
The Deadly Toxin Arsenal of the Tree-Dwelling Australian Funnel-Web Spiders

 ,
, 
 ,
,  , ,
, ,  , , and
, , and 
Abstract
:1. Introduction
2. Results
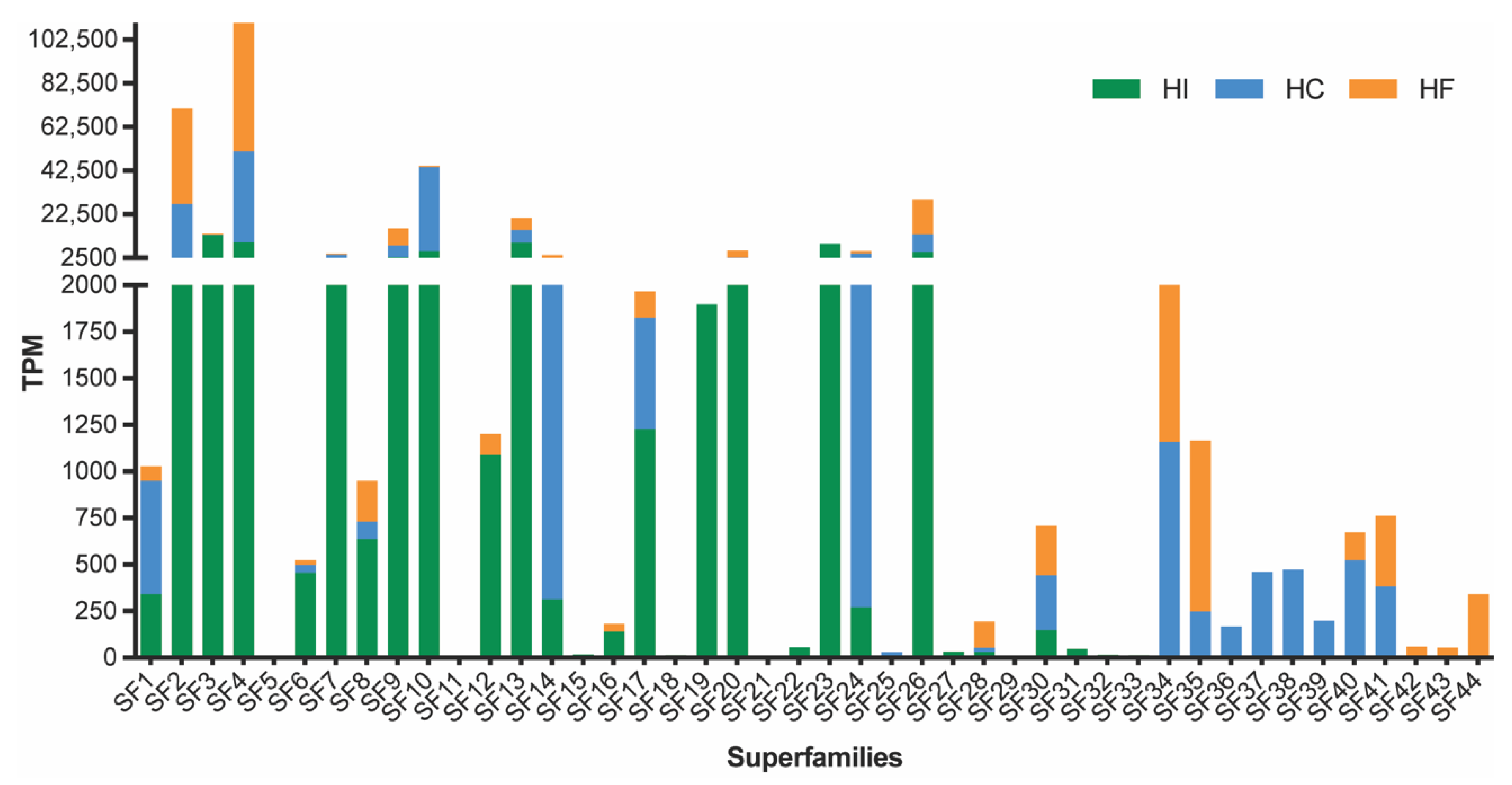
2.1. The Transcriptomic Diversity of H. cerberea and H. formidabilis Venom
2.2. Proteomics of H. formidabilis and H. cerberea Defensive Venoms
2.3. Molecular Evolution of Tree-Dweller Toxins
2.4. Insecticidal Bioassay
2.5. Bioactivity of FWS Venoms at Mammalian Ion Channels
3. Discussion
3.1. Composition and Evolution of Tree-Dwelling FWS Venoms
3.2. The Insecticidal Effects of Tree-Dwelling FWS Venoms
3.3. Explaining the Severity of the Tree-Dwelling FWS Envenomation
4. Materials and Methods
4.1. Venoms
4.2. H. cerberea and H. formidabilis Library Construction and Sequencing
4.3. Proteomics
4.4. Molecular Evolution Analysis
4.5. Blowfly Toxicity Assay
4.6. Mammalian Cell Culture
4.7. Calcium Influx Assays
4.8. Membrane Potential Assays (Nav)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Isbister, G.K.; Gray, M.R.; Balit, C.R.; Raven, R.J.; Stokes, B.J.; Porges, K.; Tankel, A.S.; Turner, E.; White, J.; Fisher, M.M. Funnel-web spider bite: A systematic review of recorded clinical cases. Med. J. Aust. 2005, 182, 407–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vetter, R.S.; Isbister, G.K. Medical aspects of spider bites. Annu. Rev. Entomol. 2008, 53, 409–429. [Google Scholar] [CrossRef] [PubMed]
- Hartman, L.J.; Sutherland, S.K. Funnel-web spider (Atrax robustus) antivenom in the treatment of human envenomation. Med. J. Aust. 1984, 141, 796–799. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.K.; Whyte, I.M.; White, J.; Keir, P.M. Clinical features and management of Hadronyche envenomation in man. Toxicon 2000, 38, 409–427. [Google Scholar] [CrossRef]
- Herzig, V.; Sunagar, K.; Wilson, D.T.R.; Pineda, S.S.; Israel, M.R.; Dutertre, S.; McFarland, B.S.; Undheim, E.A.B.; Hodgson, W.C.; Alewood, P.F.; et al. Australian funnel-web spiders evolved human-lethal δ-hexatoxins for defense against vertebrate predators. Proc. Natl. Acad. Sci. USA 2020, 117, 24920–24928. [Google Scholar] [CrossRef] [PubMed]
- Gray, M.R. A revision of the Australian funnel-web spiders (Hexathelidae: Atracinae). Rec. Aust. Mus. 2010, 62, 285–392. [Google Scholar] [CrossRef] [Green Version]
- Whitington, P.M.; Harris, K.L. A new species of Australian funnel-web spider (Mygalomorphae, Atracidae, Hadronyche) redefines the family Atracidae. Rec. Aust. Mus. 2021, 73, 115–122. [Google Scholar] [CrossRef]
- Pucca, M.B.; Knudsen, C.; Oliveira, I.S.; Rimbault, C.; Cerni, F.A.; Wen, F.H.; Sachett, J.; Sartim, M.A.; Laustsen, A.H.; Monteiro, W.M. Current knowledge on snake dry bites. Toxins 2020, 12, 668. [Google Scholar] [CrossRef] [PubMed]
- Chevreux, B.; Pfisterer, T.; Drescher, B.; Driesel, A.J.; Muller, W.E.G.; Wetter, T.; Suhai, S. Using the miraEST assembler for reliable and automated mRNA transcript assembly and SNP detection in sequenced ESTs. Genome Res. 2004, 14, 1147–1159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pineda, S.S.; Chin, Y.K.; Undheim, E.A.B.; Senff, S.; Mobli, M.; Dauly, C.; Escoubas, P.; Nicholson, G.M.; Kaas, Q.; Guo, S.; et al. Structural venomics reveals evolution of a complex venom by duplication and diversification of an ancient peptide-encoding gene. Proc. Natl. Acad. Sci. USA 2020, 117, 11399–11408. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Palagi, A.; Koh, J.M.S.; Leblanc, M.; Wilson, D.; Dutertre, S.; King, G.F.; Nicholson, G.M.; Escoubas, P. Unravelling the complex venom landscapes of Australian funnel-web spiders (Hexathelidae: Atracinae) using LC-MALDI-TOF mass spectrometry. J. Proteom. 2013, 80, 292–310. [Google Scholar] [CrossRef] [PubMed]
- Langenegger, N.; Nentwig, W.; Kuhn-Nentwig, L. Spider venom: Components, modes of action, and novel strategies in transcriptomic and proteomic analyses. Toxins 2019, 11, 611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graudins, A.; Wilson, D.; Alewood, P.F.; Broady, K.W.; Nicholson, G.M. Cross-reactivity of Sydney funnel-web spider antivenom: Neutralization of the in vitro toxicity of other Australian funnel-web (Atrax and Hadronyche) spider venoms. Toxicon 2002, 40, 259–266. [Google Scholar] [CrossRef]
- do Nascimento, S.M.; de Oliveira, U.C.; Nishiyama, M.Y.; Tashima, A.K.; da Silva, P.I. Presence of a neprilysin on Avicularia juruensis (Mygalomorphae: Theraphosidae) venom. Toxin Rev. 2022, 41, 370–379. [Google Scholar] [CrossRef]
- Chong, Y.; Hayes, J.L.; Sollod, B.; Wen, S.; Wilson, D.T.; Hains, P.G.; Hodgson, W.C.; Broady, K.W.; King, G.F.; Nicholson, G.M. The ω-atracotoxins: Selective blockers of insect M-LVA and HVA calcium channels. Biochem. Pharmacol. 2007, 74, 623–638. [Google Scholar] [CrossRef]
- Gunning, S.J.; Maggio, F.; Windley, M.J.; Valenzuela, S.M.; King, G.F.; Nicholson, G.M. The Janus-faced atracotoxins are specific blockers of invertebrate K(Ca) channels. FEBS J. 2008, 275, 4045–4059. [Google Scholar] [CrossRef] [Green Version]
- Windley, M.J.; Vetter, I.; Lewis, R.J.; Nicholson, G.M. Lethal effects of an insecticidal spider venom peptide involve positive allosteric modulation of insect nicotinic acetylcholine receptors. Neuropharmacology 2017, 127, 224–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, J.J.; Herzig, V.; King, G.F.; Alewood, P.F. The insecticidal potential of venom peptides. Cell. Mol. Life Sci. 2013, 70, 3665–3693. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Herzig, V.; King, G.F. Dipteran toxicity assays for determining the oral insecticidal activity of venoms and toxins. Toxicon 2018, 150, 297–303. [Google Scholar] [CrossRef]
- Cardoso, F.C.; Lewis, R.J. Sodium channels and pain: From toxins to therapies. Br. J. Pharmacol. 2018, 175, 2138–2157. [Google Scholar] [CrossRef] [Green Version]
- Uhlen, M.; Fagerberg, L.; Hallstrom, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, A.; Kampf, C.; Sjostedt, E.; Asplund, A.; et al. Proteomics. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Szeto, T.H.; Birinyi-Strachan, L.C.; Smith, R.; Connor, M.; Christie, M.J.; King, G.F.; Nicholson, G.M. Isolation and pharmacological characterisation of δ-atracotoxin-Hv1b, a vertebrate-selective sodium channel toxin. FEBS Lett. 2000, 470, 293–299. [Google Scholar] [CrossRef] [Green Version]
- Park, J.H.; Park, S.J.; Chung, M.K.; Jung, K.H.; Choi, M.R.; Kim, Y.; Chai, Y.G.; Kim, S.J.; Park, K.S. High expression of large-conductance Ca2+-activated K+ channel in the CD133+ subpopulation of SH-SY5Y neuroblastoma cells. Biochem. Biophys. Res. Commun. 2010, 396, 637–642. [Google Scholar] [CrossRef]
- Cardoso, F.C. Multi-targeting sodium and calcium channels using venom peptides for the treatment of complex ion channels-related diseases. Biochem. Pharmacol. 2020, 181, 114107. [Google Scholar] [CrossRef]
- Ren, Y.; Li, C.; Chang, J.; Wang, R.; Wang, Y.; Chu, X.P. Hi1a as a novel neuroprotective agent for ischemic stroke by inhibition of acid-sensing ion channel 1a. Transl. Stroke Res. 2018, 9, 96–98. [Google Scholar] [CrossRef]
- Chassagnon, I.R.; McCarthy, C.A.; Chin, Y.K.; Pineda, S.S.; Keramidas, A.; Mobli, M.; Pham, V.; De Silva, T.M.; Lynch, J.W.; Widdop, R.E.; et al. Potent neuroprotection after stroke afforded by a double-knot spider-venom peptide that inhibits acid-sensing ion channel 1a. Proc. Natl. Acad. Sci. USA 2017, 114, 3750–3755. [Google Scholar] [CrossRef] [Green Version]
- Pineda, S.S.; Chaumeil, P.A.; Kunert, A.; Kaas, Q.; Thang, M.W.C.; Li, L.; Nuhn, M.; Herzig, V.; Saez, N.J.; Cristofori-Armstrong, B.; et al. ArachnoServer 3.0: An online resource for automated discovery, analysis and annotation of spider toxins. Bioinformatics 2017, 34, 1074–1076. [Google Scholar] [CrossRef] [Green Version]
- King, G.F.; Gentz, M.C.; Escoubas, P.; Nicholson, G.M. A rational nomenclature for naming peptide toxins from spiders and other venomous animals. Toxicon 2008, 52, 264–276. [Google Scholar] [CrossRef] [Green Version]
- Giribaldi, J.; Kazandjian, T.; Amorim, F.G.; Whiteley, G.; Wagstaff, S.C.; Cazals, G.; Enjalbal, C.; Quinton, L.; Casewell, N.R.; Dutertre, S. Venomics of the asp viper Vipera aspis aspis from France. J. Proteom. 2020, 218, 103707. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C.; Batzoglou, S. Multiple sequence alignment. Curr. Opin. Struct. Biol. 2006, 16, 368–373. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z. PAML 4: Phylogenetic analysis by maximum likelihood. Mol. Biol. Evol. 2007, 24, 1586–1591. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Wong, W.S.; Nielsen, R. Bayes empirical bayes inference of amino acid sites under positive selection. Mol. Biol. Evol. 2005, 22, 1107–1118. [Google Scholar] [CrossRef] [Green Version]
- Murrell, B.; Wertheim, J.O.; Moola, S.; Weighill, T.; Scheffler, K.; Kosakovsky Pond, S.L. Detecting individual sites subject to episodic diversifying selection. PLoS Genet. 2012, 8, e1002764. [Google Scholar] [CrossRef] [Green Version]
- Maldonado, E.; Sunagar, K.; Almeida, D.; Vasconcelos, V.; Antunes, A. IMPACT_S: Integrated multiprogram platform to analyze and combine tests of selection. PLoS ONE 2014, 9, e96243. [Google Scholar] [CrossRef] [Green Version]
- Herzig, V.; Hodgson, W.C. Neurotoxic and insecticidal properties of venom from the Australian theraphosid spider Selenotholus foelschei. Neurotoxicology 2008, 29, 471–475. [Google Scholar] [CrossRef]
- Cardoso, F.C.; Dekan, Z.; Rosengren, K.J.; Erickson, A.; Vetter, I.; Deuis, J.; Herzig, V.; Alewood, P.; King, G.F.; Lewis, R.J. Identification and characterization of ProTx-III [μ-TRTX-Tp1a], a new voltage-gated sodium channel inhibitor from venom of the tarantula Thrixopelma pruriens. Mol. Pharmacol. 2015, 88, 291–303. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, F.C.; Castro, J.; Grundy, L.; Schober, G.; Garcia-Caraballo, S.; Zhao, T.; Herzig, V.; King, G.F.; Brierley, S.M.; Lewis, R.J. A spider-venom peptide with multitarget activity on sodium and calcium channels alleviates chronic visceral pain in a model of irritable bowel syndrome. Pain 2021, 162, 569–581. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PAML (M8) | TreeSAAP | MEME Sites | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Toxin Superfamily | PS | ω | Conservative Changes in A.A Properties | Radical Changes in A.A Properties | |||||
| Chemical | Physical | Total | Chemical | Physical | Total | ||||
| Hadronyche formidablis: True Tree-Dweller | |||||||||
| SF13 | - | 0.64 | - | - | - | - | - | - | 1 |
| SF9 | - | 0.62 | - | - | - | - | - | - | 19 |
| SF10 | 3 | 2.59 | 2 | - | 2 | - | 3 | 3 | 0 |
| SF4 | 10 | 0.98 | - | 16 | 16 | 51 | 51 | 45 | |
| SF26 | 21 | 1.55 | - | 14 | 14 | - | 53 | 53 | 6 |
| Hadronyche cerberea: Newly Adapted Tree-Dweller | |||||||||
| SF13 | - | 0.20 | - | - | - | - | - | - | 0 |
| SF9 | 2 | 1.90 | - | - | - | - | - | - | 1 |
| SF10 | - | 0.75 | - | - | - | - | - | - | 1 |
| SF4 | 23 | 2.85 | - | 19 | 19 | - | 72 | 72 | 18 |
| SF26 | 24 | 1.85 | 12 | 6 | 18 | - | 81 | 81 | 18 |
| Hydronyche infensa: Ground-Dweller from Atracidae Family | |||||||||
| SF13 | 17 | 1.58 | - | 32 | 32 | - | 7 | 7 | 8 |
| SF9 | - | 0.64 | - | - | - | - | - | - | 15 |
| SF10 | 11 | 0.94 | - | 14 | 14 | 9 | - | 9 | 8 |
| SF4 | 30 | 1.42 | - | 59 | 79 | - | 327 | 327 | 73 |
| SF26 | 23 | 1.18 | - | 10 | 10 | - | 119 | 119 | 14 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cardoso, F.C.; Pineda, S.S.; Herzig, V.; Sunagar, K.; Shaikh, N.Y.; Jin, A.-H.; King, G.F.; Alewood, P.F.; Lewis, R.J.; Dutertre, S. The Deadly Toxin Arsenal of the Tree-Dwelling Australian Funnel-Web Spiders. Int. J. Mol. Sci. 2022, 23, 13077. https://doi.org/10.3390/ijms232113077
Cardoso FC, Pineda SS, Herzig V, Sunagar K, Shaikh NY, Jin A-H, King GF, Alewood PF, Lewis RJ, Dutertre S. The Deadly Toxin Arsenal of the Tree-Dwelling Australian Funnel-Web Spiders. International Journal of Molecular Sciences. 2022; 23(21):13077. https://doi.org/10.3390/ijms232113077
Chicago/Turabian StyleCardoso, Fernanda C., Sandy S. Pineda, Volker Herzig, Kartik Sunagar, Naeem Yusuf Shaikh, Ai-Hua Jin, Glenn F. King, Paul F. Alewood, Richard J. Lewis, and Sébastien Dutertre. 2022. "The Deadly Toxin Arsenal of the Tree-Dwelling Australian Funnel-Web Spiders" International Journal of Molecular Sciences 23, no. 21: 13077. https://doi.org/10.3390/ijms232113077
APA StyleCardoso, F. C., Pineda, S. S., Herzig, V., Sunagar, K., Shaikh, N. Y., Jin, A. -H., King, G. F., Alewood, P. F., Lewis, R. J., & Dutertre, S. (2022). The Deadly Toxin Arsenal of the Tree-Dwelling Australian Funnel-Web Spiders. International Journal of Molecular Sciences, 23(21), 13077. https://doi.org/10.3390/ijms232113077