

Targeting Immune Senescence in Atherosclerosis
 ,
,  , ,
, ,  and
and 
{kind=link}
Abstract
:1. Introduction
2. Pathophysiology of Atherosclerosis
3. The Immune System in Atherosclerosis
4. Immunosenescence in Atherosclerosis
5. Features of Cellular Senescence
5.1. Morphological Changes
5.2. Cell Surface Markers Expression
5.2.1. Intracellular Adhesion Molecule-1 (ICAM-1)
5.2.2. CD36
5.2.3. Urokinase-Type Plasminogen Activator Receptor (uPAR)
5.2.4. Beta-2 Microglobulin (B2MG or B2M)
5.2.5. Dipeptidyl-Peptidase 4 (DPP4 or CD26)
5.3. Cellular Metabolism and Secretory Phenotype
5.3.1. Increased Metabolic Activity
5.3.2. Senescence-Associated Secretory Phenotype (SASP)
5.4. Lysosomal Activity
5.4.1. Lysosomal Alteration
5.4.2. SA β-Galactosidase (SA-βgal) Activity
5.4.3. The Mammalian Target of Rapamycin (mTOR) Activity
5.5. Cell Cycle Arrest and Apoptosis Resistance
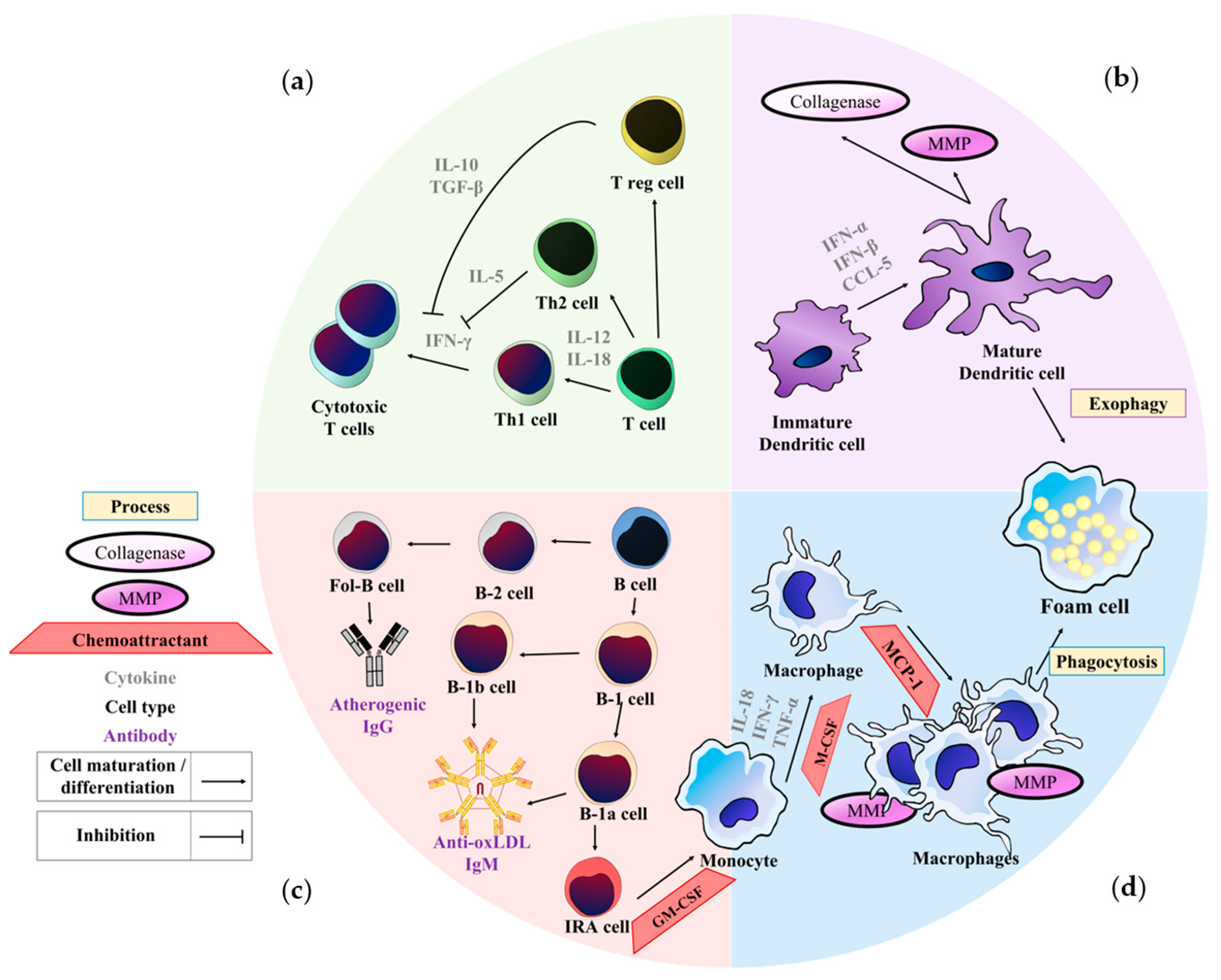
6. Regulation of Immune Cells Senescence in Atherosclerosis
6.1. Monocytes, Macrophages, and Foam Cells
6.2. Dendritic Cells (DCs)
6.3. T-Lymphocytes
6.4. B-Lymphocytes
7. Therapeutic Strategies Targeting Immune Senescent Cells in Atherosclerosis
8. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 4EBP1 | 4E binding protein 1 |
| ADA | adenosine deaminase |
| AMP | adenosine monophosphate |
| AMPK | adenosine monophosphate-activated protein kinase |
| AP-1 | activator protein 1 |
| APC | antigen presenting cell |
| ApoE | apolipoprotein E |
| ARF | alternative reading frame |
| Atg | autophagy related gene |
| ATM | ataxia-telangiectasia mutated |
| ATP | adenosine triphosphate |
| ATR | RAD3-related |
| B2MG | beta-2 microglobulin |
| BCL | B-Cell lymphoma |
| BCR | B-Cell receptor |
| CDC | conventional dendritic cell |
| Cdk2 | cyclin-dependent kinase 2 |
| CDKN1A | cyclin dependent kinase inhibitor 1A |
| c-H2AX | phosphorylated H2A histone family member X |
| CVD | cardiovascular disease |
| DAMP | damage-associated molecular pattern |
| DC | dendritic cell |
| DDR | deoxyribonucleic acid damage response |
| DNA | deoxyribonucleic acid |
| DNA-SCARS | sequence characterized amplified region |
| DPP4 | dipeptidyl-peptidase 4 |
| DRP1 | dynamin-related protein 1 |
| E2F | E2 factor |
| ECM | extracellular matrix |
| EC | endothelial cell |
| ETC | electron transport chain |
| Foxn1 | forkhead box n1 |
| FOXO1 | forkhead box transcription factor 1 |
| GIP | glucose-dependent insulinotropic polypeptide |
| GLP | glucagon-like peptide |
| GM-CSF | granulocyte-macrophage colony-stimulating factor |
| GRO1 | growth regulated alpha protein 1 |
| HDL | high density lipoprotein |
| HP1 | heterochromatin protein 1 |
| hsCRP | high-sensitivity C-reactive protein |
| HVT-SM1 | human fibroblast and human vascular smooth muscle |
| ICAM-1 | intercellular adhesion molecule 1 |
| IFN | interferon |
| Ig | immunoglobulin |
| IL | interleukin |
| IMT | carotid-intima-media thickness |
| IRA | innate response activator |
| IRAK | interleukin-1 receptor-associated kinase 1 |
| LDL | low density lipoprotein |
| LFA-1 | lymphocyte function associated antigen-1 |
| LOX-1 | lectin-like oxidized low density lipoprotein |
| LSD | lysosomal storage disease |
| LXR | liver X receptors |
| M1 | pro-inflammatory monocytes |
| MAPK | mitogen-activated protein kinase |
| MAPKAPK2 | MAP kinase–activated protein kinase 2 |
| MCP-1 | monocyte chemoattractant protein-1 |
| M-CSF | monocyte-colony stimulating factor |
| MHC | major histocompatibility complex |
| miR-33 | microRNA- 33 |
| Mitf | Melanocyte-inducing transcription factor |
| MMP | matrix metalloprotease |
| MODC | monocyte-derived dendritic cell |
| mt | mitochondria |
| mtDAMP | mitochondrial damage associated molecular pattern |
| mTORC1 | mammalian target of rapamycin complex 1 |
| mTORC1 | mammalian target of rapamycin complex 1 |
| mtROS | mitochondrial reactive oxygen species |
| NFAT | nuclear factor of activated T-cells |
| NF-κB | nuclear factor kappa-light-chain-enhancer of activated B cells |
| NADPH | nicotinamide adenine dinucleotide phosphate |
| NOX | nicotinamide adenine dinucleotide phosphate oxidase |
| NK | natural killer |
| NLRP3 | NLR Family Pyrin Domain Containing 3 |
| NO | nitric oxide |
| OxLDL | oxidized low-density lipoprotein |
| p53 | tumor suppressor gene p53 |
| PAI-1 | plasminogen activator inhibitor-1 |
| PBMC | peripheral blood mononuclear cells |
| PDGF | platelet-derived growth factor |
| PML | promyelocytic leukemia |
| PMN | polymorphonuclear cells |
| Rac1 | Rac Family Small GTPase 1 |
| RIG-1 | retinoic acid-inducible gene I |
| RNS | reactive nitrogen species |
| ROS | reactive oxygen species |
| S6K1 | S6 kinase 1 |
| SAHFs | senescence-associated heterochromatin foci |
| SAMP8 | senescence accelerated-prone 8 |
| SASP | senescence-associated secretory phenotype |
| SA-β-gal | SA β-galactosidase |
| Ser | serine |
| SIRT | sirtuin |
| SNP | single nucleotide polymorphism |
| STAT1 | signal transducer and activator of transcription 1 |
| suPAR | soluble uPAR |
| TCR | T-cell receptor |
| TEC | thymic epithelial cell |
| TFEB | Transcription factor EB |
| TEMRA | terminally differentiated effector memory cells |
| TET2 | TET methylcytosine dioxygenase 2 |
| TLR | toll-like receptors |
| TNF-α | tumor necrosis alpha |
| TRAF | tumor necrosis factor receptor-associated factor |
| Treg | Regulatory T cell |
| TREM | triggering receptor expressed on myeloid cells |
| TWEAK | tumor-necrosis factor-like weak inducers of apoptosis |
| Ulk1 | Unc-51-like Kinase 1 |
| uPAR | urokinase-type plasminogen activator receptor |
| VCAM-1 | vascular cell adhesion molecule 1 |
| VECs | vascular endothelial cells |
| VEGF | vascular endothelial growth factor |
| VSMC | vascular smooth muscle cell |
| XO | xanthine oxidase |
| ZFP36L1 | zinc-finger RNA-binding protein |
References
- World Health Organization. Cardiovascular Diseases (CVDs). 2021. Available online: https://www.who.int (accessed on 28 February 2022).
- Rocamora-Reverte, L.; Melzer, F.L.; Würzner, R.; Weinberger, B. The Complex Role of Regulatory T Cells In Immunity And Aging. Front. Immunol. 2021, 11, 616949. [Google Scholar] [CrossRef] [PubMed]
- Bektas, A.; Schurman, S.H.; Sen, R.; Ferrucci, L. Human T Cell Immunosenescence and Inflammation In Aging. J. Leukoc. Biol. 2017, 102, 977–988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robbesyn, F.; Salvayre, R.; Negre-Salvayre, A. Dual Role of Oxidized LDL On The NF-Kappab Signaling Pathway. Free Radic. Res. 2004, 38, 541–551. [Google Scholar] [CrossRef] [PubMed]
- Collins, C.; Ellie, T. Hemodynamic Forces in Endothelial Dysfunction and Vascular Aging. Exp. Gerontol. 2011, 46, 185–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawai, T.; Akira, S. TLR Signaling. Cell Death Differ. 2006, 13, 816–825. [Google Scholar] [CrossRef] [Green Version]
- Badimon, L.; Vilahur, G. Thrombosis Formation On Atherosclerotic Lesions And Plaque Rupture. J. Intern. Med. 2014, 276, 618–632. [Google Scholar] [CrossRef]
- Wang, J.C.; Bennett, M. Aging and Atherosclerosis. Circ. Res. 2012, 111, 245–259. [Google Scholar] [CrossRef] [Green Version]
- Barbulescu, K.; Becker, C.; Schalaal, J.F.; Schmitt, E.; Büschenfelde, K.H.M.; Neurath, M.F. Cutting Edge: IL-12 and IL-18 Differentially Regulate the Transcriptional Activity of the Human IFN-γ Promoter in Primary CD+ T Lymphocytes. J. Immunol. 1998, 160, 3642–3647. [Google Scholar]
- Whitman, S.C.; Ravisankar, P.; Daugherty, A. Interleukin-18 Enhances Atherosclerosis In Apolipoprotein E −/− Mice Through Release Of Interferon-γ. Circ. Res. 2002, 90, e34–e38. [Google Scholar] [CrossRef] [Green Version]
- Yasuda, K.; Nakanishi, K.; Tsutsui, H. Interleukin-18 in Health and Disease. Int. J. Mol. Sci. 2019, 20, 649. [Google Scholar] [CrossRef] [Green Version]
- Janoudi, A.; Shamoun, F.E.; Kalavakunta, J.K.; Abela, G.S. Cholesterol Crystal Induced Arterial Inflammation and Destabilization Of Atherosclerotic Plaque. Eur. Heart J. 2015, 37, 1959–1967. [Google Scholar] [CrossRef] [PubMed]
- Shirai, R.; Sato, K.; Yamashita, T.; Yamaguchi, M.; Okano, T.; Watanabe-Kominato, K.; Watanabe, R.; Matsuyama, T.; Ishibashi-Ueda, H.; Koba, S.; et al. Neopterin Counters Vascular Inflammation and Atherosclerosis. J. Am. Heart Assoc. 2018, 7, e007359. [Google Scholar] [CrossRef] [Green Version]
- Hansson, G.; Hermansson, A. The immune system in atherosclerosis. Nat. Immunol. 2011, 12, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.S.; Yen, H.C.; Pan, C.C.; Chau, L.Y. The Role of Interleukin 12 In The Development Of Atherosclerosis In Apoe-Deficient Mice. Arterioscler. Thromb. Vasc. Biol. 1999, 19, 734–742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davenport, P.; Tipping, P.G. The Role of Interleukin-4 and Interleukin-12 in the Progression of Atherosclerosis in Apolipoprotein E-Deficient Mice. Am. J. Pathol. 2003, 163, 1117–1125. [Google Scholar] [CrossRef] [Green Version]
- Hauer, A.; Uyttenhove, C.; de Vos, P.; Stroobant, V.; Renauld, J.-C.; van Berkel, T.; van Snick, J.; Kuiper, J. Blockade of Interleukin-12 Function By Protein Vaccination Attenuates Atherosclerosis. Circulation 2005, 112, 1054–1062. [Google Scholar] [CrossRef] [Green Version]
- Haka, A.; Singh, R.; Grosheva, I.; Hoffner, H.; Capetillo-Zarate, E.; Chin, H.; Anandasabapathy, N.; Maxfield, F. Monocyte-Derived Dendritic Cells Upregulate Extracellular Catabolism of Aggregated Low-Density Lipoprotein on Maturation, Leading to Foam Cell Formation. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 2092–2103. [Google Scholar] [CrossRef] [Green Version]
- Goossens, P.; Gijbels, M.; Zernecke, A.; Eijgelaar, W.; Vergouwe, M.; van der Made, I.; Vanderlocht, J.; Beckers, L.; Buurman, W.; Daemen, M.; et al. Myeloid Type I Interferon Signaling Promotes Atherosclerosis by Stimulating Macrophage Recruitment to Lesions. Cell Metab. 2010, 12, 142–153. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, M.; Tabas, I. Dendritic cells in atherosclerosis. Semin. Immunopathol. 2013, 36, 93–102. [Google Scholar] [CrossRef]
- Ilhan, F.; Kalkanli, S.T. Atherosclerosis and the role of immune cells. World J. Clin. Cases 2015, 3, 345. [Google Scholar] [CrossRef]
- Tse, K.; Tse, H.; Sidney, J.; Sette, A.; Ley, K. T cells in atherosclerosis. Int. Immunol. 2013, 25, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Minty, A.; Chalon, P.; Derocq, J.-M.; Dumont, X.; Guillemot, J.-C.; Kaghad, M.; Labit, C.; Leplatois, P.; Liauzun, P.; Miloux, B.; et al. Interleukin-13 Is A New Human Lymphokine Regulating Inflammatory and Immune Responses. Nature 1993, 362, 248–250. [Google Scholar] [CrossRef] [PubMed]
- Vannier, E.; de Waal Malefyt, R.; Salazar-Montes, A.; de Vries, J.E.; Dinarello, C.A. Interleukin-13 (IL-13) induces IL-1 receptor antagonist gene expression and protein synthesis in peripheral blood mononuclear cells: Inhibition by an IL-4 mutant protein. Blood 1996, 87, 3307. [Google Scholar] [CrossRef] [Green Version]
- Bochner, B.S.; Klunk, D.A.; Sterbinsky, R.L.; Coffman, R.L.; Schleimer, R.P. IL-13 selectively induces vascular cell adhesion molecule-1 expression in human endothelial cells. J. Immunol. 1995, 154, 799. [Google Scholar]
- Fukushi, J.; Ono, M.; Morikawa, W.; Iwamoto, Y.; Kuwano, M. The Activity of Soluble VCAM-1 in Angiogenesis Stimulated by IL-4 and IL-13. J. Immunol. 2000, 165, 2818–2823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goebeler, M.; Schnarr, B.; Toksoy, A.; Kunz, M.; Borcker, E.B.; Duschl, A.; Gillitzer, R. Interleukin-13 selectively induces monocyte chemoattractant protein-1 synthesis and secretion by human endothelial cells: Involvement of IL-4Rα and Stat6 phosphorylation. Immunology 1997, 91, 450. [Google Scholar] [CrossRef] [PubMed]
- Halloran, M.M.; Haskell, C.J.; Woods, J.M.; Hosaka, S.; Koch, A.E.; Halloran, M. Interleukin-13 is an endothelial chemotaxin. Pathobiology 1997, 65, 287. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Kim, P.; Lee, W.; Hirani, A. Interleukin-4, Oxidative Stress, Vascular Inflammation and Atherosclerosis. Biomol. Ther. 2010, 18, 135–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, C.; Heimberger, A.; Gold, J.; O’Garra, A.; Murphy, K. Differential regulation of T helper phenotype development by interleukins 4 and 10 in an alpha beta T-cell-receptor transgenic system. Proc. Natl. Acad. Sci. USA 1992, 89, 6065–6069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallat, Z.; Heymes, C.; Ohan, J.; Faggin, E.; Lesèche, G.; Tedgui, A. Expression of Interleukin-10 in Advanced Human Atherosclerotic Plaques. Arterioscler. Thromb. Vasc. Biol. 1999, 19, 611–616. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Yang, W.; Lee, S.; Ahn, H.; Park, J.; Hwang, O.; Lee, J. TGF-β1 down-regulates inflammatory cytokine-induced VCAM-1 expression in cultured human glomerular endothelial cells. Nephrol. Dial. Transplant. 2000, 15, 596–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersson, J.; Libby, P.; Hansson, G. Adaptive immunity and atherosclerosis. Clin. Immunol. 2010, 134, 33–46. [Google Scholar] [CrossRef]
- Srikakulapu, P.; Hu, D.; Yin, C.; Mohanta, S.K.; Bontha, S.V.; Peng, L.; Beer, M.; Weber, C.; McNamara, C.A.; Grassia, G.; et al. Artery Tertiary Lymphoid Organs Control Multilayered Territorialized Atherosclerosis B-Cell Responses In Aged Apoe −/− Mice. Arterioscler. Thromb. Vasc. Biol. 2016, 36, 1174–1185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sämpi, M.; Ukkola, O.; Päivänsalo, M.; Kesäniemi, Y.A.; Binder, C.J.; Hörkkö, S. Plasma Interleukin-5 Levels Are Related To Antibodies Binding To Oxidized Low-Density Lipoprotein And To Decreased Subclinical Atherosclerosis. J. Am. Coll. Cardiol. 2008, 52, 1370–1378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rauch, P.J.; Chudnovskiy, A.; Robbins, C.S.; Weber, G.F.; Etzrodt, M.; Hilgendorf, I.; Tiglao, E.; Figueiredo, J.-L.; Iwamoto, Y.; Theurl, I.; et al. Innate Response Activator B Cells Protect Against Microbial Sepsis. Science 2012, 335, 597–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robbins, C.S.; Chudnovskiy, A.; Rauch, P.J.; Figueiredo, J.L.; Iwamoto, Y.; Gorbatov, R.; Etzrodt, M.; Weber, G.F.; Ueno, T.; van Rooijen, N.; et al. Extramedullary hematopoiesis generates Ly-6C(high) monocytes that infiltrate atherosclerotic lesions. Circulation 2012, 125, 364–374. [Google Scholar] [CrossRef] [Green Version]
- Frasca, D.; Diaz, A.; Romero, M.; Landin, A.; Blomberg, B. Age effects on B cells and humoral immunity in humans. Ageing Res. Rev. 2011, 10, 330–335. [Google Scholar] [CrossRef] [Green Version]
- Srikakulapu, P.; McNamara, C. B cells and atherosclerosis. Am. J. Physiol.-Heart Circ. Physiol. 2017, 312, H1060–H1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aiello, A.; Farzaneh, F.; Candore, G.; Caruso, C.; Davinelli, S.; Gambino, C.; Ligotti, M.; Zareian, N.; Accardi, G. Immunosenescence and Its Hallmarks: How to Oppose Aging Strategically? A Review of Potential Options for Therapeutic Intervention. Front. Immunol. 2019, 10, 2247. [Google Scholar] [CrossRef] [Green Version]
- Effros, R. Roy Walford and the immunologic theory of aging. Immun. Ageing 2005, 2, 7. [Google Scholar] [CrossRef] [Green Version]
- Fulop, T.; Larbi, A.; Dupuis, G.; Le Page, A.; Frost, E.; Cohen, A.; Witkowski, J.; Franceschi, C. Immunosenescence and Inflamm-Aging As Two Sides of the Same Coin: Friends or Foes? Front. Immunol. 2018, 8, 1960. [Google Scholar] [CrossRef] [Green Version]
- Franceschi, C.; Bonafè, M.; Valensin, S.; Olivieri, F.; De Luca, M.; Ottaviani, E.; De Benedictis, G. Inflamm-aging: An Evolutionary Perspective on Immunosenescence. Ann. N. Y. Acad. Sci. 2006, 908, 244–254. [Google Scholar] [CrossRef] [PubMed]
- Cohen, A. Complex systems dynamics in aging: New evidence, continuing questions. Biogerontology 2015, 17, 205–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franceschi, C.; Salvioli, S.; Garagnani, P.; de Eguileor, M.; Monti, D.; Capri, M. Immunobiography and the Heterogeneity of Immune Responses in the Elderly: A Focus on Inflammaging and Trained Immunity. Front. Immunol. 2017, 8, 982. [Google Scholar] [CrossRef] [Green Version]
- Davalli, P.; Mitic, T.; Caporali, A.; Lauriola, A.; D’Arca, D. ROS, Cell Senescence, and Novel Molecular Mechanisms in Aging and Age-Related Diseases. Oxidative Med. Cell. Longev. 2016, 2016, 3565127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- . Fulop, T.; Dupuis, G.; Baehl, S.; Le Page, A.; Bourgade, K.; Frost, E.; Witkowski, J.; Pawelec, G.; Larbi, A.; Cunnane, S. From inflamm-aging to immune-paralysis: A slippery slope during aging for immune-adaptation. Biogerontology 2015, 17, 147–157. [Google Scholar] [CrossRef]
- Barbé-Tuana, F.; Funchal, G.; Schmitz, C.; Maurmann, R.; Bauer, M. The interplay between immunosenescence and age-related diseases. Semin. Immunopathol. 2020, 42, 545–557. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, P.; Kotla, S.; Reddy Velatooru, L.; Abe, R.; Davis, E.; Cooke, J.; Schadler, K.; Deswal, A.; Herrmann, J.; Lin, S.; et al. Senescence-Associated Secretory Phenotype as a Hinge Between Cardiovascular Diseases and Cancer. Front. Cardiovasc. Med. 2021, 8, 763930. [Google Scholar] [CrossRef]
- Childs, B.; Li, H.; van Deursen, J. Senescent cells: A therapeutic target for cardiovascular disease. J. Clin. Investig. 2018, 128, 1217–1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stojanović, S.; Fiedler, J.; Bauersachs, J.; Thum, T.; Sedding, D. Senescence-induced inflammation: An important player and key therapeutic target in atherosclerosis. Eur. Heart J. 2020, 41, 2983–2996. [Google Scholar] [CrossRef] [Green Version]
- Fülöp, T.; Dupuis, G.; Witkowski, J.; Larbi, A. The Role of Immunosenescence in the Development of Age-Related Diseases. Rev. Investng. Clin. 2016, 68, 84–91. [Google Scholar]
- Yu, H.; Park, S.; Shin, E.; Lee, W. T cell senescence and cardiovascular diseases. Clin. Exp. Med. 2015, 6, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Linehan, E.; Fitzgerald, D. Ageing and the immune system: Focus on macrophages. Eur. J. Microbiol. Immunol. 2015, 5, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Michelsen, K.; Wong, M.; Shah, P.; Zhang, W.; Yano, J.; Doherty, T.; Akira, S.; Rajavashisth, T.; Arditi, M. Lack of Toll-like receptor 4 or myeloid differentiation factor 88 reduces atherosclerosis and alters plaque phenotype in mice deficient in apolipoprotein E. Proc. Natl. Acad. Sci. USA 2004, 101, 10679–10684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.; Ghosh, S. Toll-like receptor–mediated NF-κB activation: A phylogenetically conserved paradigm in innate immunity. J. Clin. Investig. 2001, 107, 13–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jazwinski, S.; Yashin, A. Aging and health—A systems biology perspective. Introduction. Interdiscip. Top. Gerontol. 2015, 40, VII–XII. [Google Scholar]
- Dodig, S.; Čepelak, I.; Pavić, I. Hallmarks of senescence and aging. Biochem. Med. 2019, 29, 483–497. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Espín, D.; Serrano, M. Cellular senescence: From physiology to pathology. Nat. Rev. Mol. Cell Biol. 2014, 15, 482–496. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yang, B.; Wang, J.; Cheng, F.; Shi, K.; Ying, L.; Wang, C.; Xia, K.; Huang, X.; Gong, Z.; et al. Cell Senescence: A Nonnegligible Cell State under Survival Stress in Pathology of Intervertebral Disc Degeneration. Oxidative Med. Cell. Longev. 2020, 2020, 9503562. [Google Scholar] [CrossRef]
- Herranz, N.; Gil, J. Mechanisms and functions of cellular senescence. J. Clin. Investig. 2018, 128, 1238–1246. [Google Scholar] [CrossRef] [Green Version]
- Kopp, H.; Hooper, A.; Shmelkov, S.; Rafii, S. ß-Galactosidase staining on bone marrow. The osteoclast pitfall. Histol. Histopathol. 2007, 22, 971–976. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.; Ryu, S.; Oh, Y.; Park, J.; Lee, J.; Kim, H.; Kim, K.; Jang, I.; Park, S. Morphological Adjustment of Senescent Cells by Modulating Caveolin-1 Status. J. Biol. Chem. 2004, 279, 42270–42278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, G.; Wordsworth, J.; Wang, C.; Jurk, D.; Lawless, C.; Martin-Ruiz, C.; von Zglinicki, T. A senescent cell bystander effect: Senescence-induced senescence. Aging Cell 2012, 11, 345–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hubackova, S.; Krejcikova, K.; Bartek, J.; Hodny, Z. IL1- and TGFβ-Nox4 signaling, oxidative stress and DNA damage response are shared features of replicative, oncogene-induced, and drug-induced paracrine ‘Bystander senescence’. Aging 2012, 4, 932–951. [Google Scholar] [CrossRef]
- Acosta, J.; Banito, A.; Wuestefeld, T.; Georgilis, A.; Janich, P.; Morton, J.; Athineos, D.; Kang, T.; Lasitschka, F.; Andrulis, M.; et al. A complex secretory program orchestrated by the inflammasome controls paracrine senescence. Nat. Cell Biol. 2013, 15, 978–990. [Google Scholar] [CrossRef]
- González-Gualda, E.; Baker, A.; Fruk, L.; Muñoz-Espín, D. A guide to assessing cellular senescence in vitro and in vivo. FEBS J. 2020, 288, 56–80. [Google Scholar] [CrossRef]
- Bojko, A.; Czarnecka-Herok, J.; Charzynska, A.; Dabrowski, M.; Sikora, E. Diversity of the Senescence Phenotype of Cancer Cells Treated with Chemotherapeutic Agents. Cells 2019, 8, 1501. [Google Scholar] [CrossRef] [Green Version]
- de Oliveira Mann, C.; Kranzusch, P. cGAS Conducts Micronuclei DNA Surveillance. Trends Cell Biol. 2017, 27, 697–698. [Google Scholar] [CrossRef]
- Rossi, M.; Abdelmohsen, K. The Emergence of Senescent Surface Biomarkers as Senotherapeutic Targets. Cells 2021, 10, 1740. [Google Scholar] [CrossRef]
- Lawson, C.; Wolf, S. ICAM-1 signaling in endothelial cells. Pharmacol. Rep. 2009, 61, 22–32. [Google Scholar] [CrossRef]
- Gorgoulis, V.; Pratsinis, H.; Zacharatos, P.; Demoliou, C.; Sigala, F.; Asimacopoulos, P.; Papavassiliou, A.; Kletsas, D. p53-Dependent ICAM-1 overexpression in senescent human cells identified in atherosclerotic lesions. Lab. Investig. 2005, 85, 502–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poston, R.; Haskard, D.; Coucher, J.; Gall, N.; Johnson-Tidey, R. Expression of intercellular adhesion molecule-1 in atherosclerotic plaques. Am. J. Pathol. 1992, 140, 665–673. [Google Scholar] [PubMed]
- Gorgoulis, V.G.; Zacharatos, P.; Kotsinas, A.; Kletsas, D.; Mariatos, G.; Zoumpourlis, V.; Ryan, K.M.; Kittas, C.; Papavassiliou, A.G. p53 activates ICAM-1 (CD54) expression in an NF-kappaB-independent manner. EMBO J. 2003, 22, 1567–1578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gross, M.; Bielinski, S.; Suarez-Lopez, J.; Reiner, A.; Bailey, K.; Thyagarajan, B.; Carr, J.; Duprez, D.; Jacobs, D. Circulating Soluble Intercellular Adhesion Molecule 1 and Subclinical Atherosclerosis: The Coronary Artery Risk Development in Young Adults Study. Clin. Chem. 2012, 58, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Park, Y. CD36, a scavenger receptor implicated in atherosclerosis. Exp. Mol. Med. 2014, 46, e99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, K.; Febbraio, M.; Li, W.; Silverstein, R. A Specific CD36-Dependent Signaling Pathway Is Required for Platelet Activation by Oxidized Low-Density Lipoprotein. Circ. Res. 2008, 102, 1512–1519. [Google Scholar] [CrossRef] [Green Version]
- Moore, K.; El Khoury, J.; Medeiros, L.; Terada, K.; Geula, C.; Luster, A.; Freeman, M. A CD36-initiated Signaling Cascade Mediates Inflammatory Effects of β-Amyloid. J. Biol. Chem. 2002, 277, 47373–47379. [Google Scholar] [CrossRef] [Green Version]
- Dawson, D.; Pearce, S.; Zhong, R.; Silverstein, R.; Frazier, W.; Bouck, N. CD36 Mediates the In Vitro Inhibitory Effects of Thrombospondin-1 on Endothelial Cells. J. Cell Biol. 1997, 138, 707–717. [Google Scholar] [CrossRef] [Green Version]
- Karuppagounder, V.; Giridharan, V.; Arumugam, S.; Sreedhar, R.; Palaniyandi, S.; Krishnamurthy, P.; Quevedo, J.; Watanabe, K.; Konishi, T.; Thandavarayan, R. Modulation of Macrophage Polarization and HMGB1-TLR2/TLR4 Cascade Plays a Crucial Role for Cardiac Remodeling in Senescence-Accelerated Prone Mice. PLoS ONE 2016, 11, e0152922. [Google Scholar] [CrossRef] [Green Version]
- Guy, E.; Kuchibhotla, S.; Silverstein, R.; Febbraio, M. Continued inhibition of atherosclerotic lesion development in long term Western diet fed CD36°/apoE° mice. Atherosclerosis 2007, 192, 123–130. [Google Scholar] [CrossRef]
- Yoon, I.; Kim, H.; Kim, Y.; Song, I.; Kim, W.; Kim, S.; Baek, S.; Kim, J.; Kim, J. Exploration of replicative senescence-associated genes in human dermal fibroblasts by cDNA microarray technology. Exp. Gerontol. 2004, 39, 1369–1378. [Google Scholar] [CrossRef]
- Amor, C.; Feucht, J.; Leibold, J.; Ho, Y.; Zhu, C.; Alonso-Curbelo, D.; Mansilla-Soto, J.; Boyer, J.; Li, X.; Giavridis, T.; et al. Senolytic CAR T cells reverse senescence-associated pathologies. Nature 2020, 583, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, N.; Mihalcioiu, C.; Rabbani, S. Multifaceted Role of the Urokinase-Type Plasminogen Activator (uPA) and Its Receptor (uPAR): Diagnostic, Prognostic, and Therapeutic Applications. Front. Oncol. 2018, 8, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ploplis, V.; Balsara, R.; Sandoval-Cooper, M.; Yin, Z.; Batten, J.; Modi, N.; Gadoua, D.; Donahue, D.; Martin, J.; Castellino, F. Enhanced in Vitro Proliferation of Aortic Endothelial Cells from Plasminogen Activator Inhibitor-1-deficient Mice. J. Biol. Chem. 2004, 279, 6143–6151. [Google Scholar] [CrossRef] [PubMed]
- Coppé, J.; Patil, C.; Rodier, F.; Sun, Y.; Muñoz, D.; Goldstein, J.; Nelson, P.; Desprez, P.; Campisi, J. Senescence-Associated Secretory Phenotypes Reveal Cell-Nonautonomous Functions of Oncogenic RAS and the p53 Tumor Suppressor. PLoS Biol. 2008, 6, e301. [Google Scholar] [CrossRef]
- Edsfeldt, A.; Nitulescu, M.; Grufman, H.; Grönberg, C.; Persson, A.; Nilsson, M.; Persson, M.; Björkbacka, H.; Gonçalves, I. Soluble Urokinase Plasminogen Activator Receptor is Associated with Inflammation in the Vulnerable Human Atherosclerotic Plaque. Stroke 2012, 43, 3305–3312. [Google Scholar] [CrossRef] [Green Version]
- Beaufort, N.; Leduc, D.; Rousselle, J.; Magdolen, V.; Luther, T.; Namane, A.; Chignard, M.; Pidard, D. Proteolytic Regulation of the Urokinase Receptor/CD87 on Monocytic Cells by Neutrophil Elastase and Cathepsin, G. J. Immunol. 2003, 172, 540–549. [Google Scholar] [CrossRef] [Green Version]
- Liang, S.; Li, Q.; Lai, Q.; Zhou, Y.; Zhang, H.; Chen, X.; Yao, B.; Xu, W.; Yang, X. Beta-2-Microglobulin is an Independent Risk Factor for Asymptomatic Carotid Atherosclerosis in Patients with Primary Aldosteronism. J. Atheroscler. Thromb. 2022, 29, 937–952. [Google Scholar] [CrossRef]
- Amighi, J.; Hoke, M.; Mlekusch, W.; Schlager, O.; Exner, M.; Haumer, M.; Pernicka, E.; Koppensteiner, R.; Minar, E.; Rumpold, H.; et al. Beta 2 Microglobulin and the Risk for Cardiovascular Events in Patients with Asymptomatic Carotid Atherosclerosis. Stroke 2011, 42, 1826–1833. [Google Scholar] [CrossRef] [Green Version]
- Qu, A.; Wu, X.; Li, S.; Sun, M.; Xu, L.; Kuang, H.; Xu, C. An NIR-Responsive DNA-Mediated Nanotetrahedron Enhances the Clearance of Senescent Cells. Adv. Mater. 2020, 32, 2000184. [Google Scholar] [CrossRef]
- Huang, J.; Liu, X.; Wei, Y.; Li, X.; Gao, S.; Dong, L.; Rao, X.; Zhong, J. Emerging Role of Dipeptidyl Peptidase-4 in Autoimmune Disease. Front. Immunol. 2022, 13, 830863. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Noh, J.; Bodogai, M.; Martindale, J.; Yang, X.; Indig, F.; Basu, S.; Ohnuma, K.; Morimoto, C.; Johnson, P.; et al. Identification of senescent cell surface targetable protein DPP4. Genes Dev. 2017, 31, 1529–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, Z.; Kampfrath, T.; Deiuliis, J.; Zhong, J.; Pineda, C.; Ying, Z.; Xu, X.; Lu, B.; Moffatt-Bruce, S.; Durairaj, R.; et al. Long-Term Dipeptidyl-Peptidase 4 Inhibition Reduces Atherosclerosis and Inflammation via Effects on Monocyte Recruitment and Chemotaxis. Circulation 2011, 124, 2338–2349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ta, N.; Schuyler, C.; Li, Y.; Lopes-Virella, M.; Huang, Y. DPP-4 (CD26) Inhibitor Alogliptin Inhibits Atherosclerosis in Diabetic Apolipoprotein E–Deficient Mice. J. Cardiovasc. Pharmacol. 2011, 58, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Ta, N.; Li, Y.; Schuyler, C.; Lopes-Virella, M.; Huang, Y. DPP-4 (CD26) inhibitor alogliptin inhibits TLR4-mediated ERK activation and ERK-dependent MMP-1 expression by U937 histiocytes. Atherosclerosis 2010, 213, 429–435. [Google Scholar] [CrossRef]
- Calvert, P.; Liew, T.; Gorenne, I.; Clarke, M.; Costopoulos, C.; Obaid, D.; O’Sullivan, M.; Shapiro, L.; McNab, D.; Densem, C.; et al. Leukocyte Telomere Length Is Associated With High-Risk Plaques on Virtual Histology Intravascular Ultrasound and Increased Proinflammatory Activity. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 2157–2164. [Google Scholar] [CrossRef] [Green Version]
- Martin, N.; Bernard, D. Calcium signaling and cellular senescence. Cell Calcium 2018, 70, 16–23. [Google Scholar] [CrossRef]
- Chong, Z.; Wang, S.; Shang, Y.; Maiese, K. Targeting cardiovascular disease with novel SIRT1 pathways. Future Cardiol. 2012, 8, 89–100. [Google Scholar] [CrossRef] [Green Version]
- Hodge, G.; Tran, H.; Reynolds, P.; Jersmann, H.; Hodge, S. Lymphocyte senescence in COPD is associated with decreased sirtuin 1 expression in steroid resistant pro-inflammatory lymphocytes. Ther. Adv. Respir. Dis. 2020, 14, 1753466620905280. [Google Scholar] [CrossRef]
- Jeng, M.; Hull, P.; Fei, M.; Kwon, H.; Tsou, C.; Kasler, H.; Ng, C.; Gordon, D.; Johnson, J.; Krogan, N.; et al. Metabolic reprogramming of human CD8+ memory T cells through loss of SIRT1. J. Exp. Med. 2017, 215, 51–62. [Google Scholar] [CrossRef] [Green Version]
- Moorhead, W.; Chu, C.; Cuevas, R.; Callahan, J.; Wong, R.; Regan, C.; Boufford, C.; Sur, S.; Liu, M.; Gomez, D.; et al. Dysregulation of FOXO1 (Forkhead Box O1 Protein) Drives Calcification in Arterial Calcification due to Deficiency of CD73 and Is Present in Peripheral Artery Disease. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 1680–1694. [Google Scholar] [CrossRef] [PubMed]
- Sardiello, M. Transcription factor EB: From master coordinator of lysosomal pathways to candidate therapeutic target in degenerative storage diseases. Ann. N. Y. Acad. Sci. 2016, 1, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Merino, A.; Buendia, P.; Martin-Malo, A.; Aljama, P.; Ramirez, R.; Carracedo, J. Senescent CD14+CD16+Monocytes Exhibit Proinflammatory and Proatherosclerotic Activity. J. Immunol. 2010, 186, 1809–1815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robbins, E.; Levine, E.; Eagle, H. Morphologic Changes Accompanying Senescence of Cultured Human Diploid Cells. J. Exp. Med. 1970, 131, 1211–1222. [Google Scholar] [CrossRef]
- Ng, K.; Gugiu, B.; Renganathan, K.; Davies, M.; Gu, X.; Crabb, J.; Kim, S.; Różanowska, M.; Bonilha, V.; Rayborn, M.; et al. Retinal Pigment Epithelium Lipofuscin Proteomics. Mol. Cell. Proteom. 2008, 7, 1397–1405. [Google Scholar] [CrossRef] [Green Version]
- Adler, L.; Boyer, N.; Chen, C.; Ablonczy, Z.; Crouch, R.; Koutalos, Y. The 11-cis Retinal Origins of Lipofuscin in the Retina. Prog. Mol. Biol. Transl. Sci. 2015, 134, e1–e12. [Google Scholar] [CrossRef]
- Terman, A.; Abrahamsson, N.; Brunk, U. Ceroid/Lipofuscin-loaded human fibroblasts show increased susceptibility to oxidative stress. Exp. Gerontol. 1999, 34, 755–770. [Google Scholar] [CrossRef]
- Brunk, U.; Terman, A. Lipofuscin: Mechanisms of age-related accumulation and influence on cell function. Free Radic. Biol. Med. 2002, 33, 611–619. [Google Scholar] [CrossRef]
- Kakimoto, Y.; Okada, C.; Kawabe, N.; Sasaki, A.; Tsukamoto, H.; Nagao, R.; Osawa, M. Myocardial lipofuscin accumulation in ageing and sudden cardiac death. Sci. Rep. 2019, 9, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Gerland, L.; Genestier, L.; Peyrol, S.; Michallet, M.; Hayette, S.; Urbanowicz, I.; Ffrench, P.; Magaud, J.; Ffrench, M. Autolysosomes accumulate during in vitro CD8+ T-lymphocyte aging and may participate in induced death sensitization of senescent cells. Exp. Gerontol. 2004, 39, 789–800. [Google Scholar] [CrossRef]
- Carroll, B.; Hewitt, G.; Korolchuk, V. Autophagy and ageing: Implications for age-related neurodegenerative diseases. Essays Biochem. 2013, 55, 119–131. [Google Scholar] [CrossRef]
- Ahmad, F.; Leake, D. Lysosomal oxidation of LDL alters lysosomal pH, induces senescence, and increases secretion of pro-inflammatory cytokines in human macrophages. J. Lipid Res. 2019, 60, 98–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marques, A.; Ramos, C.; Machado-Oliveira, G.; Vieira, O. Lysosome (Dys)function in Atherosclerosis—A Big Weight on the Shoulders of a Small Organelle. Front. Cell Dev. Biol. 2021, 9, 658995. [Google Scholar] [CrossRef] [PubMed]
- de Magalhães, J.; Passos, J. Stress, cell senescence and organismal ageing. Mech. Ageing Dev. 2018, 170, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Kurz, D.; Decary, S.; Hong, Y.; Erusalimsky, J. Senescence-associated (beta)-galactosidase reflects an increase in lysosomal mass during replicative ageing of human endothelial cells. J. Cell Sci. 2000, 113, 3613–3622. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Han, J.; Im, J.; Morrone, A.; Johung, K.; Goodwin, E.; Kleijer, W.; DiMaio, D.; Hwang, E. Senescence-associated β-galactosidase is lysosomal β-galactosidase. Aging Cell 2006, 5, 187–195. [Google Scholar] [CrossRef]
- Martínez-Zamudio, R.; Dewald, H.; Vasilopoulos, T.; Gittens-Williams, L.; Fitzgerald-Bocarsly, P.; Herbig, U. Senescence-associated β-galactosidase reveals the abundance of senescent CD8+ T cells in aging humans. Aging Cell 2021, 20, e13344. [Google Scholar] [CrossRef]
- de Mera-Rodríguez, J.; Álvarez-Hernán, G.; Gañán, Y.; Martín-Partido, G.; Rodríguez-León, J.; Francisco-Morcillo, J. Is Senescence-Associated β-Galactosidase a Reliable in vivo Marker of Cellular Senescence During Embryonic Development? Front. Cell Dev. Biol. 2021, 9, 623175. [Google Scholar] [CrossRef]
- Raffaele, M.; Kovacovicova, K.; Bonomini, F.; Rezzani, R.; Frohlich, J.; Vinciguerra, M. Senescence-like phenotype in post-mitotic cells of mice entering middle age. Aging 2020, 12, 13979–13990. [Google Scholar] [CrossRef]
- Gardner, S.; Humphry, M.; Bennett, M.; Clarke, M. Senescent Vascular Smooth Muscle Cells Drive Inflammation Through an Interleukin-1α–Dependent Senescence-Associated Secretory Phenotype. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 1963–1974. [Google Scholar] [CrossRef] [Green Version]
- Carosi, J.; Fourrier, C.; Bensalem, J.; Sargeant, T. The mTOR–lysosome axis at the centre of ageing. FEBS Open Bio 2021, 12, 739–757. [Google Scholar] [CrossRef] [PubMed]
- Sabbatinelli, J.; Prattichizzo, F.; Olivieri, F.; Procopio, A.; Rippo, M.; Giuliani, A. Where Metabolism Meets Senescence: Focus on Endothelial Cells. Front. Physiol. 2019, 10, 1523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weichhart, T. mTOR as Regulator of Lifespan, Aging, and Cellular Senescence: A Mini-Review. Gerontology 2017, 64, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Laberge, R.; Sun, Y.; Orjalo, A.; Patil, C.; Freund, A.; Zhou, L.; Curran, S.; Davalos, A.; Wilson-Edell, K.; Liu, S.; et al. MTOR regulates the pro-tumorigenic senescence-associated secretory phenotype by promoting IL1A translation. Nat. Cell Biol. 2015, 17, 1049–1061. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Lee, Y.; Cho, K.; Park, S. Adjustment of the lysosomal-mitochondrial axis for control of cellular senescence. Ageing Res. Rev. 2018, 47, 176–182. [Google Scholar] [CrossRef]
- Herranz, N.; Gallage, S.; Mellone, M.; Wuestefeld, T.; Klotz, S.; Hanley, C.; Raguz, S.; Acosta, J.; Innes, A.; Banito, A.; et al. mTOR regulates MAPKAPK2 translation to control the senescence-associated secretory phenotype. Nat. Cell Biol. 2015, 17, 1205–1217. [Google Scholar] [CrossRef] [Green Version]
- Cayo, A.; Segovia, R.; Venturini, W.; Moore-Carrasco, R.; Valenzuela, C.; Brown, N. mTOR Activity and Autophagy in Senescent Cells, a Complex Partnership. Int. J. Mol. Sci. 2021, 22, 8149. [Google Scholar] [CrossRef]
- Wong, P.; Puente, C.; Ganley, I.; Jiang, X. The ULK1 complex. Autophagy 2013, 9, 124–137. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Kundu, M.; Viollet, B.; Guan, K. AMPK and mTOR regulate autophagy through direct phosphorylation of Ulk1. Nat. Cell Biol. 2011, 13, 132–141. [Google Scholar] [CrossRef] [Green Version]
- Kiffin, R.; Bandyopadhyay, U.; Cuervo, A. Oxidative Stress and Autophagy. Antioxid. Redox Signal. 2006, 8, 152–162. [Google Scholar] [CrossRef]
- Jahrling, J.; Lin, A.; DeRosa, N.; Hussong, S.; Van Skike, C.; Girotti, M.; Javors, M.; Zhao, Q.; Maslin, L.; Asmis, R.; et al. mTOR drives cerebral blood flow and memory deficits in LDLR−/− mice modeling atherosclerosis and vascular cognitive impairment. J. Cereb. Blood Flow Metab. 2017, 38, 58–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Fan, Y.; Zhang, Z.; Fang, Y.; Cheng, X.; Yang, Q.; Liu, J.; Xie, J. mTOR in the Mechanisms of Atherosclerosis and Cardiovascular Disease. Discov. Med. 2021, 31, 129–140. [Google Scholar] [PubMed]
- Stallone, G.; Infante, B.; Prisciandaro, C.; Grandaliano, G. mTOR and Aging: An Old-Fashioned Dress. Int. J. Mol. Sci. 2019, 20, 2774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamano, T. Klotho upregulation by rapamycin protects against vascular disease in CKD. Kidney Int. 2015, 88, 660–662. [Google Scholar] [CrossRef]
- Omarjee, L.; Janin, A.; Perrot, F.; Laviolle, B.; Meilhac, O.; Mahe, G. Targeting T-cell senescence and cytokine storm with rapamycin to prevent severe progression in COVID-19. Clin. Immunol. 2020, 216, 108464. [Google Scholar] [CrossRef]
- Wang, R.; Yu, Z.; Sunchu, B.; Shoaf, J.; Dang, I.; Zhao, S.; Caples, K.; Bradley, L.; Beaver, L.; Ho, E.; et al. Rapamycin inhibits the secretory phenotype of senescent cells by a Nrf2-independent mechanism. Aging Cell 2017, 16, 564–574. [Google Scholar] [CrossRef]
- Childs, B.; Baker, D.; Wijshake, T.; Conover, C.; Campisi, J.; van Deursen, J. Senescent intimal foam cells are deleterious at all stages of atherosclerosis. Science 2016, 354, 472–477. [Google Scholar] [CrossRef] [Green Version]
- Hannou, S.; Wouters, K.; Paumelle, R.; Staels, B. Functional genomics of the CDKN2A/B locus in cardiovascular and metabolic disease: What have we learned from GWASs? Trends Endocrinol. Metab. 2015, 26, 176–184. [Google Scholar] [CrossRef]
- Holdt, L.; Sass, K.; Gäbel, G.; Bergert, H.; Thiery, J.; Teupser, D. Expression of Chr9p21 genes CDKN2B (p15INK4b), CDKN2A (p16INK4a, p14ARF) and MTAP in human atherosclerotic plaque. Atherosclerosis 2011, 214, 264–270. [Google Scholar] [CrossRef]
- Cardenas, J.; Owens, A.; Krishnamurthy, J.; Sharpless, N.; Whinna, H.; Church, F. Overexpressio144. n of the Cell Cycle Inhibitor p16 INK4a Promotes a Prothrombotic Phenotype Following Vascular Injury in Mice. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 827–833. [Google Scholar] [CrossRef] [Green Version]
- LaPak, K.; Burd, C. The Molecular Balancing Act of p16INK4a in Cancer and Aging. Mol. Cancer Res. 2014, 12, 167–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aw, N.; Canetti, E.; Suzuki, K.; Goh, J. Monocyte Subsets in Atherosclerosis and Modification with Exercise in Humans. Antioxidants 2018, 7, 196. [Google Scholar] [CrossRef] [Green Version]
- Grage-Griebenow, E.; Flad, H.; Ernst, M. Heterogeneity of human peripheral blood monocyte subsets. J. Leukoc. Biol. 2001, 69, 11–20. [Google Scholar] [PubMed]
- Ong, S.; Hadadi, E.; Dang, T.; Yeap, W.; Tan, C.; Ng, T.; Larbi, A.; Wong, S. The pro-inflammatory phenotype of the human non-classical monocyte subset is attributed to senescence. Cell Death Dis. 2018, 9, 266. [Google Scholar] [CrossRef]
- Wan, W.; Murphy, P. Regulation of Atherogenesis by Chemokines and Chemokine Receptors. Arch. Immunol. Ther. Exp. 2012, 61, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Bernhagen, J.; Krohn, R.; Lue, H.; Gregory, J.; Zernecke, A.; Koenen, R.; Dewor, M.; Georgiev, I.; Schober, A.; Leng, L.; et al. MIF is a noncognate ligand of CXC chemokine receptors in inflammatory and atherogenic cell recruitment. Nat. Med. 2007, 13, 587–596. [Google Scholar] [CrossRef]
- Weyand, C.; Brandes, J.; Schmidt, D.; Fulbright, J.; Goronzy, J. Functional properties of CD4+CD28− T cells in the aging immune system. Mech. Ageing Dev. 1998, 102, 131–147. [Google Scholar] [CrossRef]
- 1Jones, R.G.; Plas, D.R.; Kubek, S.; Buzzai, M.; Mu, J.; Xu, Y.; Birnbaum, M.J.; Thompson, C.B. AMP-activated protein kinase induces a p53-dependent metabolic checkpoint. Mol. Cell 2005, 18, 283–293. [Google Scholar] [CrossRef]
- Zawada, A.; Rogacev, K.; Rotter, B.; Winter, P.; Marell, R.; Fliser, D.; Heine, G. SuperSAGE evidence for CD14++CD16+ monocytes as a third monocyte subset. Blood 2011, 118, e50–e61. [Google Scholar] [CrossRef] [Green Version]
- Fan, H.; Hall, P.; Santos, L.; Gregory, J.; Fingerle-Rowson, G.; Bucala, R.; Morand, E.; Hickey, M. Macrophage Migration Inhibitory Factor and CD74 Regulate Macrophage Chemotactic Responses via MAPK and Rho GTPase. J. Immunol. 2011, 186, 4915–4924. [Google Scholar] [CrossRef] [Green Version]
- Machado-Oliveira, G.; Ramos, C.; Marques, A.; Vieira, O. Cell Senescence, Multiple Organelle Dysfunction and Atherosclerosis. Cells 2020, 9, 2146. [Google Scholar] [CrossRef]
- Watanabe, N.; Ikeda, U. Matrix metalloproteinases and atherosclerosis. Curr. Atheroscler. Rep. 2004, 6, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Banerjee, S.; Dey, N.; LeJeune, W.; Sarkar, P.; Brobey, R.; Rosenblatt, K.; Tilton, R.; Choudhary, S. Klotho Depletion Contributes to Increased Inflammation in Kidney of the db/db Mouse Model of Diabetes via RelA (Serine) 536 Phosphorylation. Diabetes 2011, 60, 1907–1916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aw, D.; Silva, A.; Palmer, D. Immunosenescence: Emerging challenges for an ageing population. Immunology 2007, 120, 435–446. [Google Scholar] [CrossRef] [PubMed]
- Plowden, J.; Renshaw-Hoelscher, M.; Engleman, C.; Katz, J.; Sambhara, S. Innate immunity in aging: Impact on macrophage function. Aging Cell 2004, 3, 161–167. [Google Scholar] [CrossRef]
- Fukuda, D.; Nishimoto, S.; Aini, K.; Tanaka, A.; Nishiguchi, T.; Kim-Kaneyama, J.; Lei, X.; Masuda, K.; Naruto, T.; Tanaka, K.; et al. Toll-Like Receptor 9 Plays A Pivotal Role In Angiotensin II–Induced Atherosclerosis. J. Am. Heart Assoc. 2019, 8, e010860. [Google Scholar] [CrossRef] [Green Version]
- Roberts, A.; Lee, B.; Deguine, J.; John, S.; Shlomchik, M.; Barton, G. Tissue-Resident Macrophages Are Locally Programmed for Silent Clearance of Apoptotic Cells. Immunity 2017, 47, 913–927. [Google Scholar] [CrossRef] [Green Version]
- Sene, A.; Khan, A.; Cox, D.; Nakamura, R.; Santeford, A.; Kim, B.; Sidhu, R.; Onken, M.; Harbour, J.; Hagbi-Levi, S.; et al. Impaired Cholesterol Efflux in Senescent Macrophages Promotes Age-Related Macular Degeneration. Cell Metab. 2013, 17, 549–561. [Google Scholar] [CrossRef] [Green Version]
- Willemsen, L.; Winther, M. Macrophage subsets in atherosclerosis as defined by single-cell technologies. J. Pathol. 2020, 250, 705–714. [Google Scholar] [CrossRef] [Green Version]
- Beckers, C.; Simpson, K.; Griffin, K.; Brown, J.; Cheah, L.; Smith, K.; Vacher, J.; Cordell, P.; Kearney, M.; Grant, P.; et al. Cre/lox Studies Identify Resident Macrophages as the Major Source of Circulating Coagulation Factor XIII-A. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 1494–1502. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Nishi, H.; Poles, J.; Niu, X.; Mccauley, C.; Rahman, K.; Brown, E.; Yeung, S.; Vozhilla, N.; Weinstock, A.; et al. Single-cell analysis of fate-mapped macrophages reveals heterogeneity, including stem-like properties, during atherosclerosis progression and regression. JCI Insight 2019, 4, e124574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, J.; Kim, E.; Son, Y.; Lee, D.; Park, Y.; Choi, J.; Cho, K.; Kwon, K.; Kim, J. CD9 induces cellular senescence and aggravates atherosclerotic plaque formation. Cell Death Differ. 2020, 27, 2681–2696. [Google Scholar] [CrossRef] [PubMed]
- Varasteh, Z.; Mohanta, S.; Li, Y.; López Armbruster, N.; Braeuer, M.; Nekolla, S.; Habenicht, A.; Sager, H.; Raes, G.; Weber, W.; et al. Targeting mannose receptor expression on macrophages in atherosclerotic plaques of apolipoprotein E-knockout mice using 68Ga-NOTA-anti-MMR nanobody: Non-invasive imaging of atherosclerotic plaques. EJNMMI Res. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Ouimet, M.; Ediriweera, H.; Afonso, M.; Ramkhelawon, B.; Singaravelu, R.; Liao, X.; Bandler, R.; Rahman, K.; Fisher, E.; Rayner, K.; et al. MicroRNA-33 Regulates Macrophage Autophagy in Atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 1058–1067. [Google Scholar] [CrossRef] [PubMed]
- Marquart, T.; Allen, R.; Ory, D.; Baldán, Á. miR-33 links SREBP-2 induction to repression of sterol transporters. Proc. Natl. Acad. Sci. USA 2010, 107, 12228–12232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Najafi-Shoushtari, S.; Kristo, F.; Li, Y.; Shioda, T.; Cohen, D.; Gerszten, R.; Näär, A. MicroRNA-33 and the SREBP Host Genes Cooperate to Control Cholesterol Homeostasis. Science 2010, 328, 1566–1569. [Google Scholar] [CrossRef] [Green Version]
- Rayner, K.; Suárez, Y.; Dávalos, A.; Parathath, S.; Fitzgerald, M.; Tamehiro, N.; Fisher, E.; Moore, K.; Fernández-Hernando, C. MiR-33 Contributes to the Regulation of Cholesterol Homeostasis. Science 2010, 328, 1570–1573. [Google Scholar] [CrossRef] [Green Version]
- Ouimet, M.; Koster, S.; Sakowski, E.; Ramkhelawon, B.; van Solingen, C.; Oldebeken, S.; Karunakaran, D.; Portal-Celhay, C.; Sheedy, F.; Ray, T.D.; et al. Mycobacterium tuberculosis induces the miR-33 locus to reprogram autophagy and host lipid metabolism. Nat. Immunol. 2016, 17, 677–686. [Google Scholar] [CrossRef] [Green Version]
- Hall, B.; Balan, V.; Gleiberman, A.; Strom, E.; Krasnov, P.; Virtuoso, L.; Rydkina, E.; Vujcic, S.; Balan, K.; Gitlin, I.; et al. Aging of mice is associated with p16(Ink4a)- and β-galactosidase-positive macrophage accumulation that can be induced in young mice by senescent cells. Aging 2016, 8, 1294–1315. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Li, J.; Chen, Y.; Zhang, S.; Feng, C.; Hou, Z.; Cai, J.; Wang, Y.; Hui, R.; Lv, B.; et al. MicroRNA-216a promotes M1 macrophages polarization and atherosclerosis progression by activating telomerase via the Smad3/NF-κB pathway. Biochim. Et Biophys. Acta (BBA)-Mol. Basis Dis. 2019, 1865, 1772–1781. [Google Scholar] [CrossRef]
- Korns, D.; Frasch, S.; Fernandez-Boyanapalli, R.; Henson, P.; Bratton, D. Modulation of Macrophage Efferocytosis in Inflammation. Front. Immunol. 2011, 2, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Maeyer, R.; Chambers, E.S. The impact of ageing on monocytes and macrophages. Immunol. Lett. 2021, 230, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, A.; Gupta, S. Impact of aging on dendritic cell functions in humans. Ageing Res. Rev. 2011, 10, 336–345. [Google Scholar] [CrossRef] [Green Version]
- Ruparelia, N.; Chai, J.T.; Fisher, E.A.; Choudhury, R.P. Inflammatory processes in cardiovascular disease: A route to targeted therapies. Nat. Rev. Cardiol. 2017, 14, 133–144. [Google Scholar] [CrossRef] [PubMed]
- American Heart Association. Inflammation and Heart Disease. 2015. Available online: https://www.heart.org/en/health-topics/consumer-healthcare/what-is-cardiovascular-disease/inflammation-and-heart-disease (accessed on 26 September 2022).
- Chougnet, C.; Thacker, R.; Shehata, H.; Hennies, C.; Lehn, M.; Lages, C.; Janssen, E. Loss of Phagocytic and Antigen Cross-Presenting Capacity in Aging Dendritic Cells Is Associated with Mitochondrial Dysfunction. J. Immunol. 2015, 195, 2624–2632. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.; Cheong, C.; Dandamudi, D.; Park, C.; Rodriguez, A.; Mehandru, S.; Velinzon, K.; Jung, I.; Yoo, J.; Oh, G.; et al. Flt3 Signaling-Dependent Dendritic Cells Protect against Atherosclerosis. Immunity 2011, 35, 819–831. [Google Scholar] [CrossRef] [Green Version]
- Castelo-Branco, C.; Soveral, I. The immune system and aging: A review. Gynecol. Endocrinol. 2014, 30, 16–22. [Google Scholar] [CrossRef]
- Agrawal, A.; Sridharan, A.; Prakash, S.; Agrawal, H. Dendritic cells and aging: Consequences for autoimmunity. Expert Rev. Clin. Immunol. 2012, 8, 73–80. [Google Scholar] [CrossRef] [Green Version]
- Lafuse, W.; Rajaram, M.; Wu, Q.; Moliva, J.; Torrelles, J.; Turner, J.; Schlesinger, L. Identification of an Increased Alveolar Macrophage Subpopulation in Old Mice That Displays Unique Inflammatory Characteristics and Is Permissive to Mycobacterium tuberculosis Infection. J. Immunol. 2019, 203, 2252–2264. [Google Scholar] [CrossRef]
- Zhang, X.; Niessner, A.; Nakajima, T.; Ma-Krupa, W.; Kopecky, S.; Frye, R.; Goronzy, J.; Weyand, C. Interleukin 12 Induces T-Cell Recruitment Into the Atherosclerotic Plaque. Circ. Res. 2006, 98, 524–531. [Google Scholar] [CrossRef]
- Gardner, J.K.; Mamotte, C.D.; McGonigle, T. Lipid-laden partially-activated plasmacytoid and CD4−CD8α+ dendritic cells accumulate in tissues in elderly mice. Immun. Ageing 2014, 11, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostan, R.; Monti, D.; Gueresi, P.; Bussolotto, M.; Franceschi, C.; Baggio, G. Gender, aging and longevity in humans: An update of an intriguing/neglected scenario paving the way to a gender-specific medicine. Clin. Sci. 2016, 130, 1711–1725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, J.; Brondstetter, T.; English, C.; Lee, H.; Virts, E.; Thoman, M. IL-7 Gene Therapy in Aging Restores Early Thymopoiesis without Reversing Involution. J. Immunol. 2004, 173, 4867–4874. [Google Scholar] [CrossRef] [Green Version]
- Ferrara, J.; Levine, J. Graft-Versus-Host Disease in the 21st Century: New Perspectives on an Old Problem. Semin. Hematol. 2006, 43, 1–2. [Google Scholar] [CrossRef]
- Reustle, A.; Torzewski, M. Role of p38 MAPK in Atherosclerosis and Aortic Valve Sclerosis. Int. J. Mol. Sci. 2018, 19, 3761. [Google Scholar] [CrossRef] [Green Version]
- Liuzzo, G.; Goronzy, J.; Yang, H.; Kopecky, S.; Holmes, D.; Frye, R.; Weyand, C. Monoclonal T-Cell Proliferation and Plaque Instability in Acute Coronary Syndromes. Circulation 2000, 101, 2883–2888. [Google Scholar] [CrossRef] [Green Version]
- Téo, F.; de Oliveira, R.; Mamoni, R.; Ferreira, M.; Nadruz, W.; Coelho, O.; Fernandes, J.; Blotta, M. Characterization of CD4+CD28null T cells in patients with coronary artery disease and individuals with risk factors for atherosclerosis. Cell. Immunol. 2013, 281, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Frasca, D. Senescent B cells in aging and age-related diseases: Their role in the regulation of antibody responses. Exp. Gerontol. 2018, 107, 55–58. [Google Scholar] [CrossRef] [PubMed]
- Forrest, A.; Kanamori-Katayama, M.; Tomaru, Y.; Lassmann, T.; Ninomiya, N.; Takahashi, Y.; de Hoon, M.; Kubosaki, A.; Kaiho, A.; Suzuki, M.; et al. Induction of microRNAs, mir-155, mir-222, mir-424 and mir-503, promotes monocytic differentiation through combinatorial regulation. Leukemia 2009, 24, 460–466. [Google Scholar] [CrossRef] [Green Version]
- Pirillo, A.; Norata, G.; Catapano, A. LOX-1, OxLDL, and Atherosclerosis. Mediat. Inflamm. 2013, 2013, 152786. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Thavarajah, T.; Gu, W.; Cai, J.; Xu, Q. Impact of miRNA in Atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2018, 38, e159–e170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feinberg, M.; Moore, K. MicroRNA Regulation of Atherosclerosis. Circ. Res. 2016, 118, 703–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.; Lin, L.; Cao, J.; Mao, X.; Qu, Y.; Xi, B. Up-regulated miR-93 contributes to coronary atherosclerosis pathogenesis through targeting ABCA1. Int. J. Clin. Exp. Med. 2015, 8, 674–681. [Google Scholar] [PubMed]
- Morton, A.; Rothman, A.; Greenwood, J.; Gunn, J.; Chase, A.; Clarke, B.; Hall, A.; Fox, K.; Foley, C.; Banya, W.; et al. The effect of interleukin-1 receptor antagonist therapy on markers of inflammation in non-ST elevation acute coronary syndromes: The MRC-ILA Heart Study. Eur. Heart J. 2014, 36, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Ridker, P.; MacFadyen, J.; Everett, B.; Libby, P.; Thuren, T.; Glynn, R. Relationship of C-reactive protein reduction to cardiovascular event reduction following treatment with canakinumab: A secondary analysis from the CANTOS randomised controlled trial. Lancet 2018, 391, 319–328. [Google Scholar] [CrossRef]
- Smith, H.; Marshall, C. Regulation of cell signalling by uPAR. Nat. Rev. Mol. Cell Biol. 2010, 11, 23–36. [Google Scholar] [CrossRef]
- Chimeric Antigen Receptor T Regulatory Cells for the Treatment of Atherosclerosis. U.S. Patent 62/663,776, 27 April 2018. Available online: https://patents.justia.com/patent/20210060071 (accessed on 6 June 2022).
- Navya, P.; Hema, K.; Munikumar, M.; Swargam, S.; Umamaheswari, A. Molecular docking of a beta-2-microglobulin drug target. Online J. Bioinform. 2012, 13, 222–231. [Google Scholar]
- Apostolakis, S.; Vogiatzi, K.; Amanatidou, V.; Spandidos, D. Interleukin 8 and cardiovascular disease. Cardiovasc. Res. 2009, 84, 353–360. [Google Scholar] [CrossRef] [Green Version]
- Hiebert, P.; Boivin, W.; Zhao, H.; McManus, B.; Granville, D. Perforin and Granzyme B Have Separate and Distinct Roles during Atherosclerotic Plaque Development in Apolipoprotein E Knockout Mice. PLoS ONE 2013, 8, e78939. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vellasamy, D.M.; Lee, S.-J.; Goh, K.W.; Goh, B.-H.; Tang, Y.-Q.; Ming, L.C.; Yap, W.H. Targeting Immune Senescence in Atherosclerosis. Int. J. Mol. Sci. 2022, 23, 13059. https://doi.org/10.3390/ijms232113059
Vellasamy DM, Lee S-J, Goh KW, Goh B-H, Tang Y-Q, Ming LC, Yap WH. Targeting Immune Senescence in Atherosclerosis. International Journal of Molecular Sciences. 2022; 23(21):13059. https://doi.org/10.3390/ijms232113059
Chicago/Turabian StyleVellasamy, Danusha Michelle, Sin-Jye Lee, Khang Wen Goh, Bey-Hing Goh, Yin-Quan Tang, Long Chiau Ming, and Wei Hsum Yap. 2022. "Targeting Immune Senescence in Atherosclerosis" International Journal of Molecular Sciences 23, no. 21: 13059. https://doi.org/10.3390/ijms232113059
APA StyleVellasamy, D. M., Lee, S. -J., Goh, K. W., Goh, B. -H., Tang, Y. -Q., Ming, L. C., & Yap, W. H. (2022). Targeting Immune Senescence in Atherosclerosis. International Journal of Molecular Sciences, 23(21), 13059. https://doi.org/10.3390/ijms232113059