
The Ring World: Eversion of Small Double-Stranded Polynucleotide Circlets at the Origin of DNA Double Helix, RNA Polymerization, Triplet Code, Twenty Amino Acids, and Strand Asymmetry



{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
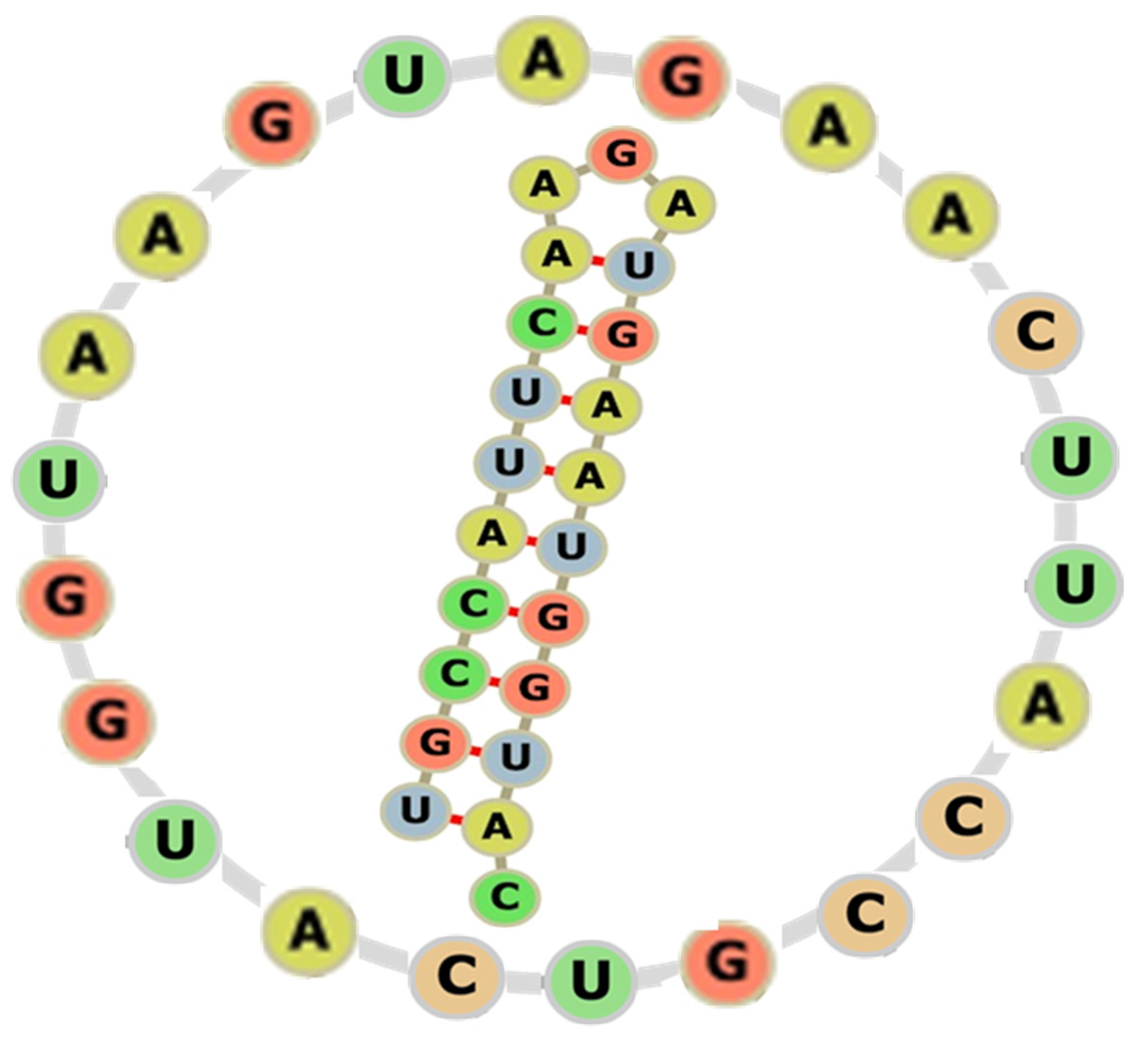
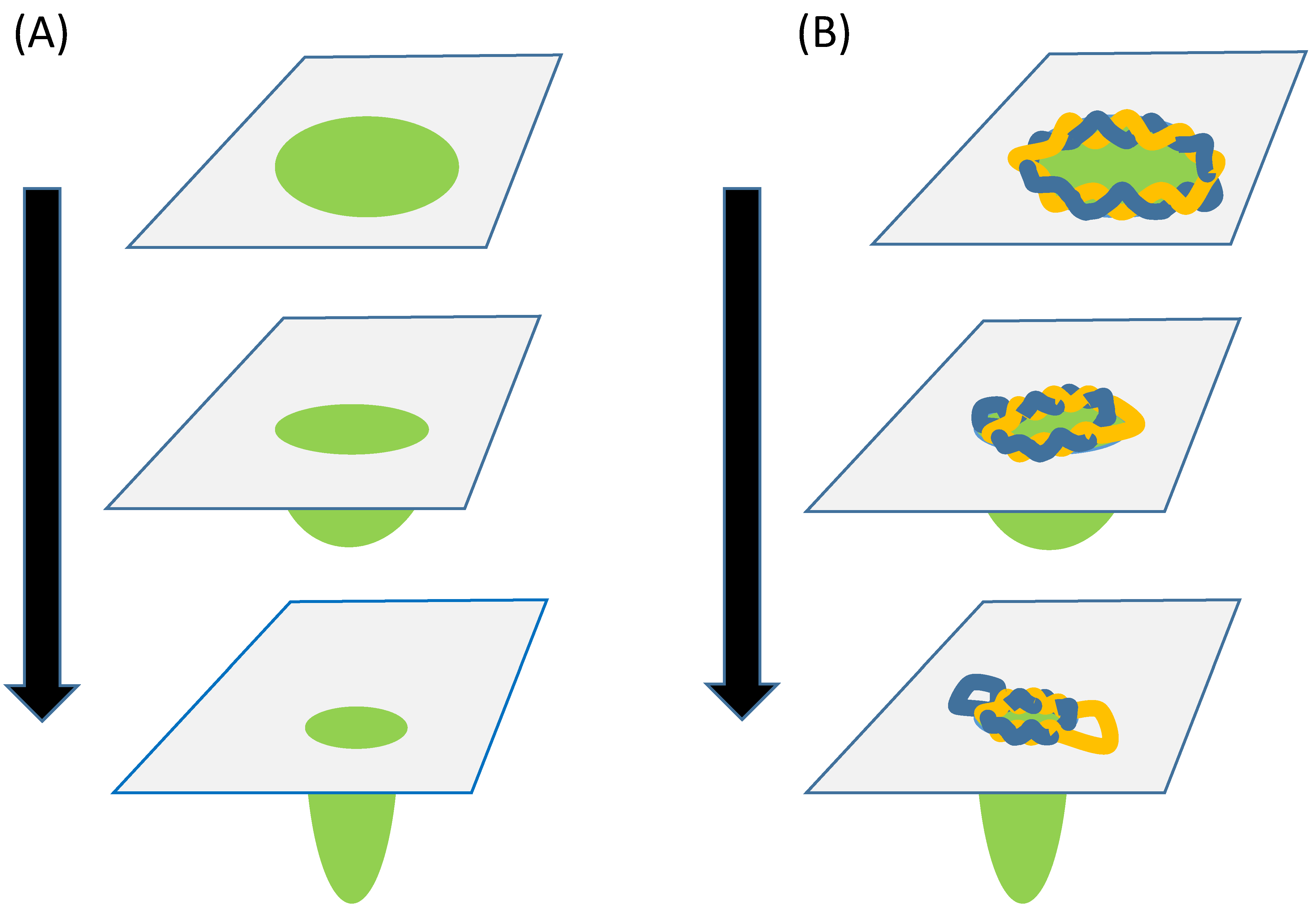
2. The Circlet Hypothesis
3. Evidence
3.1. Creation of Small RNA and DNA Circles
3.2. Topological Strains
3.3. Catalysis by Polynucleotides
3.4. Strand Asymmetry
3.5. Polynucleotide Networks
3.6. Lipid Interactions
4. Testing the Hypothesis
4.1. The Principal Prediction
4.2. A Limited Prediction
4.3. A Specific Prediction
4.4. A More General Prediction
4.5. A Highly Speculative Prediction
4.6. The Assembly of Circlets
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wagner, I.; Musso, H. New naturally occurring amino acids. Angew. Chem. Int. Ed. Engl. 1983, 22, 816–828. [Google Scholar] [CrossRef]
- Meringer, M.; Cleaves, H.J., 2nd; Freeland, S.J. Beyond terrestrial biology: Charting the chemical universe of alpha-amino acid structures. J. Chem. Inf. Model. 2013, 53, 2851–2862. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Lozoya-Colinas, A.; Gallego, I.; Grover, M.A.; Hud, N.V. Solvent viscosity facilitates replication and ribozyme catalysis from an RNA duplex in a model prebiotic process. Nucleic Acids Res. 2019, 47, 6569–6577. [Google Scholar] [CrossRef]
- Jerome, C.A.; Kim, H.J.; Mojzsis, S.J.; Benner, S.A.; Biondi, E. Catalytic Synthesis of Polyribonucleic Acid on Prebiotic Rock Glasses. Astrobiology 2022, 22, 629–636. [Google Scholar] [CrossRef] [PubMed]
- Raine, D.J.; Norris, V. Lipid domain boundaries as prebiotic catalysts of peptide bond formation. J. Theor. Biol. 2007, 246, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Olasagasti, F.; Rajamani, S. Lipid-assisted polymerization of nucleotides. Life 2019, 9, 83. [Google Scholar] [CrossRef] [Green Version]
- Hassenkam, T.; Damer, B.; Mednick, G.; Deamer, D. Afm images of viroid-sized rings that self-assemble from mononucleotides through wet-dry cycling: Implications for the origin of life. Life 2020, 10, 321. [Google Scholar] [CrossRef]
- Diener, T.O. Viroids: “Living fossils” of primordial RNAs? Biol. Direct 2016, 11, 15. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Ding, D.; Szostak, J.W. The virtual circular genome model for primordial RNA replication. RNA 2021, 27, 1–11. [Google Scholar] [CrossRef]
- Demongeot, J.; Seligmann, H. Codon assignment evolvability in theoretical minimal RNA rings. Gene 2021, 769, 145208. [Google Scholar] [CrossRef]
- Segre, D.; Ben-Eli, D.; Lancet, D. Compositional genomes: Prebiotic information transfer in mutually catalytic noncovalent assemblies. Proc. Natl. Acad. Sci. USA 2000, 97, 4112–4117. [Google Scholar] [CrossRef] [Green Version]
- Hunding, A.; Kepes, F.; Lancet, D.; Minsky, A.; Norris, V.; Raine, D.; Sriram, K.; Root-Bernstein, R. Compositional complementarity and prebiotic ecology in the origin of life. Bioessays 2006, 28, 399–412. [Google Scholar] [CrossRef]
- Norris, V.; Loutelier-Bourhis, C.; Thierry, A. How did metabolism and genetic replication get married? Orig. Life Evol. Biosph. 2012, 42, 487–495. [Google Scholar] [CrossRef]
- Yin, S.; Kiong Ho, C.; Miller, E.S.; Shuman, S. Characterization of bacteriophage kvp40 and t4 RNA ligase 2. Virology 2004, 319, 141–151. [Google Scholar] [CrossRef] [Green Version]
- Cheng, K.; An, R.; Cui, Y.; Zhang, Y.; Han, X.; Sui, Z.; Chen, H.; Liang, X.; Komiyama, M. Rna ligation of very small pseudo nick structures by t4 RNA ligase 2, leading to efficient production of versatile RNA rings. RSC Adv. 2019, 9, 8620–8627. [Google Scholar] [CrossRef] [Green Version]
- Demongeot, J.; Moreira, A. A possible circular RNA at the origin of life. J. Theor. Biol. 2007, 249, 314–324. [Google Scholar] [CrossRef]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature 2013, 495, 333–338. [Google Scholar] [CrossRef]
- Liu, C.X.; Chen, L.L. Circular RNAs: Characterization, cellular roles, and applications. Cell 2022, 185, 2016–2034. [Google Scholar] [CrossRef]
- Wei, X.; Li, H.; Yang, J.; Hao, D.; Dong, D.; Huang, Y.; Lan, X.; Plath, M.; Lei, C.; Lin, F.; et al. Circular RNA profiling reveals an abundant circlmo7 that regulates myoblasts differentiation and survival by sponging mir-378a-3p. Cell Death Dis. 2017, 8, e3153. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Liang, D.; Chen, C.; Wang, Y.; Amu, G.; Yang, J.; Yu, L.; Dmochowski, I.J.; Tang, X. Circular siRNAs for reducing off-target effects and enhancing long-term gene silencing in cells and mice. Mol. Ther. Nucleic Acids 2018, 10, 237–244. [Google Scholar] [CrossRef]
- Rechendorff, K.; Witz, G.; Adamcik, J.; Dietler, G. Persistence length and scaling properties of single-stranded DNA adsorbed on modified graphite. J. Chem. Phys. 2009, 131, 095103. [Google Scholar] [CrossRef]
- Roth, E.; Glick Azaria, A.; Girshevitz, O.; Bitler, A.; Garini, Y. Measuring the conformation and persistence length of single-stranded DNA using a DNA origami structure. Nano Lett. 2018, 18, 6703–6709. [Google Scholar] [CrossRef] [PubMed]
- Gubu, A.; Wang, J.; Jin, H.; Tang, X. Synthesis and “DNA interlocks” formation of small circular oligodeoxynucleotides. ACS Appl. Mater. Interfaces 2020, 12, 12584–12590. [Google Scholar] [CrossRef] [PubMed]
- Vafabakhsh, R.; Ha, T. Extreme bendability of DNA less than 100 base pairs long revealed by single-molecule cyclization. Science 2012, 337, 1097–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumann, C.G.; Smith, S.B.; Bloomfield, V.A.; Bustamante, C. Ionic effects on the elasticity of single DNA molecules. Proc. Natl. Acad. Sci. USA 1997, 94, 6185–6190. [Google Scholar] [CrossRef] [Green Version]
- Vologodskii, A.; Du, Q.; Frank-Kamenetskii, M.D. Bending of short DNA helices. Artif. DNA PNA XNA 2013, 4, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Wang, I.X.; Grunseich, C.; Fox, J.; Burdick, J.; Zhu, Z.; Ravazian, N.; Hafner, M.; Cheung, V.G. Human proteins that interact with RNA/DNA hybrids. Genome Res. 2018, 28, 1405–1414. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Liu, W.; Deng, X.; Xie, W.; Chen, H.; Zhong, Z.; Ma, J. Gc-content dependence of elastic and overstretching properties of DNA:RNA hybrid duplexes. Biophys. J. 2020, 119, 852–861. [Google Scholar] [CrossRef]
- Zhang, C.; Fu, H.; Yang, Y.; Zhou, E.; Tan, Z.; You, H.; Zhang, X. The mechanical properties of RNA-DNA hybrid duplex stretched by magnetic tweezers. Biophys. J. 2019, 116, 196–204. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Meisburger, S.P.; Pabit, S.A.; Sutton, J.L.; Webb, W.W.; Pollack, L. Ionic strength-dependent persistence lengths of single-stranded RNA and DNA. Proc. Natl. Acad. Sci. USA 2012, 109, 799–804. [Google Scholar] [CrossRef]
- Pal, A.; Levy, Y. Structure, stability and specificity of the binding of ssDNA and ssRNA with proteins. PLoS Comput. Biol. 2019, 15, e1006768. [Google Scholar] [CrossRef]
- Besson, J. Topology of circular DNA. Ann. Génét. 1977, 20, 145–152. [Google Scholar]
- Megalathan, A.; Cox, B.D.; Wilkerson, P.D.; Kaur, A.; Sapkota, K.; Reiner, J.E.; Dhakal, S. Single-molecule analysis of i-motif within self-assembled DNA duplexes and nanocircles. Nucleic Acids Res. 2019, 47, 7199–7212. [Google Scholar] [CrossRef] [Green Version]
- Pyne, A.L.B.; Noy, A.; Main, K.H.S.; Velasco-Berrelleza, V.; Piperakis, M.M.; Mitchenall, L.A.; Cugliandolo, F.M.; Beton, J.G.; Stevenson, C.E.M.; Hoogenboom, B.W.; et al. Base-pair resolution analysis of the effect of supercoiling on DNA flexibility and major groove recognition by triplex-forming oligonucleotides. Nat. Commun. 2021, 12, 1053. [Google Scholar] [CrossRef]
- Bates, A.D.; Maxwell, A. DNA Topology; Oxford University Press: Oxford, UK, 2005. [Google Scholar]
- Du, Q.; Kotlyar, A.; Vologodskii, A. Kinking the double helix by bending deformation. Nucleic Acids Res. 2008, 36, 1120–1128. [Google Scholar] [CrossRef] [Green Version]
- Lankas, F.; Lavery, R.; Maddocks, J.H. Kinking occurs during molecular dynamics simulations of small DNA minicircles. Structure 2006, 14, 1527–1534. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, J.S.; Laughton, C.A.; Harris, S.A. Atomistic simulations reveal bubbles, kinks and wrinkles in supercoiled DNA. Nucleic Acids Res. 2011, 39, 3928–3938. [Google Scholar] [CrossRef] [Green Version]
- Lionberger, T.A.; Demurtas, D.; Witz, G.; Dorier, J.; Lillian, T.; Meyhofer, E.; Stasiak, A. Cooperative kinking at distant sites in mechanically stressed DNA. Nucleic Acids Res. 2011, 39, 9820–9832. [Google Scholar] [CrossRef] [Green Version]
- Alexiou, T.S.; Alatas, P.V.; Tsalikis, D.G.; Mavrantzas, V.G. Conformational and dynamic properties of short DNA minicircles in aqueous solution from atomistic molecular dynamics simulations. Macromolecules 2020, 53, 5903–5918. [Google Scholar] [CrossRef]
- Breaker, R.R. DNA aptamers and DNA enzymes. Curr. Opin. Chem. Biol. 1997, 1, 26–31. [Google Scholar] [CrossRef]
- Cuenoud, B.; Szostak, J.W. A DNA metalloenzyme with DNA ligase activity. Nature 1995, 375, 611–614. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Kusumaatmaja, H.; Lipowsky, R.; Dimova, R. Wetting-induced budding of vesicles in contact with several aqueous phases. J. Phys. Chem. B 2012, 116, 1819–1823. [Google Scholar] [CrossRef] [PubMed]
- Kristoffersen, E.L.; Burman, M.; Noy, A.; Holliger, P. Rolling circle RNA synthesis catalyzed by RNA. eLife 2022, 11, e75186. [Google Scholar] [CrossRef] [PubMed]
- Root-Bernstein, R. Simultaneous origin of homochirality, the genetic code and its directionality. Bioessays 2007, 29, 689–698. [Google Scholar] [CrossRef] [PubMed]
- Lipowsky, R.; Dimova, R. Domains in membranes and vesicles. J. Phys. Condens. Matter 2002, 15, S31–S45. [Google Scholar] [CrossRef] [Green Version]
- Wongsirojkul, N.; Shimokawa, N.; Opaprakasit, P.; Takagi, M.; Hamada, T. Osmotic-tension-induced membrane lateral organization. Langmuir 2020, 36, 2937–2945. [Google Scholar] [CrossRef]
- Johnson, D.B.; Wang, L. Imprints of the genetic code in the ribosome. Proc. Natl. Acad. Sci. USA 2010, 107, 8298–8303. [Google Scholar] [CrossRef] [Green Version]
- Trifonov, E.N. The triplet code from first principles. J. Biomol. Struct. Dyn. 2004, 22, 1–11. [Google Scholar] [CrossRef]
- Polyansky, A.A.; Zagrovic, B. Evidence of direct complementary interactions between messenger RNAs and their cognate proteins. Nucleic Acids Res. 2013, 41, 8434–8443. [Google Scholar] [CrossRef]
- Demongeot, J.; Seligmann, H. Theoretical minimal RNA rings mimick molecular evolution before tRNA-mediated translation: Codon-amino acid affinities increase from early to late RNA rings. CRAS Biol. 2020, 343, 111–122. [Google Scholar] [CrossRef]
- Demongeot, J.; Seligmann, H. Comparisons between small ribosomal RNA and theoretical minimal RNA ring secondary structures confirm phylogenetic and structural accretion histories. Sci. Rep. 2020, 10, 7693. [Google Scholar] [CrossRef]
- Demongeot, J.; Moreira, A.; Seligmann, H. Negative CG dinucleotide bias: An explanation based on feedback loops between Arginine codon assignments and theoretical minimal RNA rings. Bioessays 2021, 43, 2000071. [Google Scholar] [CrossRef]
- Tamura, K.; Schimmel, P. Peptide synthesis with a template-like RNA guide and aminoacyl phosphate adaptors. Proc. Natl. Acad. Sci. USA 2003, 100, 8666–8669. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; Wang, Y. Protein-free ribosomal RNA scaffolds can assemble poly-lysine oligos from charged tRNA fragments. Biochem. Biophys. Res. Commun. 2021, 544, 81–85. [Google Scholar] [CrossRef]
- Li, J.; Mohammed-Elsabagh, M.; Paczkowski, F.; Li, Y. Circular nucleic acids: Discovery, functions and applications. ChemBiochem 2020, 21, 1547–1566. [Google Scholar] [CrossRef]
- Bose, T.; Fridkin, G.; Davidovich, C.; Krupkin, M.; Dinger, N.; Falkovich, A.H.; Peleg, Y.; Agmon, I.; Bashan, A.; Yonath, A. Origin of life: Protoribosome forms peptide bonds and links RNA and protein dominated worlds. Nucleic Acids Res. 2022, 50, 1815–1828. [Google Scholar] [CrossRef]
- Bhaduri, S.; Ranjan, N.; Arya, D.P. An overview of recent advances in duplex DNA recognition by small molecules. Beilstein J. Org. Chem. 2018, 14, 1051–1086. [Google Scholar] [CrossRef]
- Sinden, R.R.; Carlson, J.O.; Pettijohn, D.E. Torsional tension in the DNA double helix measured with trimethylpsoralen in living E. coli cells: Analogous measurements in insect and human cells. Cell 1980, 21, 773–783. [Google Scholar] [CrossRef]
- Conan, M.; Théret, N.; Langouet, S.; Siegel, A. Constructing xenobiotic maps of metabolism to predict enzymes catalyzing metabolites capable of binding to DNA. BMC Bioinform. 2021, 22, 450. [Google Scholar] [CrossRef]
- Thomas, R. Recherches sur la dénaturation des acides désoxyribonucléiques. Biochim. Biophys. Acta 1954, 14, 231–240. [Google Scholar] [CrossRef]
- Diener, T.O. Circular RNAs: Relics of precellular evolution? Proc. Natl. Acad. Sci. USA 1989, 86, 9370–9374. [Google Scholar] [CrossRef] [PubMed]
- Ivica, N.A.; Obermayer, B.; Campbell, G.W.; Rajamani, S.; Gerland, U.; Chen, I.A. The paradox of dual roles in the RNA world: Resolving the conflict between stable folding and templating ability. J. Mol. Evol. 2013, 77, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Szathmary, E.; Smith, J.M. The evolution of chromosomes. Ii. Molecular mechanisms. J. Theor. Biol. 1993, 164, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, N.; Hogeweg, P.; Kaneko, K. The origin of a primordial genome through spontaneous symmetry breaking. Nat. Commun. 2017, 8, 250. [Google Scholar] [CrossRef] [Green Version]
- Rocha, E.P.; Fralick, J.; Vediyappan, G.; Danchin, A.; Norris, V. A strand-specific model for chromosome segregation in bacteria. Mol. Microbiol. 2003, 49, 895–903. [Google Scholar] [CrossRef] [PubMed]
- Gangwe Nana, G.Y.; Ripoll, C.; Cabin-Flaman, A.; Gibouin, D.; Delaune, A.; Janniere, L.; Grancher, G.; Chagny, G.; Loutelier-Bourhis, C.; Lentzen, E.; et al. Division-based, growth rate diversity in bacteria. Front. Microbiol. 2018, 9, 849. [Google Scholar] [CrossRef]
- Norris, V.; Ripoll, C. Generation of bacterial diversity by segregation of DNA strands. Front. Microbiol. 2021, 12, 550856. [Google Scholar] [CrossRef]
- Klar, A.J. Developmental choices in mating-type interconversion in fission yeast. Trends Genet. 1992, 8, 208–213. [Google Scholar] [CrossRef]
- Levin, M.; Klar, A.J.; Ramsdell, A.F. Introduction to provocative questions in left-right asymmetry. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371, 20150399. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; O’Hagan, M.P.; Neumann, E.; Nechushtai, R.; Willner, I. Integration of photocatalytic and dark-operating catalytic biomimetic transformations through DNA-based constitutional dynamic networks. Nat. Commun. 2021, 12, 4224. [Google Scholar] [CrossRef]
- Szabat, M.; Kierzek, R. Parallel-stranded DNA and RNA duplexes—Structural features and potential applications. FEBS J. 2017, 284, 3986–3998. [Google Scholar] [CrossRef]
- Budin, I.; Szostak, J.W. Expanding roles for diverse physical phenomena during the origin of life. Annu. Rev. Biophys. 2010, 39, 245–263. [Google Scholar] [CrossRef] [Green Version]
- Deamer, D. The role of lipid membranes in life’s origin. Life 2017, 7, 5. [Google Scholar] [CrossRef]
- Paecht-Horowitz, M.; Berger, J.; Katchalsky, A. Prebiotic synthesis of polypeptides by heterogeneous polycondensation of amino-acid adenylates. Nature 1970, 228, 636–639. [Google Scholar] [CrossRef]
- Legent, G.; Delaune, A.; Norris, V.; Delcorte, A.; Gibouin, D.; Lefebvre, F.; Misevic, G.; Thellier, M.; Ripoll, C. Method for macromolecular colocalization using atomic recombination in dynamic sims. J. Phys. Chem. B 2008, 112, 5534–5546. [Google Scholar] [CrossRef]
- Basu, A.; Bobrovnikov, D.G.; Qureshi, Z.; Kayikcioglu, T.; Ngo, T.T.M.; Ranjan, A.; Eustermann, S.; Cieza, B.; Morgan, M.T.; Hejna, M.; et al. Measuring DNA mechanics on the genome scale. Nature 2021, 589, 462–467. [Google Scholar] [CrossRef]
- Cabin-Flaman, A.; Monnier, A.F.; Coffinier, Y.; Audinot, J.N.; Gibouin, D.; Wirtz, T.; Boukherroub, R.; Migeon, H.N.; Bensimon, A.; Janniere, L.; et al. Combed single DNA molecules imaged by secondary ion mass spectrometry. Anal. Chem. 2011, 83, 6940–6947. [Google Scholar] [CrossRef]
- Kauffman, S.A. Investigations; Oxford University Press: Oxford, UK, 2000. [Google Scholar]
- Montevil, M.; Mossio, M. Biological organisation as closure of constraints. J. Theor. Biol. 2015, 372, 179–191. [Google Scholar] [CrossRef] [Green Version]
- Dwyer, D.S. Amino acid sequence homology between ligands and their receptors: Potential identification of binding sites. Life Sci. 1989, 45, 421–429. [Google Scholar] [CrossRef]
- Root-Bernstein, R.S.; Dillon, P.F. Molecular complementarity I: The complementarity theory of the origin and evolution of life. J. Theor. Biol. 1997, 188, 447–479. [Google Scholar] [CrossRef]
- Norris, V.; Onoda, T.; Pollaert, H.; Grehan, G. The mechanical advantages of DNA. Biosystems 1999, 49, 71–78. [Google Scholar] [CrossRef]
- Levintov, L.; Paul, S.; Vashisth, H. Reaction coordinate and thermodynamics of base flipping in RNA. J. Chem. Theory Comput. 2021, 17, 1914–1921. [Google Scholar] [CrossRef]
- Gavette, J.V.; Stoop, M.; Hud, N.V.; Krishnamurthy, R. Rna-DNA chimeras in the context of an RNA world transition to an RNA/DNA world. Angew. Chem. Int. Ed. Engl. 2016, 55, 13204–13209. [Google Scholar] [CrossRef] [PubMed]
- Bhowmik, S.; Krishnamurthy, R. The role of sugar-backbone heterogeneity and chimeras in the simultaneous emergence of RNA and DNA. Nat. Chem. 2019, 11, 1009–1018. [Google Scholar] [CrossRef] [PubMed]
- Norris, V.; Reusch, R.N.; Igarashi, K.; Root-Bernstein, R. Molecular complementarity between simple, universal molecules and ions limited phenotype space in the precursors of cells. Biol. Direct 2015, 10, 28. [Google Scholar] [CrossRef] [Green Version]
- Norris, V.; Woldringh, C.; Mileykovskaya, E. A hypothesis to explain division site selection in escherichia coli by combining nucleoid occlusion and min. FEBS Lett. 2004, 561, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Lafontaine, C.; Valleton, J.M.; Orange, N.; Norris, V.; Mileykovskaya, E.; Alexandre, S. Behaviour of bacterial division protein FtsZ under a monolayer with phospholipid domains. Biochim. Biophys. Acta 2007, 1768, 2812–2821. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Norris, V.; Demongeot, J. The Ring World: Eversion of Small Double-Stranded Polynucleotide Circlets at the Origin of DNA Double Helix, RNA Polymerization, Triplet Code, Twenty Amino Acids, and Strand Asymmetry. Int. J. Mol. Sci. 2022, 23, 12915. https://doi.org/10.3390/ijms232112915
Norris V, Demongeot J. The Ring World: Eversion of Small Double-Stranded Polynucleotide Circlets at the Origin of DNA Double Helix, RNA Polymerization, Triplet Code, Twenty Amino Acids, and Strand Asymmetry. International Journal of Molecular Sciences. 2022; 23(21):12915. https://doi.org/10.3390/ijms232112915
Chicago/Turabian StyleNorris, Victor, and Jacques Demongeot. 2022. "The Ring World: Eversion of Small Double-Stranded Polynucleotide Circlets at the Origin of DNA Double Helix, RNA Polymerization, Triplet Code, Twenty Amino Acids, and Strand Asymmetry" International Journal of Molecular Sciences 23, no. 21: 12915. https://doi.org/10.3390/ijms232112915
APA StyleNorris, V., & Demongeot, J. (2022). The Ring World: Eversion of Small Double-Stranded Polynucleotide Circlets at the Origin of DNA Double Helix, RNA Polymerization, Triplet Code, Twenty Amino Acids, and Strand Asymmetry. International Journal of Molecular Sciences, 23(21), 12915. https://doi.org/10.3390/ijms232112915