

Characteristic of the Ascorbate Oxidase Gene Family in Beta vulgaris and Analysis of the Role of AAO in Response to Salinity and Drought in Beet

 , ,
, ,  ,
,  and
and
Abstract
:1. Introduction
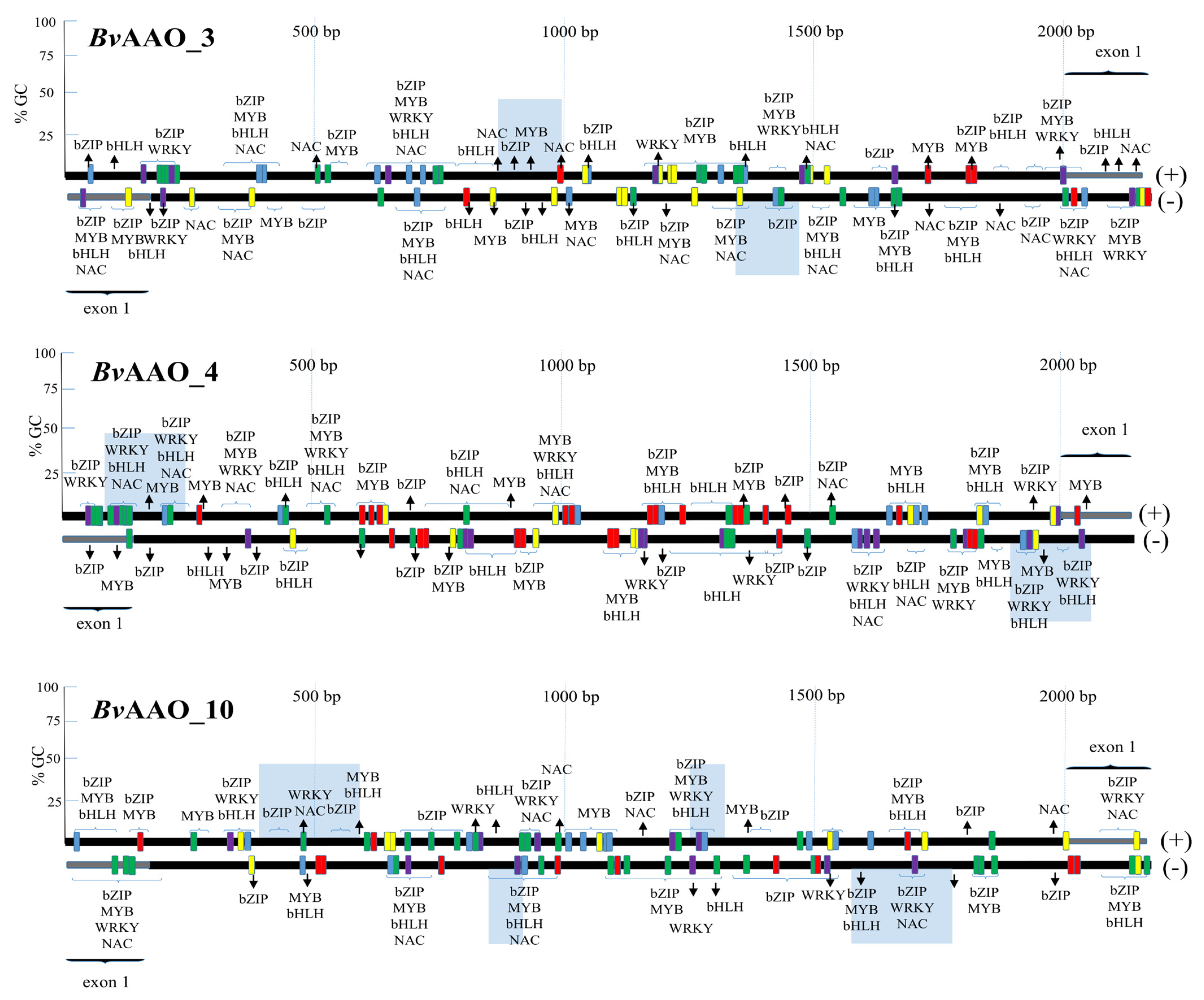
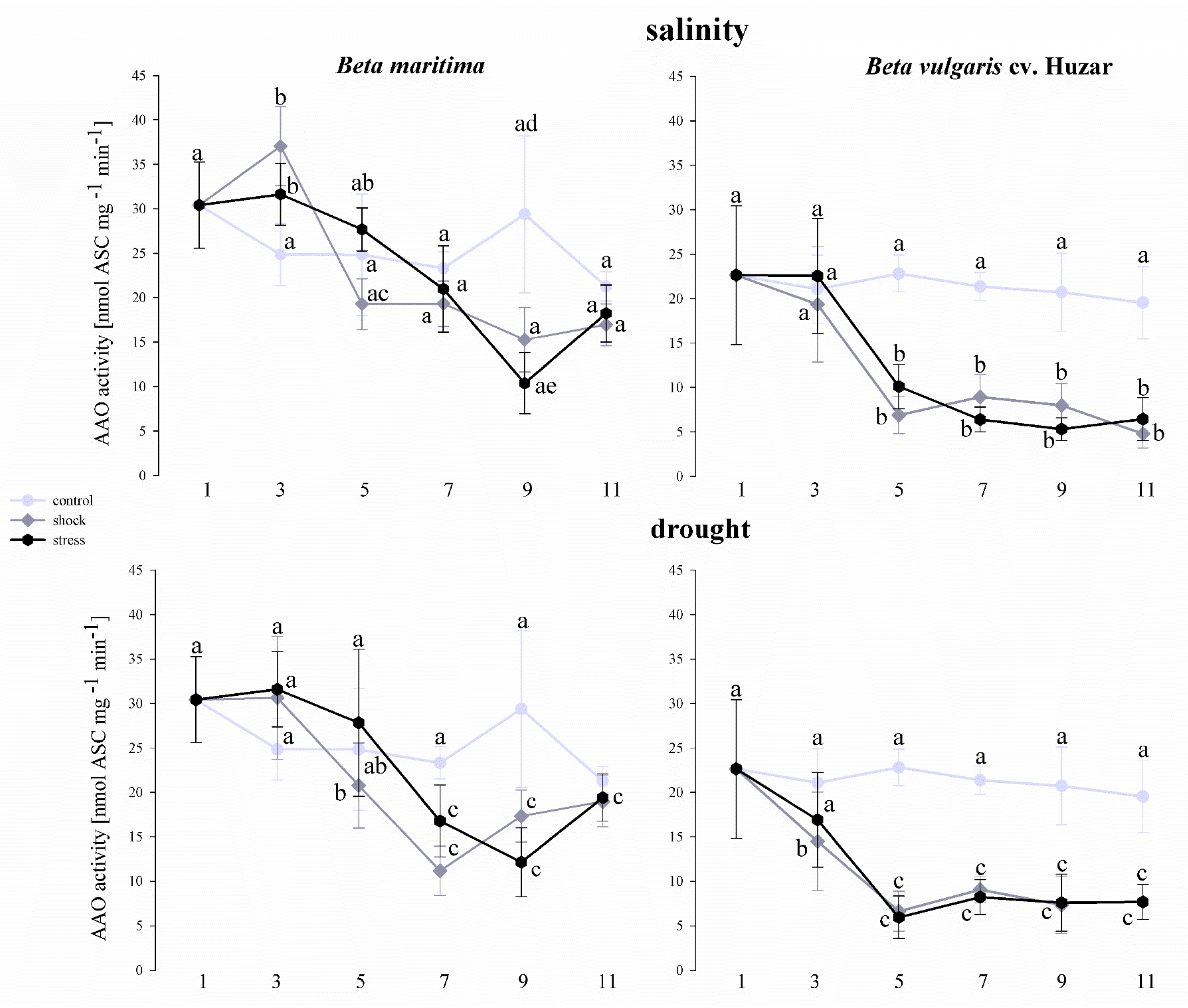
2. Results
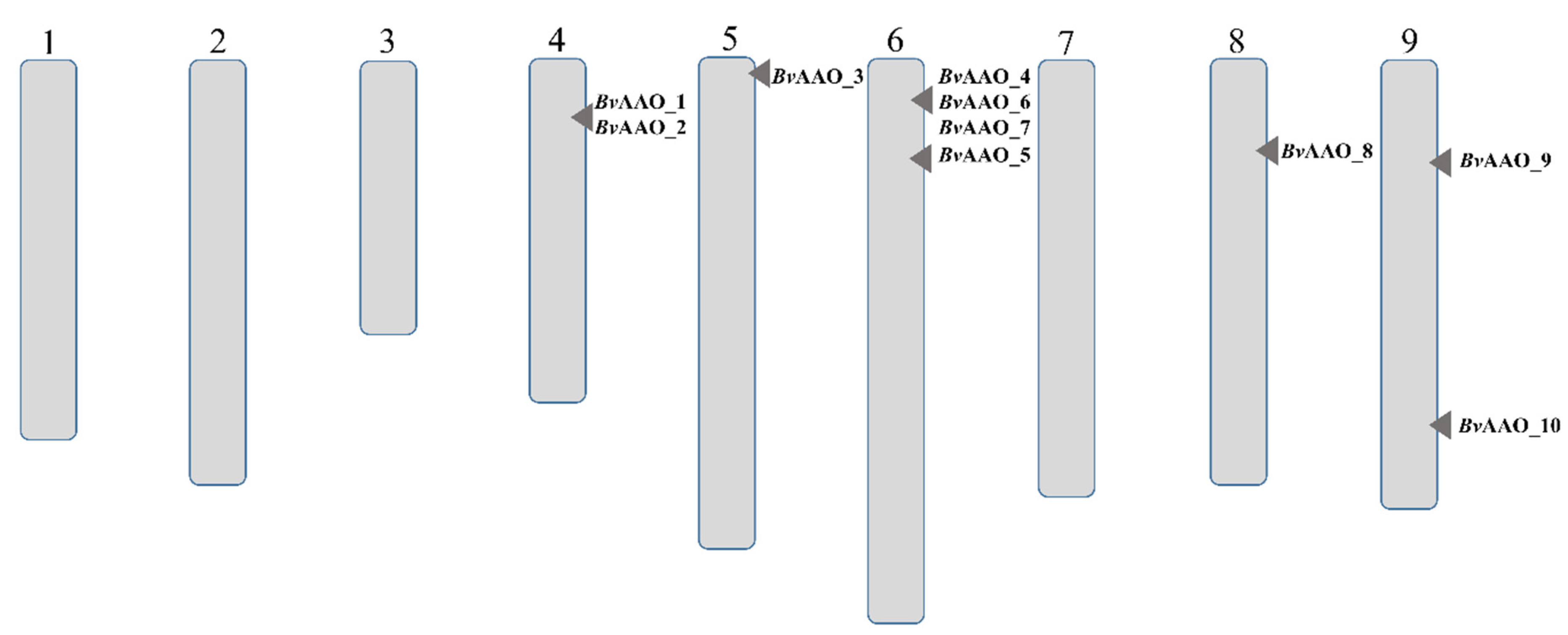
2.1. Characteristics of Genes and Proteins of Beta vulgaris Ascorbate Oxidases
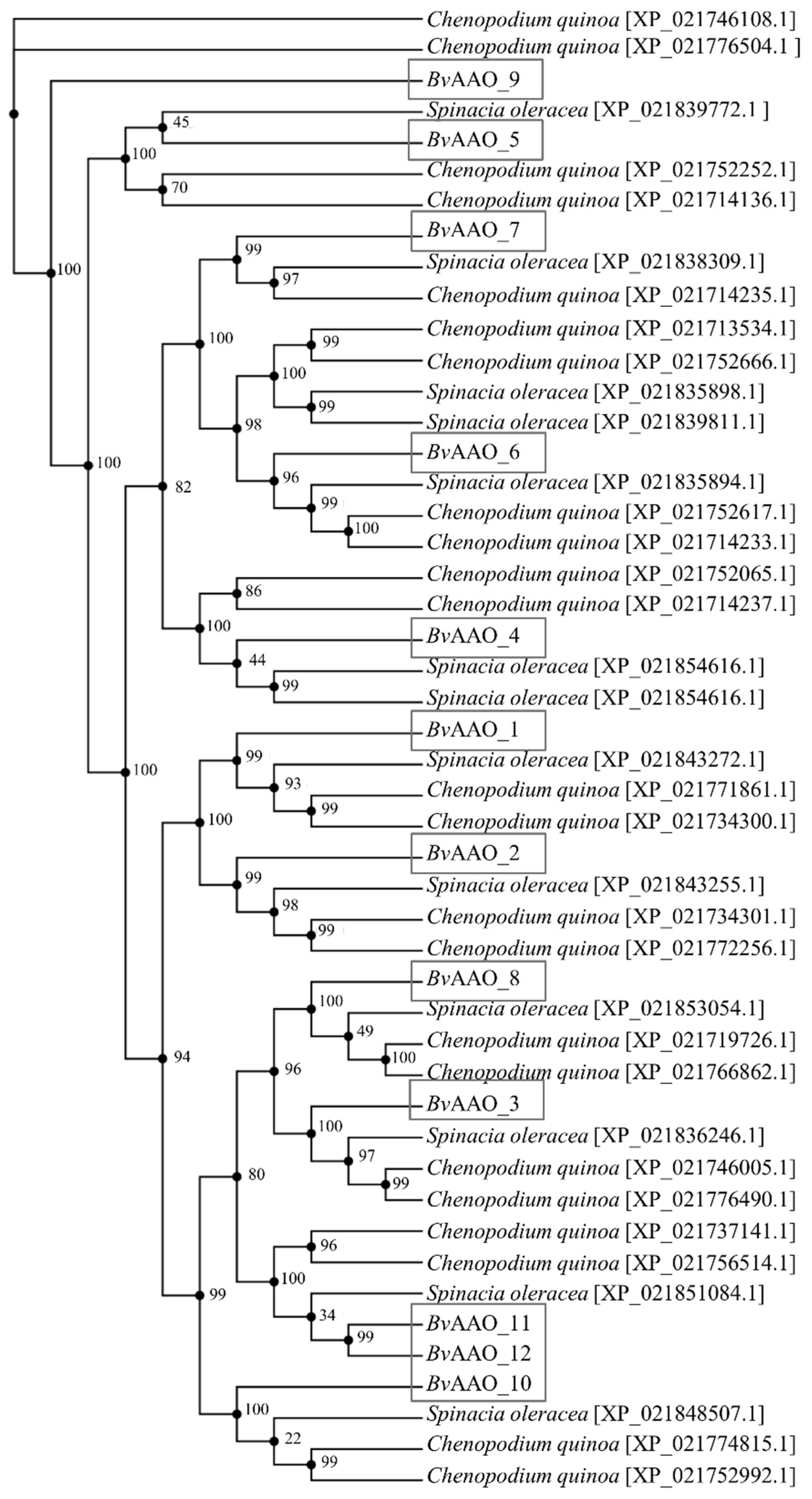
2.2. Phylogenetic and Evolutionary Analyses of AAO Gene Family
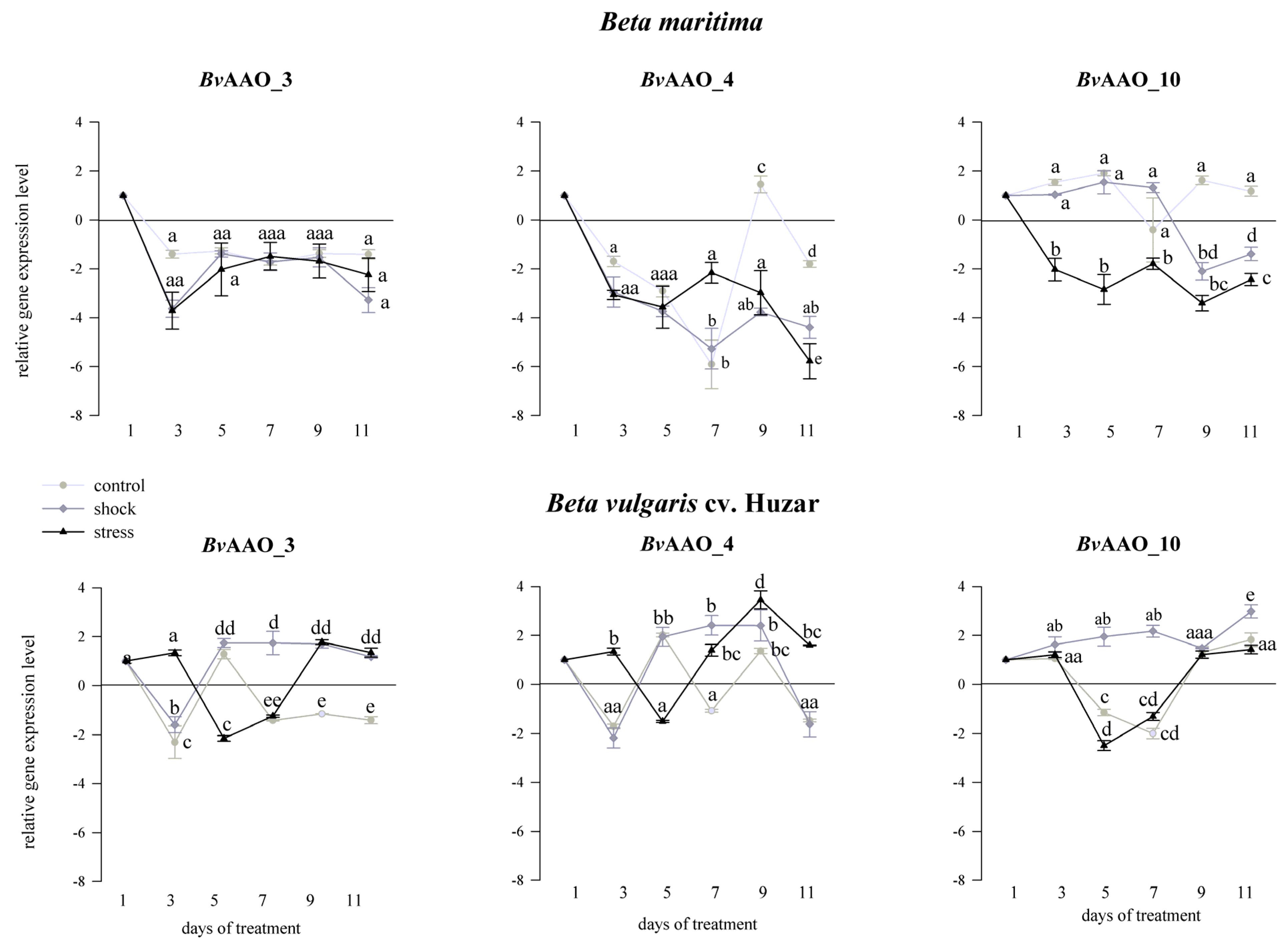
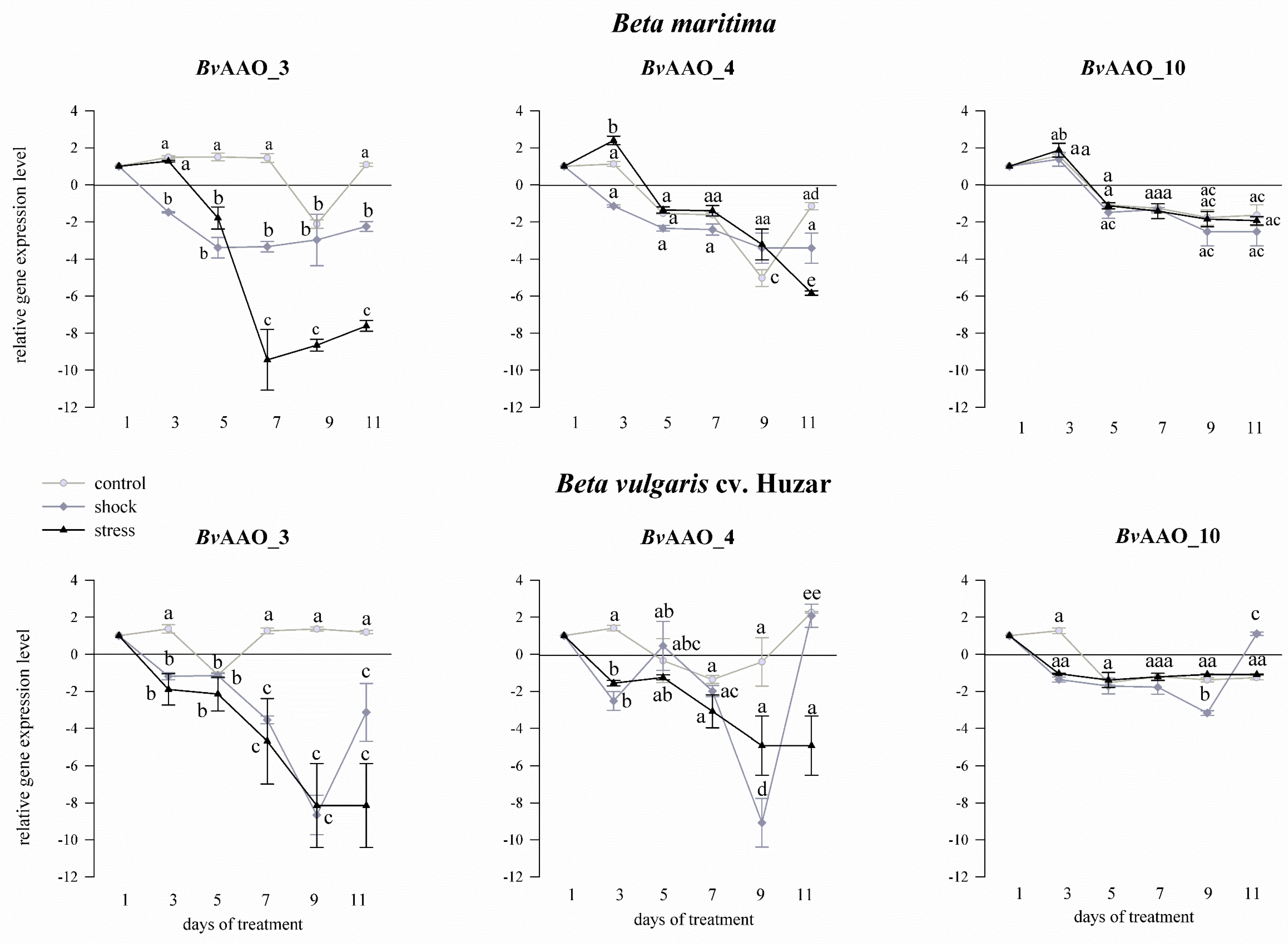
2.3. Analysis of the AAO Beta vulgaris Gene Expression Profile under Drought and Salinity Conditions
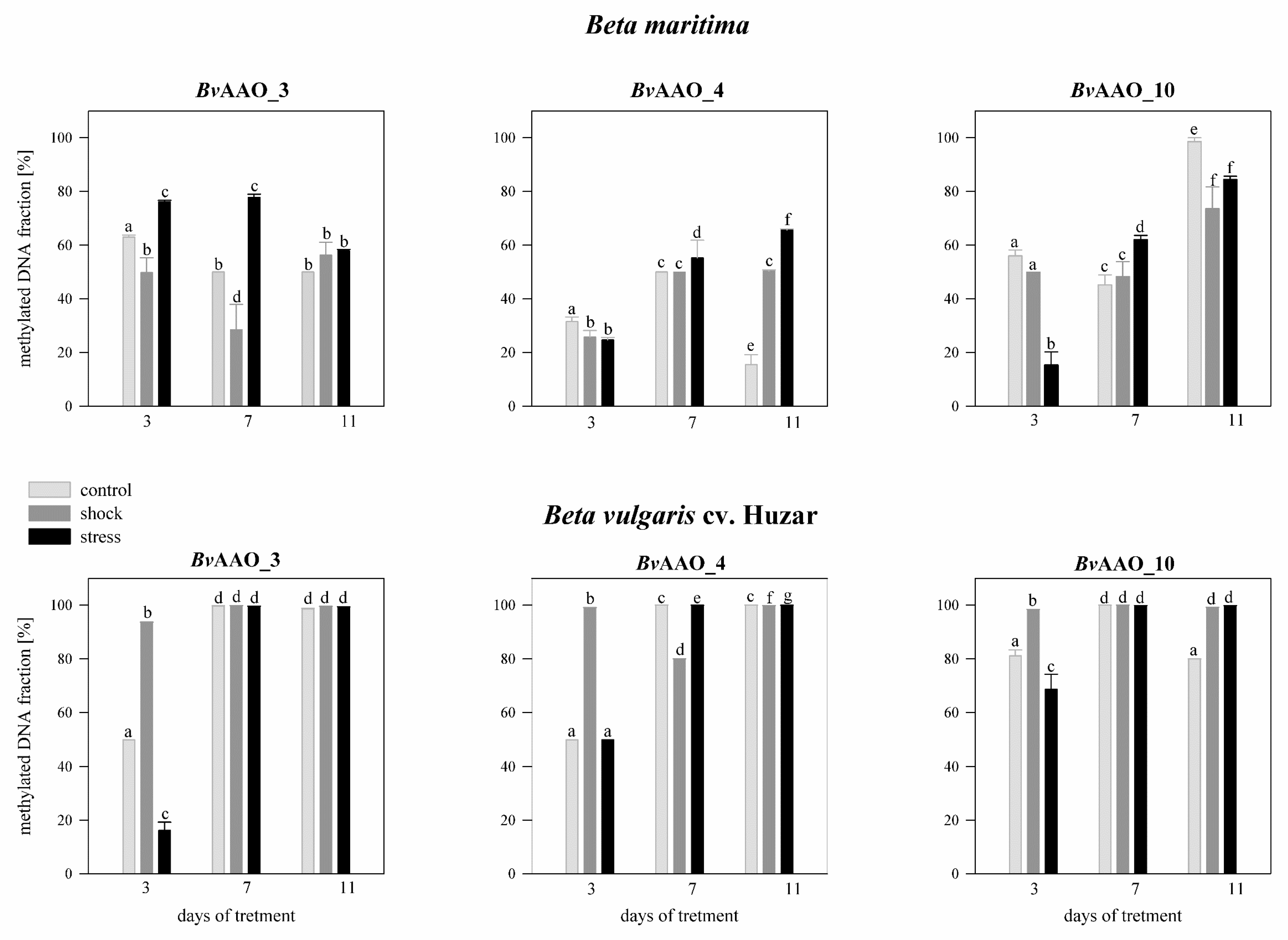
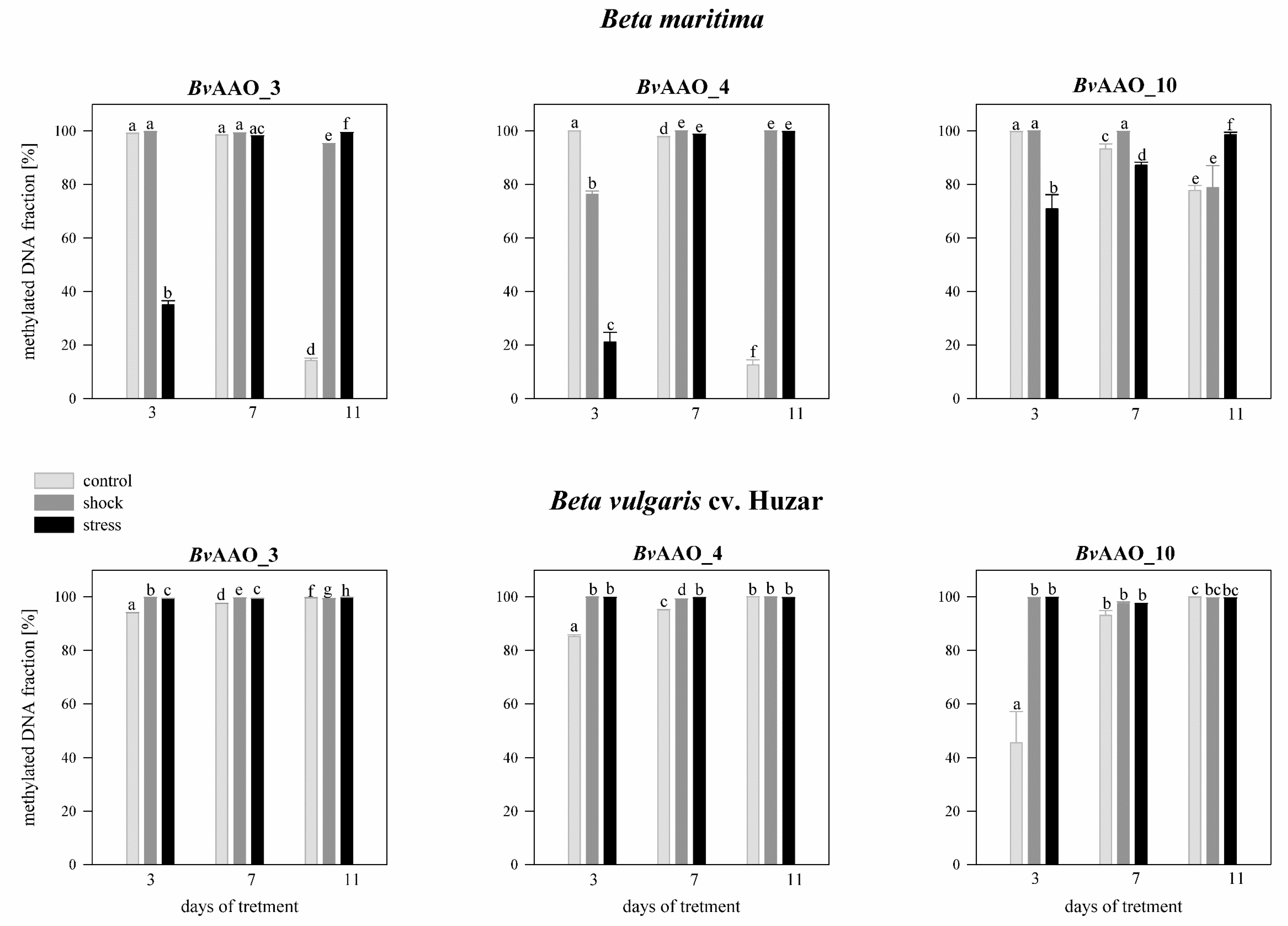
2.4. Analysis of the Methylation Level of the CpG Islands of the Promoter Regions of the AAO Beta vulgaris Genes under Drought and Salinity Conditions
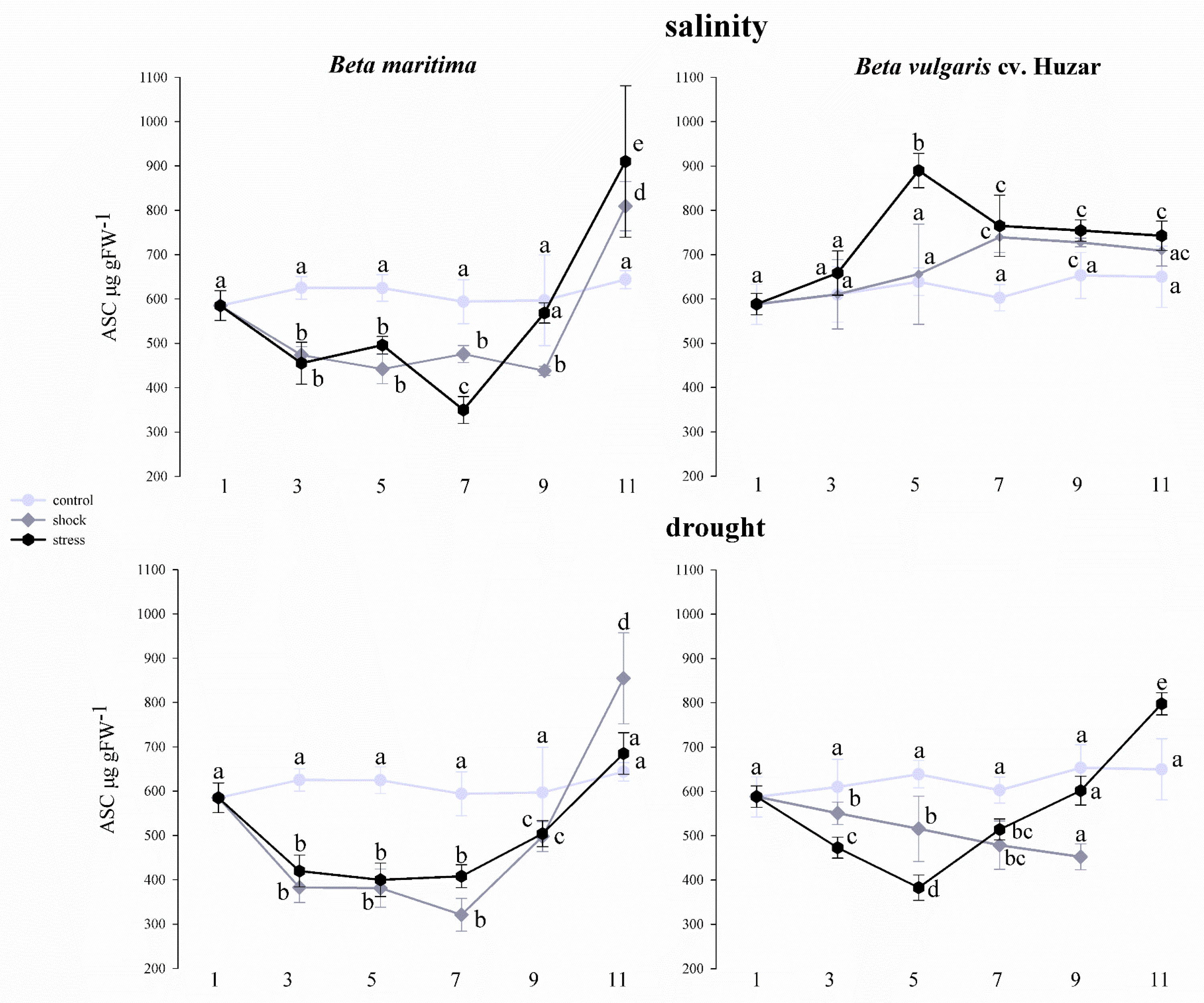
2.5. Analysis of Total Ascorbate Concentration in Beta vulgaris Leaves under Drought and Salinity Conditions
2.6. Analysis of Ascorbate Oxidase Activity in Beta vulgaris Leaves in Drought and Saline Conditions
3. Discussion
4. Materials and Methods
4.1. Identification of Ascorbate Oxidase Sequences in Beta vulgaris
4.2. Bioinformatic Analysis
4.3. Sequence Alignment and Phylogenetic Analysis
4.4. Plant Material
4.5. Exposing Plants to Salinity
4.6. Exposing Plants to Drought
4.7. RT qPCR Analysis
4.8. Analysis of the CpG Island Methylation Level of Selected Beet AAO Genes
4.9. Determination of the Concentration of Total Ascorbate
4.10. Measurement of the Enzymatic Activity of Ascorbate Oxidase
4.11. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Foyer, C.H.; Kyndt, T.; Hancock, R.D. Vitamin C in Plants: Novel Concepts, New Perspectives, and Outstanding Issues. Antioxid. Redox Signal. 2020, 32, 463–485. [Google Scholar] [CrossRef] [PubMed]
- Pignocchi, C.; Foyer, C.H. Apoplastic ascorbate metabolism and its role in the regulation of cell signalling. Curr. Opin. Plant Biol. 2003, 6, 379–389. [Google Scholar] [CrossRef]
- Pignocchi, C.; Kiddle, G.; Hernandez, I.; Foster, S.J.; Asensi, A.; Taybi, T.; Barnes, J.; Foyer, C.H. Ascorbate oxidase-dependent changes in the redox state of the apoplast modulate gene transcript accumulation leading to modified hormone signaling and orchestration of defense processes in tobacco. Plant Physiol. 2006, 141, 423–435. [Google Scholar] [CrossRef] [Green Version]
- Fotopoulos, V.; De Tullio, M.C.; Barnes, J.; Kanellis, A.K. Altered stomatal dynamics in ascorbate oxidase over-expressing tobacco plants suggest a role for dehydroascorbate signalling. J. Exp. Bot. 2008, 59, 729–737. [Google Scholar] [CrossRef]
- De Tullio, M.C.; Guether, M.; Balestrini, R. Ascorbate oxidase is the potential conductor of a symphony of signaling pathways. Plant Signal. Behav. 2013, 8, e23213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smirnoff, N. The function and metabolism of ascorbic acid in plants. Ann. Bot. 1996, 78, 661–669. [Google Scholar] [CrossRef] [Green Version]
- Garchery, C.; Gest, N.; Do, P.T.; Alhagdow, M.; Baldet, P.; Menard, G.; Rothan, C.; Massot, C.; Gautier, H.; Aarrouf, J.; et al. A diminution in ascorbate oxidase activity affects carbon allocation and improves yield in tomato under water deficit. Plant Cell Environ. 2013, 36, 159–175. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.R.; Nobleza, N.; Demeestere, K.; Kyndt, T. Ascorbate Oxidase Induces Systemic Resistance in Sugar Beet Against Cyst Nematode Heterodera schachtii. Front. Plant Sci. 2020, 11, 591715. [Google Scholar] [CrossRef] [PubMed]
- Sanmartin, M.; Pateraki, I.; Chatzopoulou, F.; Kanellis, A.K. Differential expression of the ascorbate oxidase multigene family during fruit development and in response to stress. Planta 2007, 225, 873–885. [Google Scholar] [CrossRef]
- Xiao, M.; Li, Z.; Zhu, L.; Wang, J.; Zhang, B.; Zheng, F.; Zhao, B.; Zhang, H.; Wang, Y.; Zhang, Z. The Multiple Roles of Ascorbate in the Abiotic Stress Response of Plants: Antioxidant, Cofactor, and Regulator. Front. Plant Sci. 2021, 12, 598173. [Google Scholar] [CrossRef]
- Pan, Z.; Chen, L.; Wang, F.; Song, W.; Cao, A.; Xie, S.; Chen, X.; Jin, X.; Li, H. Genome-Wide Identification and Expression Analysis of the Ascorbate Oxidase Gene Family in Gossypium hirsutum Reveals the Critical Role of GhAO1A in Delaying Dark-Induced Leaf Senescence. Int. J. Mol. Sci. 2019, 20, 6167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghaffari, H.; Tadayon, M.R.; Nadeem, M.; Cheema, M.; Razmjoo, J. Proline mediated changes in antioxidant enzymatic activities and physiology of sugar beet under drought stress. Acta Physiol. Plant. 2019, 41, 23. [Google Scholar] [CrossRef]
- Liu, H.; Wang, Q.; Yu, M.; Zhang, Y.; Wu, Y.; Zhang, H. Transgenic salt-tolerant sugar beet (Beta vulgaris L.) constitutively expressing an Arabidopsis thaliana vacuolar Na/H antiporter gene, AtNHX3, accumulates more soluble sugar but less salt in storage roots. Plant Cell Environ. 2008, 31, 1325–1334. [Google Scholar] [CrossRef] [PubMed]
- Maung, T.A.; Gustafson, C.R. The economic feasibility of sugar beet biofuel production in central North Dakota. Biomass and Bioenergy 2011, 35, 3737–3747. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Nan, J.; Yu, B. OMICS Technologies and Applications in Sugar Beet. Front. Plant Sci. 2016, 7, 900. [Google Scholar] [CrossRef] [Green Version]
- Pinheiro, C.; Ribeiro, I.C.; Reisinger, V.; Planchon, S.; Veloso, M.M.; Renaut, J.; Eichacker, L.; Ricardo, C.P. Salinity effect on germination, seedling growth and cotyledon membrane complexes of a Portuguese salt marsh wild beet ecotype. Theor. Exp. Plant Physiol. 2018, 30, 113–127. [Google Scholar] [CrossRef]
- Skorupa, M.; Golebiewski, M.; Kurnik, K.; Niedojadlo, J.; Kesy, J.; Klamkowski, K.; Wojcik, K.; Treder, W.; Tretyn, A.; Tyburski, J. Salt stress vs. salt shock-the case of sugar beet and its halophytic ancestor. BMC Plant Biol. 2019, 19, 57. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, I.C.; Pinheiro, C.; Ribeiro, C.M.; Veloso, M.M.; Simoes-Costa, M.C.; Evaristo, I.; Paulo, O.S.; Ricardo, C.P. Genetic Diversity and Physiological Performance of Portuguese Wild Beet (Beta vulgaris spp. maritima) from Three Contrasting Habitats. Front. Plant Sci. 2016, 7, 1293. [Google Scholar] [CrossRef] [Green Version]
- Kraft, T.; Fridlund, B.; Hjerdin, A.; Sall, T.; Tuvesson, S.; Hallden, C. Estimating genetic variation in sugar beets and wild beets using pools of individuals. Genome 1997, 40, 527–533. [Google Scholar] [CrossRef]
- Taghizadegan, M.; Toorchi, M.; Moghadam Vahed, M.; Khayamim, S. Evaluation of sugar beet breeding populations based morpho-physiological characters under salinity stress. Pak. J. Bot. 2019, 51, 11–17. [Google Scholar] [CrossRef]
- Daoud, S.; Harrouni, C.; Huchzermeyer, B.; Koyro, H.W. Comparison of salinity tolerance of two related subspecies of Beta vulgaris: The sea beet (Beta vulgaris ssp. maritima) and the sugar beet (Beta vulgaris ssp. vulgaris). In Biosaline Agriculture and High Salinity Tolerance; Abdelly, C., Öztürk, M., Ashraf, M., Grignon, C., Eds.; Birkhäuser Basel: Basel, Switzerland, 2008. [Google Scholar] [CrossRef]
- Koyro, H.-W.; Daoud, S.; Harrouni, C.; Huchzermeyer, B. Strategies of a potential cash crop halophyte (Beta vulgaris ssp. maritima) to avoid salt injury. Trop. Ecol. 2006, 47, 191–200. [Google Scholar]
- Yolcu, S.; Alavilli, H.; Ganesh, P.; Panigrahy, M.; Song, K. Salt and Drought Stress Responses in Cultivated Beets (Beta vulgaris L.) and Wild Beet (Beta maritima L.). Plants 2021, 10, 1843. [Google Scholar] [CrossRef] [PubMed]
- Skorupa-Kłaput, M.; Szczepanek, J.; Kurnik, K.; Tretyn, A.; Tyburski, J. The expression patterns of plasma membrane aquaporins in leaves of sugar beet and its halophyte relative, Beta vulgaris ssp. maritima, in response to salt stress. Biologia 2015, 70, 467–477. [Google Scholar] [CrossRef]
- Dunajska-Ordak, K.; Skorupa-Klaput, M.; Kurnik, K.; Tretyn, A.; Tyburski, J. Cloning and Expression Analysis of a Gene Encoding for Ascorbate Peroxidase and Responsive to Salt Stress in Beet (Beta vulgaris). Plant Mol. Biol. Report 2014, 32, 162–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bor, M.; Özdemir, F.; Türkan, I. The effect of salt stress on lipid peroxidation and antioxidants in leaves of sugar beet Beta vulgaris L. and wild beet Beta maritima L. Plant Sci. 2003, 164, 77–84. [Google Scholar] [CrossRef]
- Wisniewska, A.; Andryka-Dudek, P.; Czerwinski, M.; Choluj, D. Fodder beet is a reservoir of drought tolerance alleles for sugar beet breeding. Plant Physiol. Biochem. 2019, 145, 120–131. [Google Scholar] [CrossRef]
- Van Geyt, J.P.C.; Lange, W.; Oleo, M.; De Bock, T.S.M. Natural variation within the genus Beta and its possible use for breeding sugar beet: A review. Euphytica 1990, 49, 57–76. [Google Scholar] [CrossRef]
- Yu, B.; Chen, M.; Grin, I.; Ma, C. Mechanisms of Sugar Beet Response to Biotic and Abiotic Stresses. In Mechanisms of Genome Protection and Repair; Zharkov, D.O., Ed.; Springer International Publishing: Cham, Switzerland, 2020; pp. 167–194. [Google Scholar] [CrossRef]
- Rozema, J.; Cornelisse, D.; Zhang, Y.; Li, H.; Bruning, B.; Katschnig, D.; Broekman, R.; Ji, B.; van Bodegom, P. Comparing salt tolerance of beet cultivars and their halophytic ancestor: Consequences of domestication and breeding programmes. AoB Plants 2015, 7, plu083. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhan, Y.; Wu, C.; Gong, S.; Zhu, N.; Chen, S.; Li, H. Cloning of a cystatin gene from sugar beet M14 that can enhance plant salt tolerance. Plant Sci. 2012, 191, 93–99. [Google Scholar] [CrossRef]
- Chołuj, D.; Wisniewska, A.; Szafranski, K.M.; Cebula, J.; Gozdowski, D.; Podlaski, S. Assessment of the physiological responses to drought in different sugar beet genotypes in connection with their genetic distance. J. Plant Physiol. 2014, 171, 1221–1230. [Google Scholar] [CrossRef]
- Munns, R. Plant adaptations to salt and water stress: Differences and commonalities. Adv. Bot. Res. 2011, 57, 1–32. [Google Scholar] [CrossRef]
- Shavrukov, Y. Salt stress or salt shock: Which genes are we studying? J. Exp. Bot. 2013, 64, 119–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talamè, V.; Ozturk, N.Z.; Bohnert, H.J.; Tuberosa, R. Barley transcript profiles under dehydration shock and drought stress treatments: A comparative analysis. J. Exp. Bot. 2007, 58, 229–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wedeking, R.; Mahlein, A.K.; Steiner, U.; Oerke, E.C.; Goldbach, H.E.; Wimmer, M.A. Osmotic adjustment of young sugar beets (Beta vulgaris) under progressive drought stress and subsequent rewatering assessed by metabolite analysis and infrared thermography. Funct. Plant Biol. 2017, 44, 119–133. [Google Scholar] [CrossRef] [Green Version]
- Khayamim, S.; Afshari, R.T.; Sadeghian, S.Y.; Poustini, K.; Rouzbeh, F.; Abbasi, Z. Seed Germination, Plant Establishment, and Yield of Sugar Beet Genotypes under Salinity Stress. J. Agr. Sci. Technol. 2014, 16, 779–790. [Google Scholar]
- Lv, X.; Chen, S.; Wang, Y. Advances in Understanding the Physiological and Molecular Responses of Sugar Beet to Salt Stress. Front. Plant Sci. 2019, 10, 1431. [Google Scholar] [CrossRef] [Green Version]
- Romano, A.; Sorgonà, A.; Lupini, A.; Araniti, F.; Stevanato, P.; Cacco, G.; Abenavoli, M.R. Morpho-physiological responses of sugar beet (Beta vulgaris L.) genotypes to drought stress. Acta Physiol. Plant. 2013, 35, 853–865. [Google Scholar] [CrossRef]
- Szymanska, S.; Tyburski, J.; Piernik, A.; Sikora, M.; Mazur, J.; Katarzyna, H. Raising Beet Tolerance to Salinity through Bioaugmentation with Halotolerant Endophytes. Agronomy 2020, 10, 1571. [Google Scholar] [CrossRef]
- Kim, J.M.; To, T.K.; Nishioka, T.; Seki, M. Chromatin regulation functions in plant abiotic stress responses. Plant Cell Environ. 2010, 33, 604–611. [Google Scholar] [CrossRef]
- Skorupa, M.; Szczepanek, J.; Mazur, J.; Domagalski, K.; Tretyn, A.; Tyburski, J. Salt stress and salt shock differently affect DNA methylation in salt-responsive genes in sugar beet and its wild, halophytic ancestor. PLoS ONE 2021, 16, e0251675. [Google Scholar] [CrossRef]
- Mirouze, M.; Paszkowski, J. Epigenetic contribution to stress adaptation in plants. Curr. Opin. Plant Biol. 2011, 14, 267–274. [Google Scholar] [CrossRef]
- Colaneri, A.C.; Jones, A.M. Genome-Wide Quantitative Identification of DNA Differentially Methylated Sites in Arabidopsis Seedlings Growing at Different Water Potential. PLoS ONE 2013, 8, e59878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Y.; Ji, D.; Li, S.; Wang, P.; Li, Q.; Xiang, F. The Dynamic Changes of DNA Methylation and Histone Modifications of Salt Responsive Transcription Factor Genes in Soybean. PLoS ONE 2012, 7, e41274. [Google Scholar] [CrossRef] [PubMed]
- Paul, A.; Dasgupta, P.; Roy, D.; Chaudhuri, S. Comparative analysis of Histone modifications and DNA methylation at OsBZ8 locus under salinity stress in IR64 and Nonabokra rice varieties. Plant Mol Biol. 2017, 95, 63–88. [Google Scholar] [CrossRef] [PubMed]
- Choi, C.S.; Sano, H. Abiotic-stress induces demethylation and transcriptional activation of a gene encoding a glycerophosphodiesterase-like protein in tobacco plants. Mol. Genet. Genom. 2007, 277, 589–600. [Google Scholar] [CrossRef]
- Yolcu, S.; Ozdemir, F.; Güler, A.; Bor, M. Histone acetylation influences the transcriptional activation of POX in Beta vulgaris L. and Beta maritima L. under salt stress. Plant Physiol. Biochem. 2016, 100, 37–46. [Google Scholar] [CrossRef]
- Batth, R.; Singh, K.; Kumari, S.; Mustafiz, A. Transcript Profiling Reveals the Presence of Abiotic Stress and Developmental Stage Specific Ascorbate Oxidase Genes in Plants. Front. Plant Sci. 2017, 8, 198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelgawad, K.F.; El-Mogy, M.M.; Mohamed, M.I.A.; Garchery, C.; Stevens, R.G. Increasing ascorbic acid content and salinity tolerance of cherry tomato plants by suppressed expression of the ascorbate oxidase gene. Agronomy 2019, 9, 51. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, A.; Bhuiyan, M.N.; Waditee, R.; Tanaka, Y.; Esaka, M.; Oba, K.; Jagendorf, A.T.; Takabe, T. Suppressed expression of the apoplastic ascorbate oxidase gene increases salt tolerance in tobacco and Arabidopsis plants. J. Exp. Bot. 2005, 56, 1785–1796. [Google Scholar] [CrossRef] [Green Version]
- Fotopoulos, V.; Sanmartin, M.; Kanellis, A.K. Effect of ascorbate oxidase over-expression on ascorbate recycling gene expression in response to agents imposing oxidative stress. J. Exp. Bot. 2006, 57, 3933–3943. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Li, H.; Shu, W.; Zhang, C.; Zhang, W.; Ye, Z. Suppressed Expression of Ascorbate Oxidase Gene Promotes Ascorbic Acid Accumulation in Tomato Fruit. Plant Mol. Biol. Report. 2010, 29, 638–645. [Google Scholar] [CrossRef]
- Caputo, E.; Ceglie, V.; Lippolis, M.; La Rocca, N.; Tullio, M. Identification of a NaCl-induced ascorbate oxidase activity in Chaetomorpha linum suggests a novel mechanism of adaptation to increased salinity. Environ. Exp. Bot. 2010, 69, 63–67. [Google Scholar] [CrossRef]
- He, S.B.; Hu, A.L.; Zhuang, Q.Q.; Peng, H.P.; Deng, H.H.; Chen, W.; Hong, G.L. Ascorbate Oxidase Mimetic Activity of Copper(II) Oxide Nanoparticles. Chembiochem 2020, 21, 978–984. [Google Scholar] [CrossRef]
- Garg, R.; Narayana Chevala, V.; Shankar, R.; Jain, M. Divergent DNA methylation patterns associated with gene expression in rice cultivars with contrasting drought and salinity stress response. Sci. Rep. 2015, 5, 14922. [Google Scholar] [CrossRef] [Green Version]
- Yaish, M.W.; Al-Lawati, A.; Al-Harrasi, I.; Patankar, H.V. Genome-wide DNA Methylation analysis in response to salinity in the model plant caliph medic (Medicago truncatula). BMC Genomics 2018, 19, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoang, M.T.T.; Doan, M.T.A.; Nguyen, T.; Tra, D.-P.; Chu, T.N.; Dang, T.P.T.; Quach, P.N.D. Phenotypic Characterization of Arabidopsis Ascorbate and Glutathione Deficient Mutants under Abiotic Stresses. Agronomy 2021, 11, 764. [Google Scholar] [CrossRef]
- Farooq, A.; Bukhari, S.A.; Akram, N.A.; Ashraf, M.; Wijaya, L.; Alyemeni, M.N.; Ahmad, P. Exogenously Applied Ascorbic Acid-Mediated Changes in Osmoprotection and Oxidative Defense System Enhanced Water Stress Tolerance in Different Cultivars of Safflower (Carthamus tinctorious L.). Plants 2020, 9, 104. [Google Scholar] [CrossRef] [Green Version]
- Noman, A.; Ali, S.; Naheed, F.; Ali, Q.; Farid, M.; Rizwan, M.; Irshad, M.K. Foliar application of ascorbate enhances the physiological and biochemical attributes of maize (Zea mays L.) cultivars under drought stress. Arch. Agron. Soil Sci. 2015, 61, 1659–1672. [Google Scholar] [CrossRef]
- Chen, X.; Zhou, Y.; Cong, Y.; Zhu, P.; Xing, J.; Cui, J.; Xu, W.; Shi, Q.; Diao, M.; Liu, H.-y. Ascorbic Acid-Induced Photosynthetic Adaptability of Processing Tomatoes to Salt Stress Probed by Fast OJIP Fluorescence Rise. Front. Plant Sci. 2021, 12, 594400. [Google Scholar] [CrossRef] [PubMed]
- Airaki, M.; Leterrier, M.; Mateos, R.M.; Valderrama, R.; Chaki, M.; Barroso, J.B.; Del RÍO, L.A.; Palma, J.M.; Corpas, F.J. Metabolism of reactive oxygen species and reactive nitrogen species in pepper (Capsicum annuum L.) plants under low temperature stress. Plant Cell Environ. 2012, 35, 281–295. [Google Scholar] [CrossRef] [PubMed]
- Kanayama, Y.; Sato, K.; Ikeda, H.; Tamura, T.; Nishiyama, M.; Kanahama, K. Seasonal changes in abiotic stress tolerance and concentrations of tocopherol, sugar, and ascorbic acid in sea buckthorn leaves and stems. Sci. Hortic. 2013, 164, 232–237. [Google Scholar] [CrossRef]
- Venkatesh, J.; Park, S.W. Role of L-ascorbate in alleviating abiotic stresses in crop plants. Bot. Stud. 2014, 55, 38. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name in This Work | Name/ GeneID | Location | ORF | Exons | AA | MW (kDa) | pI | Signal P | Plant- mPLoc | Yloc | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | BvAAO_1 | LOC104890152 | Chromosome 4, NC_025815.2 (3428816–3432297) | 2253 | 4 | 557 | 62.94 | 8.98 | Yes, 24–25 (0.96) | Cell membrane | Peroxisome (62.51%) |
| 2 | BvAAO_2 | LOC104890153 | Chromosome 4, NC_025815.2 (3436425–3439102) | 2145 | 4 | 558 | 63.30 | 8.79 | Yes, 25–26 (0.94) | Cell membrane | Cytoplasm (86%) |
| 3 | BvAAO_3 | LOC104892300 | Chromosome 5, NC_025816.2 (3577518–3582274) | 2478 | 7 | 549 | 61.00 | 8.40 | Yes, 24–25 (0.93) | Cell membrane | Peroxisome (64.50%) |
| 4 | BvAAO_4 | LOC104895593 | Chromosome 6, NC_025817.2 (8750906–8755896) | ||||||||
| variant X1 | 2063 | 9 | 538 | 59.68 | 9.41 | Yes, 24–25 (0.96) | Cell membrane | Cytoplasm (74.22%) | |||
| variant X2 | 2053 | 9 | 538 | 59.68 | 9.41 | Yes, 24–25 (0.96) | Cell membrane | Cytoplasm (74.22%) | |||
| 5 | BvAAO_5 | LOC104895712 | Chromosome 6, NC_025817.2 (10362403–10365259) | 2095 | 5 | 588 | 65.90 | 6.68 | Yes, 35–36 (0.57) | Cell membrane | Cytoplasm (85.23%) |
| 6 | BvAAO_6 | LOC104895596 | Chromosome 6, NC_025817.2 (8771987–8776770) | 2044 | 8 | 537 | 60.78 | 6.71 | Yes, 24–25 (0.78) | Cell membrane | Cytoplasm (87.40%) |
| 7 | BvAAO_7 | LOC104895595 | Chromosome 6, NC_025817.2 (8760298–8765330) | ||||||||
| variant X1 | 2054 | 8 | 542 | 61.25 | 9.06 | Yes, 24–25 (0.93) | Cell membrane | Extracellular space (98.22%) | |||
| variant X2 | 1798 | 8 | 542 | 61.25 | 9.06 | Yes, 24–25 (0.93) | Cell membrane | Extracellular space (98.22%) | |||
| variant X3 | 1884 | 8 | 542 | 61.25 | 9.06 | Yes, 24–25 (0.93) | Cell membrane | Extracellular space (98.22%) | |||
| 8 | BvAAO_8 | LOC104900974 | Chromosome 8, NC_025819.2 (8345583–8354836) | ||||||||
| variant X1 | 1989 | 8 | 548 | 62.61 | 9.84 | Yes, 29–30 (0.92) | Cell membrane | Cytoplasm (75.26%) | |||
| variant X2 | 2024 | 8 | 543 | 62.29 | 9.76 | Yes, 24–25 (0.91) | Cell membrane | Cytoplasm (80.90%) | |||
| 9 | BvAAO_9 | LOC104903081 | Chromosome 9, NC_025820.2 (11718104–11729554) | 2063 | 5 | 588 | 65.60 | 8.55 | No | Cell membrane | Cytoplasm (91.22%) |
| 10 | BvAAO_10 | LOC104903859 | Chromosome 9, NC_025820.2 (32099171–32106493) | 2117 | 8 | 537 | 60.84 | 8.96 | Yes, 22–23 (0.97) | Cell membrane | Cytoplasm (64.34%) |
| 11 | BvAAO_11 | LOC104884409 | NW_017567596.1 (164256–159396) | 2176 | 7 | 539 | 59.82 | 9.40 | Yes, 20–21 (0.95) | Cell membrane | Extracellular space (82.94%) |
| 12 | BvAAO_12 | LOC104884407 | NW_017567596.1 (158898–154368) | 3014 | 7 | 539 | 59.89 | 9.46 | Yes, 20–21 (0.93) | Cell membrane | Secretory Pathway (96.60%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skorupa, M.; Szczepanek, J.; Yolcu, S.; Mazur, J.; Tretyn, A.; Tyburski, J. Characteristic of the Ascorbate Oxidase Gene Family in Beta vulgaris and Analysis of the Role of AAO in Response to Salinity and Drought in Beet. Int. J. Mol. Sci. 2022, 23, 12773. https://doi.org/10.3390/ijms232112773
Skorupa M, Szczepanek J, Yolcu S, Mazur J, Tretyn A, Tyburski J. Characteristic of the Ascorbate Oxidase Gene Family in Beta vulgaris and Analysis of the Role of AAO in Response to Salinity and Drought in Beet. International Journal of Molecular Sciences. 2022; 23(21):12773. https://doi.org/10.3390/ijms232112773
Chicago/Turabian StyleSkorupa, Monika, Joanna Szczepanek, Seher Yolcu, Justyna Mazur, Andrzej Tretyn, and Jarosław Tyburski. 2022. "Characteristic of the Ascorbate Oxidase Gene Family in Beta vulgaris and Analysis of the Role of AAO in Response to Salinity and Drought in Beet" International Journal of Molecular Sciences 23, no. 21: 12773. https://doi.org/10.3390/ijms232112773
APA StyleSkorupa, M., Szczepanek, J., Yolcu, S., Mazur, J., Tretyn, A., & Tyburski, J. (2022). Characteristic of the Ascorbate Oxidase Gene Family in Beta vulgaris and Analysis of the Role of AAO in Response to Salinity and Drought in Beet. International Journal of Molecular Sciences, 23(21), 12773. https://doi.org/10.3390/ijms232112773