, A.; Zolla, L. Metabolomics and cancer drug discovery: Let the cells do the talking. Drug Discov. Today 2012, 17, 3–9. [Google Scholar] [CrossRef]
- Zitzmann, M. Testosterone deficiency, insulin resistance and the metabolic syndrome. Nat. Rev. Endocrinol. 2009, 5, 673–681. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.K.; Fillmore, J.J.; Chen, Y.; Yu, C.; Moore, I.K.; Pypaert, M.; Lutz, E.P.; Kako, Y.; Velez-Carrasco, W.; Goldberg, I.J.; et al. Tissue-specific overexpression of lipoprotein lipase causes tissue-specific insulin resistance. Proc. Natl. Acad. Sci. USA 2001, 98, 7522–7527. [Google Scholar] [CrossRef] [PubMed]
- Delarue, J.; Magnan, C. Free fatty acids and insulin resistance. Curr. Opin. Clin. Nutr. Metab. Care 2007, 10, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Fanelli, G.; Belardo, A.; Savino, R.; Rinalducci, S.; Zolla, L. Testosterone replacement therapy in insulin-sensitive hypogonadal men restores phosphatidylcholine levels by regulation of arachidonic acid metabolism. J. Cell. Mol. Med. 2020, 24, 8266–8269. [Google Scholar] [CrossRef]
- Shah, S.H.; Crosslin, D.R.; Haynes, C.S.; Nelson, S.; Turer, C.B.; Stevens, R.D.; Muehlbauer, M.J.; Wenner, B.R.; Bain, J.R.; Laferrère, B.; et al. Branched-chain amino acid levels are associated with improvement in insulin resistance with weight loss. Diabetologia 2012, 55, 321–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laitinen, E.-M.; Hero, M.; Vaaralahti, K.; Tommiska, J.; Raivio, T. Bone mineral density, body composition and bone turnover in patients with congenital hypogonadotropic hypogonadism. Int. J. Androl. 2012, 35, 534–540. [Google Scholar] [CrossRef]
- Shigehara, K.; Izumi, K.; Kadono, Y.; Mizohami, A. Testosterone and bone health in men: A narrative review. J. Clin. Med. 2021, 10, 530. [Google Scholar] [CrossRef]
- Kok-Yong, C.; Ima-Nirwana, S.; Isa Naina, M.; Wan, Z.; Wan, N. Serum testosterone, sex hormone-binding globulin and total calcium levels predict the calcaneal speed of sound in men. Clinics 2012, 67, 911–916. [Google Scholar]
- Kok-Yong, C.; Soelaiman, I.M. Effects of annatto-derived tocotrienol supplementation on osteoporosis induced by testosterone deficiency in rats. Clin. Interv. Aging 2014, 9, 1247–1259. [Google Scholar]
- Peñafiel, R.; Ruzafa, C.; Monserrat, F.; Cremades, A. Gender-related differences in carnosine, anserine and lysine content of murine skeletal muscle. Amino Acids 2004, 26, 53–58. [Google Scholar] [CrossRef]
- Varanoske, A.N.; Hoffman, J.R.; Church, D.D.; Wang, R.; Baker, K.M.; Dodd, S.J.; Coker, N.A.; Oliveira, L.P.; Dawson, V.L.; Fukuda, D.H.; et al. Influence of Skeletal Muscle Carnosine Content on Fatigue during Repeated Resistance Exercise in Recreationally Active Women. Nutrients 2017, 9, 988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zolla, L.; Ceci, M. Plasma Metabolomics Profile of “Insulin Sensitive” Male Hypogonadism after Testosterone Replacement Therapy. Int. J. Mol. Sci. 2022, 23, 1916. [Google Scholar] [CrossRef] [PubMed]
- Zolla, L.; Grande, G.; Milardi, D. Plasma Metabonomics in Insulin-Resistant Hypogonadic Patients Induced by Testosterone Treatment. Int. J. Mol. Sci. 2022, 23, 7754. [Google Scholar] [CrossRef]
- Salloum, B.A.; Herkimer, C.; Lee, J.S.; Veiga-Lopez, A.; Padmanabhan, V. Developmental Programming: Prenatal and Postnatal Contribution of Androgens and Insulin in the Reprogramming of Estradiol Positive Feedback Disruptions in Prenatal Testosterone-Treated Sheep. Endocrinology 2012, 153, 2813–2822. [Google Scholar] [CrossRef]
- Munzer, T.; Mitchell, H.; Sorkin, J.D.; Blackman, M.R. Growth hormone and sex steroid effects on serum glucose, insulin, and lipid concentrations in healthy older women and man. J. Clin. Endocrinol. Metab. 2009, 94, 3833–3841. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Iemitsu, M.; Aizawa, K.; Ajisaka, R. Testosterone and DHEA activate the glucose metabolism-related signaling pathway in skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2008, 294, E961–E968. [Google Scholar] [CrossRef] [Green Version]
- Enoki, T.; Yoshida, Y.; Lally, J.; Hatta, H.; Bonen, A. Testosterone increases lactate transport, monocarboxylate transporter (MCT) 1 and MCT4 in rat skeletal muscle. J. Physiol. 2006, 577, 433–443. [Google Scholar] [CrossRef]
- Burns, J.S.; Manda, G. Metabolic Pathways of the Warburg Effect in Health and Disease: Perspectives of Choice, Chain or Chance. Int. J. Mol. Sci. 2017, 18, 2755. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.; Wang, S.-W.; Wang, R.-Y.; Wang, P.S. Stimulatory effect of lactate on testosterone production by rat Leydig cells. J. Cell. Biochem. 2001, 83, 147–154. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, J.; Wang, H. Correlation of blood glucose, serum chemerin and insulin resistance with NAFLD in patients with type 2 diabetes mellitus. Exp. Ther. Med. 2018, 15, 2936–2940. [Google Scholar] [CrossRef] [Green Version]
- Rato, L.; Alves, M.G.; Duarte, A.I.; Santos, M.S.; Moreira, P.I.; Cavaco, J.E.; Oliveira, P.F. Testosterone deficiency induced by progressive stages of diabetes mellitus impairs glucose metabolism and favours glycogenesis in mature rat Sertoli cells. Int. J. Biochem. Cell Biol. 2015, 66, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Go, Y.; Jeong, J.Y.; Jeoung, N.H.; Jeon, J.-H.; Park, B.-Y.; Kang, H.-J.; Ha, C.-M.; Choi, Y.-K.; Lee, S.J.; Ham, H.J.; et al. Inhibition of Pyruvate Dehydrogenase Kinase 2 Protects against Hepatic Steatosis through Modulation of Tricarboxylic Acid Cycle Anaplerosis and Ketogenesis. Diabetes 2016, 65, 2876–2887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laffel, L. Ketone Bodies: A Review of Physiology, Pathophysiology and Application of Monitoring to Diabetes. Diabetes Metab. Res. Rev. 1999, 15, 412–426. [Google Scholar] [CrossRef]
- Herzig, S.; Raemy, E.; Montessuit, S.; Veuthey, J.-L.; Zamboni, N.; Westermann, B.; Kunji, E.R.S.; Martinou, J.-C. Identification and Functional Expression of the Mitochondrial Pyruvate Carrier. Science 2012, 337, 93–96. [Google Scholar] [CrossRef] [PubMed]
- Fukami, K.; Yamagishi, S.-I.; Sakai, K.; Nasu, M.; Okuda, S. Effects of switching from oral administration to intravenous injection of L-carnitine on lipid metabolism in hemodialysis patients. Clin. Kidney J. 2014, 7, 470–474. [Google Scholar] [CrossRef] [Green Version]
- D’Antona, G.; Ragni, M.; Cardile, A.; Tedesco, L.; Dossena, M.; Bruttini, F.; Caliaro, F.; Corsetti, G.; Bottinelli, R.; Carruba, M.O.; et al. Branched-Chain Amino Acid Supplementation Promotes Survival and Supports Cardiac and Skeletal Muscle Mitochondrial Biogenesis in Middle-Aged Mice. Cell Metab. 2010, 12, 362–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheffield-Moore, M. Androgens and the control of skeletal muscle protein synthesis. Ann. Med. 2000, 32, 181–186. [Google Scholar] [CrossRef]
- Tenover, J.S. Effects of testosterone supplementation in the aging male. J. Clin. Endocrinol. Metab. 1992, 75, 1092–1098. [Google Scholar]
- Wang, C.; Cunningham, G.; Dobs, A.; Iranmanesh, A.; Matsumoto, A.M.; Snyder, P.J.; Weber, T.; Berman, N.; Hull, L.; Swerdloff, R.S. Long-Term Testosterone Gel (AndroGel) Treatment Maintains Beneficial Effects on Sexual Function and Mood, Lean and Fat Mass, and Bone Mineral Density in Hypogonadal Men. J. Clin. Endocrinol. Metab. 2004, 89, 2085–2098. [Google Scholar] [CrossRef] [Green Version]
- Thumelin, S.; Kohl, C.; Girard, J.; Pégorier, J.-P. Atypical expression of mitochondrial 3-hydroxy-3-methylglutaryl-CoA synthase in subcutaneous adipose tissue of male rats. J. Lipid Res. 1999, 40, 1071–1077. [Google Scholar] [CrossRef]
- Widenius, T.; Orava, M.; Vihko, R.; Ylikahri, R.; Eriksson, C. Inhibition of testosterone biosynthesis by ethanol: Multiple sites and mechanisms in dispersed leydig cells. J. Steroid Biochem. 1987, 28, 185–188. [Google Scholar] [CrossRef]
- Veech, R.L. The therapeutic implications of ketone bodies: The effects of ketone bodies in pathological conditions: Ketosis, ketogenic diet, redox states, insulin resistance, and mitochondrial metabolism. Prostaglandins Leukot. Essent. Fatty Acids 2004, 70, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, Y.; Harada, E.; Nakagawa, H.; Morikawa, Y.; Shono, M.; Kugimiya, F.; Yoshimura, M.; Yasue, H. The diabetic heart utilizes ketone bodies as an energy source. Metab. Clin. Exp. 2017, 77, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Bostock, E.C.S.; Kirkby, K.C.; Taylor, B.V.; Hawrelak, J.A. Consumer Reports of “Keto Flu” Associated with the Ketogenic Diet. Front. Nutr. 2020, 7, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bostock, E.C.S.; Kirkby, K.C.; Taylor, B.V.M. The Current Status of the Ketogenic Diet in Psychiatry. Front. Psychiatry 2017, 8, 43. [Google Scholar] [CrossRef]
- Van der Vaart, A.; Knol, M.G.E.; De Bost, M.H.; Bakker, S.J.L.; Connelly, M.A.; Garcia, E.; Bilo, H.J.G.; van Dijk, P.R.; Dullant, R.P.F. The paradoxical role of circulating ketone bodies in glycemic controlof individuals with type 2 diabetes: High risk, high reward? Biomolecules 2022, 12, 1318. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zolla, L. On the Need to Distinguish between Insulin-Normal and Insulin-Resistant Patients in Testosterone Therapy. Int. J. Mol. Sci. 2022, 23, 12730. https://doi.org/10.3390/ijms232112730
Zolla L. On the Need to Distinguish between Insulin-Normal and Insulin-Resistant Patients in Testosterone Therapy. International Journal of Molecular Sciences. 2022; 23(21):12730. https://doi.org/10.3390/ijms232112730
Chicago/Turabian StyleZolla, Lello. 2022. "On the Need to Distinguish between Insulin-Normal and Insulin-Resistant Patients in Testosterone Therapy" International Journal of Molecular Sciences 23, no. 21: 12730. https://doi.org/10.3390/ijms232112730
APA StyleZolla, L. (2022). On the Need to Distinguish between Insulin-Normal and Insulin-Resistant Patients in Testosterone Therapy. International Journal of Molecular Sciences, 23(21), 12730. https://doi.org/10.3390/ijms232112730