

Quercetin and Isorhamnetin Reduce Benzo[a]pyrene-Induced Genotoxicity by Inducing RAD51 Expression through Downregulation of miR−34a

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
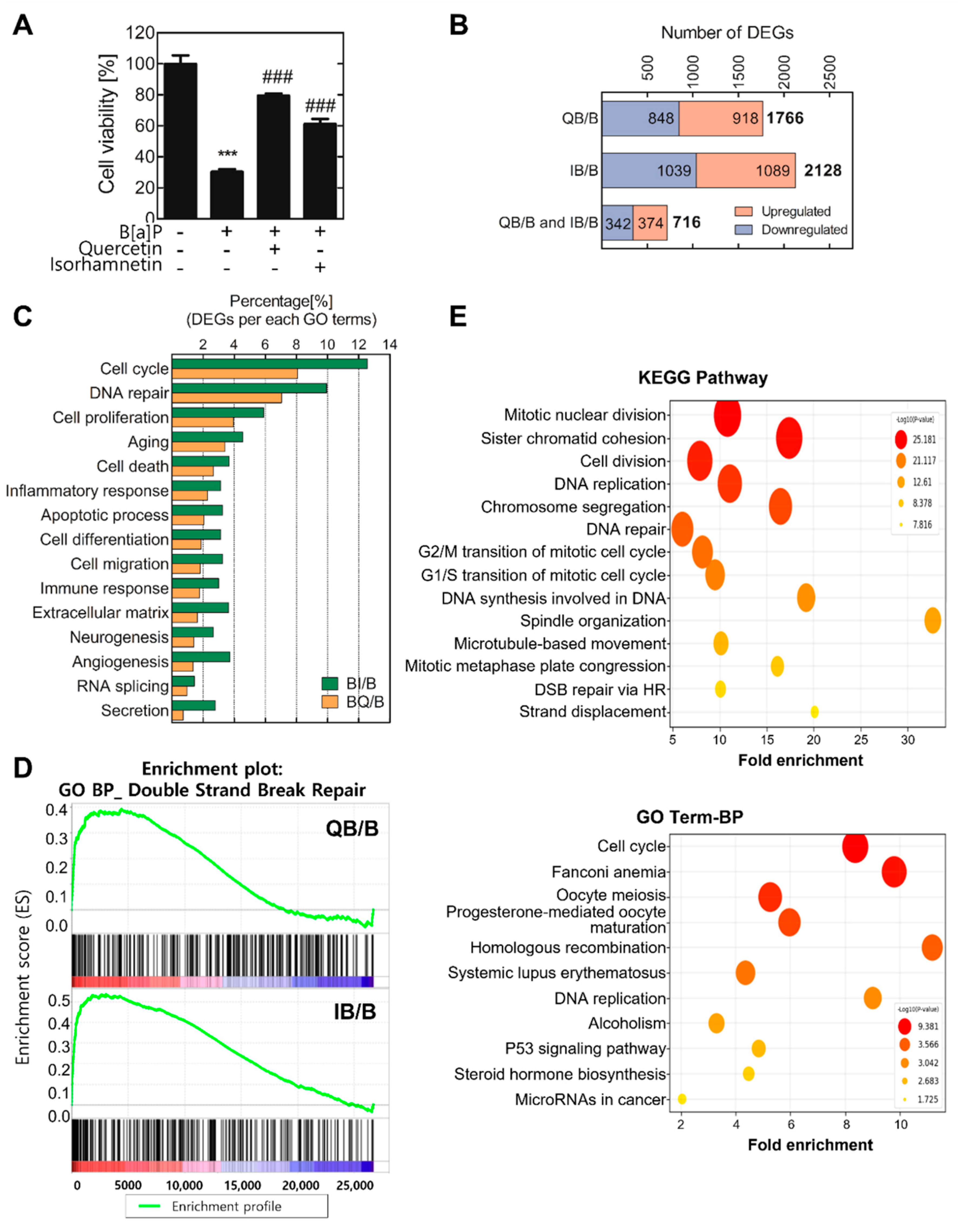
2.1. Quercetin and Isorhamnetin Each Upregulate Homologous Recombination through RAD51 Expression
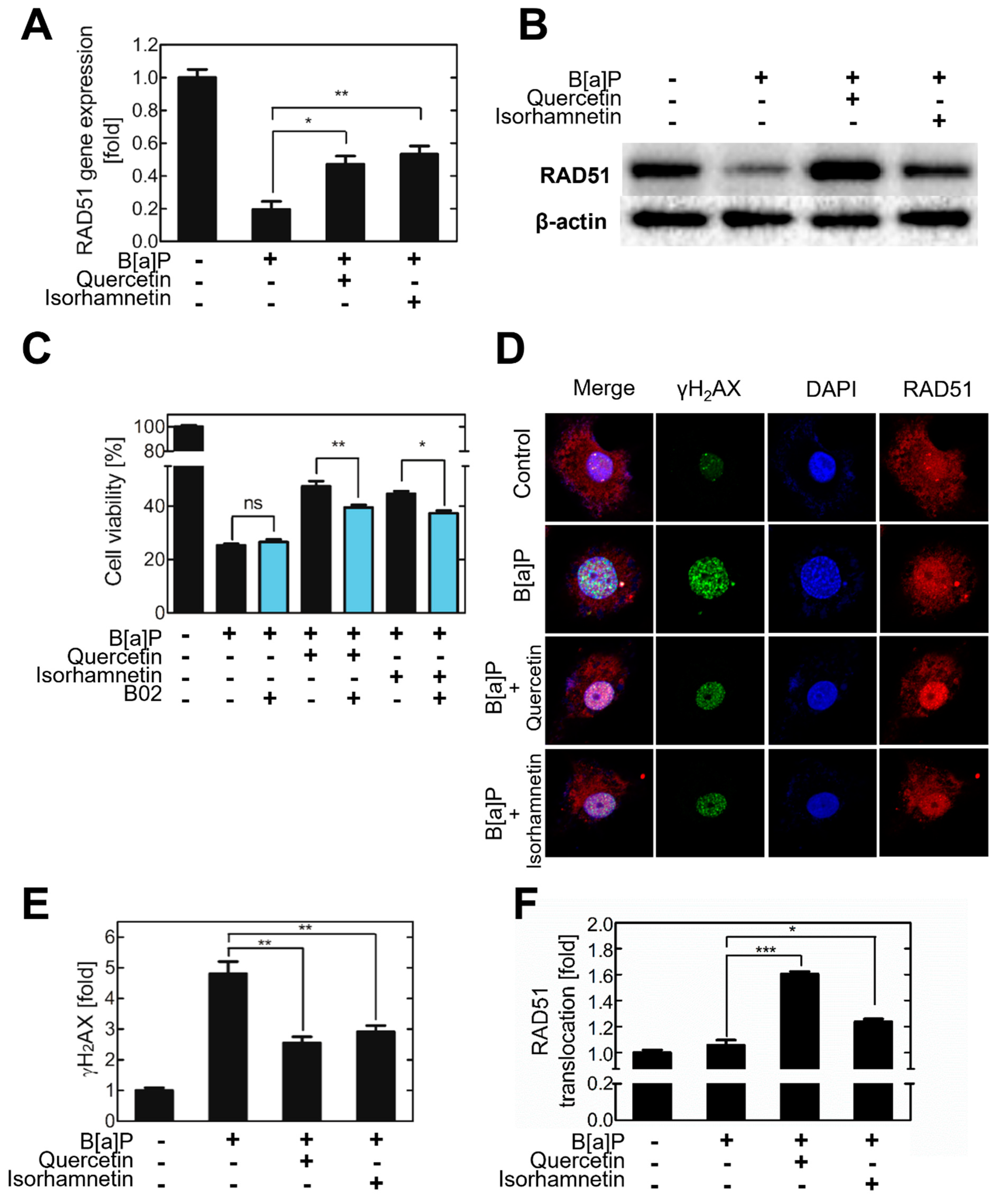
2.2. Quercetin and Isorhamnetin Reduce DNA DSBs and Increase RAD51 Expression
2.3. Quercetin and Isorhamnetin Decrease Cellular miR−34a Levels Which Increased by B[a]P
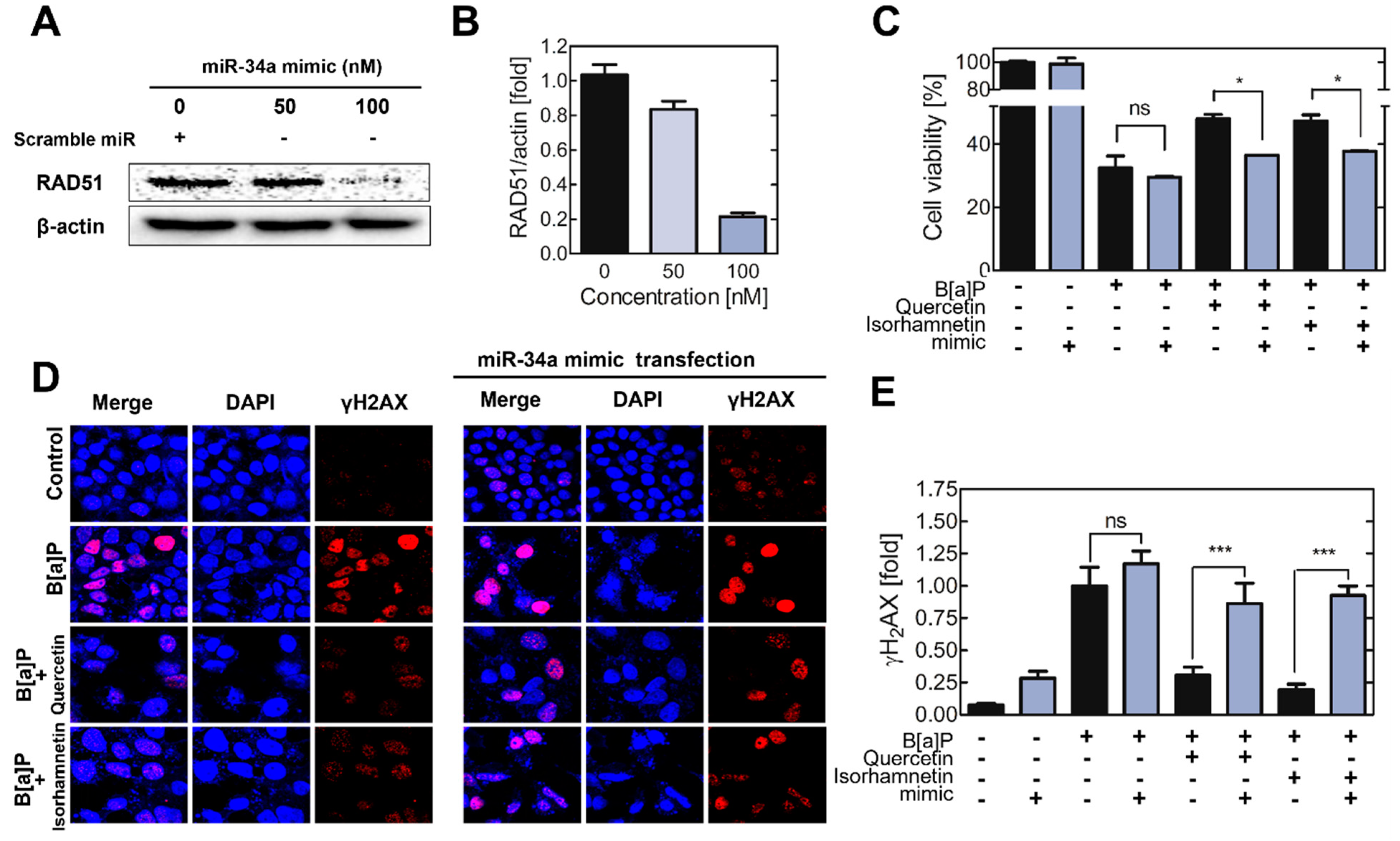
2.4. Transfection of the Cells with a miR−34a Mimic Mitigate the Effects of Quercetin and Isorhamnetin on DSBs and RAD51 Expression
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Culture and Treatment
4.3. Cell-Viability Analysis
4.4. mRNA Extraction
4.5. Microarray Analysis
4.6. Gene Ontology and Pathway Analysis
4.7. Protein–Protein Interaction (PPI) Mapping
4.8. Gene Set Enrichment Analysis (GSEA)
4.9. RT-qPCR
4.10. Immunofluorescence Analysis
4.11. Western-Blot Analysis
4.12. Transfection
4.13. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Baan, R.; Grosse, Y.; Straif, K.; Secretan, B.; El Ghissassi, F.; Bouvard, V.; Benbrahim−tallaa, L.; Guha, N.; Freeman, C.; Galichet, L.; et al. A review of human carcinogens—Part F: Chemical agents and related occupations. Lancet Oncol. 2009, 10, 1143–1144. [Google Scholar] [CrossRef]
- Burchiel, S.W.; Thompson, T.A.; Lauer, F.T.; Oprea, T.I. Activation of dioxin response element (DRE)-associated genes by benzo(a)pyrene 3,6-quinone and benzo(a)pyrene 1,6-quinone in MCF-10A human mammary epithelial cells. Toxicol. Appl. Pharmacol. 2007, 221, 203–214. [Google Scholar] [CrossRef] [Green Version]
- Straif, K.; Baan, R.; Grosse, Y.; Secretan, B.; El Ghissassi, F.; Cogliano, V.; WHO International Agency for Research on Cancer Monograph Working Group. Carcinogenicity of polycyclic aromatic hydrocarbons. Lancet Oncol. 2005, 6, 931–932. [Google Scholar] [CrossRef]
- Huberman, E.; Sachs, L.; Yang, S.K.; Gelboin, V. Identification of mutagenic metabolites of benzo(a)pyrene in mammalian cells. Proc. Natl. Acad. Sci. USA 1976, 73, 607–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papamichos-Chronakis, M.; Peterson, C.L. Chromatin and the genome integrity network. Nat. Rev. Genet. 2013, 14, 62–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cannan, W.J.; Pederson, D.S. Mechanisms and Consequences of Double-Strand DNA Break Formation in Chromatin. J. Cell. Physiol. 2016, 231, 3–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kushman, M.E.; Kabler, S.L.; Fleming, M.H.; Ravoori, S.; Gupta, R.C.; Doehmer, J.; Morrow, C.S.; Townsend, A.J. Expression of human glutathione S−transferase P1 confers resistance to benzo[a]pyrene or benzo[a]pyrene-7,8-dihydrodiol mutagenesis, macromolecular alkylation and formation of stable N2-Gua-BPDE adducts in stably transfected V79MZ cells co-expressing hCYP1A1. Carcinogenesis 2007, 28, 207–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterjee, N.; Walker, G.C. Mechanisms of DNA damage, repair, and mutagenesis. Environ. Mol. Mutagen. 2017, 58, 235–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christmann, M.; Kaina, B. Transcriptional regulation of human DNA repair genes following genotoxic stress: Trigger mechanisms, inducible responses and genotoxic adaptation. Nucleic Acids Res. 2013, 41, 8403–8420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lieber, M.R. The mechanism of double-strand DNA break repair by the nonhomologous DNA end-joining pathway. Annu. Rev. Biochem. 2010, 79, 181–211. [Google Scholar] [CrossRef] [PubMed]
- Mao, Z.; Bozzella, M.; Seluanov, A.; Gorbunova, V. DNA repair by nonhomologous end joining and homologous recombination during cell cycle in human cells. Cell Cycle 2008, 7, 2902–2906. [Google Scholar] [CrossRef] [PubMed]
- Wright, W.D.; Shah, S.S.; Heyer, W.D. Homologous recombination and the repair of DNA double-strand breaks. J. Biol. Chem. 2018, 293, 10524–10535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krejci, L.; Altmannova, V.; Spirek, M.; Zhao, X. Homologous recombination and its regulation. Nucleic Acids Res. 2012, 40, 5795–5818. [Google Scholar] [CrossRef] [PubMed]
- Tavares, E.M.; Wright, W.D.; Heyer, W.D.; Le Cam, E.; Dupaigne, P. In vitro role of Rad54 in Rad51-ssDNA filament-dependent homology search and synaptic complexes formation. Nat. Commun. 2019, 10, 4058. [Google Scholar] [CrossRef] [Green Version]
- Rupaimoole, R.; Slack, F.J. MicroRNA therapeutics: Towards a new era for the management of cancer and other diseases. Nat. Rev. Drug Discov. 2017, 16, 203–222. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [Green Version]
- Friedman, R.C.; Farh, K.K.; Burge, C.B.; Bartel, D.P. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef] [Green Version]
- Felekkis, K.; Touvana, E.; Stefanou, C.; Deltas, C. microRNAs: A newly described class of encoded molecules that play a role in health and disease. Hippokratia 2010, 14, 236–240. [Google Scholar]
- Tanaka, S.; Trakooncharoenvit, A.; Nishikawa, M.; Ikushiro, S.; Hara, H. Comprehensive Analyses of Quercetin Conjugates by LC/MS/MS Revealed That Isorhamnetin-7-O-glucuronide-4′-O-sulfate Is a Major Metabolite in Plasma of Rats Fed with Quercetin Glucosides. J. Agric. Food Chem. 2019, 67, 4240–4249. [Google Scholar] [CrossRef]
- Kim, M.; Jee, S.C.; Kim, K.S.; Kim, H.S.; Yu, K.N.; Sung, J.S. Quercetin and Isorhamnetin Attenuate Benzo[a]pyrene-Induced Toxicity by Modulating Detoxification Enzymes through the AhR and NRF2 Signaling Pathways. Antioxidants 2021, 10, 787. [Google Scholar] [CrossRef]
- Zhu, B.T.; Ezell, E.L.; Liehr, J.G. Catechol-O-Methyltransferase-Catalyzed Rapid O-Methylation of Mutagenic Flavonoids - Metabolic Inactivation as a Possible Reason for Their Lack of Carcinogenicity in-Vivo. J. Biol. Chem. 1994, 269, 292–299. [Google Scholar] [CrossRef]
- Liu, Y.; Wu, Y.M.; Zhang, P.Y. Protective effects of curcumin and quercetin during benzo(a)pyrene induced lung carcinogenesis in mice. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 1736–1743. [Google Scholar] [PubMed]
- Hashemzaei, M.; Far, A.D.; Yari, A.; Heravi, R.E.; Tabrizian, K.; Taghdisi, S.M.; Sadegh, S.E.; Tsarouhas, K.; Kouretas, D.; Tzanakakis, G.; et al. Anticancer and apoptosis-inducing effects of quercetin in vitro and in vivo. Oncol. Rep. 2017, 38, 819–828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.; Jee, S.C.; Kim, S.; Hwang, K.H.; Sung, J.S. Identification and Characterization of mRNA Biomarkers for Sodium Cyanide Exposure. Toxics 2021, 9, 288. [Google Scholar] [CrossRef]
- Aygun, S.F.; Kabadayi, F. Determination of benzo[a]pyrene in charcoal grilled meat samples by HPLC with fluorescence detection. Int. J. Food Sci. Nutr. 2005, 56, 581–585. [Google Scholar] [CrossRef]
- Alhmoud, J.F.; Woolley, J.F.; Al Moustafa, A.E.; Malki, M.I. DNA Damage/Repair Management in Cancers. Cancers 2020, 12, 1050. [Google Scholar] [CrossRef]
- Hess, M.T.; Gunz, D.; Luneva, N.; Geacintov, N.E.; Naegeli, H. Base pair conformation-dependent excision of benzo[a]pyrene diol epoxide-guanine adducts by human nucleotide excision repair enzymes. Mol. Cell. Biol. 1997, 17, 7069–7076. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Zhou, Y.; Wang, Z.; Hao, Q.; Ren, S. [Effect of benzo[a]pyrene on DNA damage and expression of genes involved in nucleotide excision repair in lung cancer cells]. Zhonghua Lao Dong Wei Sheng Zhi Ye Bing Za Zhi = Zhonghua Laodong Weisheng Zhiyebing Zazhi = Chin. J. Ind. Hyg. Occup. Dis. 2002, 20, 443–445. [Google Scholar]
- Kress, J.M.; Dio, L.D.; Heck, L.; Pulliero, A.; Izzotti, A.; Laarmann, K.; Fritz, G.; Kaina, B. Human primary endothelial cells are impaired in nucleotide excision repair and sensitive to benzo[a]pyrene compared with smooth muscle cells and pericytes. Sci. Rep. 2019, 9, 13800. [Google Scholar] [CrossRef] [Green Version]
- Misaki, K.; Matsui, S.; Matsuda, T. Metabolic enzyme induction by HepG2 cells exposed to oxygenated and nonoxygenated polycyclic aromatic hydrocarbons. Chem. Res. Toxicol. 2007, 20, 277–283. [Google Scholar] [CrossRef]
- Manach, C.; Texier, O.; Regerat, F.; Agullo, G.; Demigne, C.; Remesy, C. Dietary quercetin is recovered in rat plasma as conjugated derivatives of isorhamnetin and quercetin. J. Nutr. Biochem. 1996, 7, 375–380. [Google Scholar] [CrossRef]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Chen, Q.; Dahan, A.; Xue, J.; Wei, L.; Tan, W.; Zhang, G. Transcriptomic analyses reveal the molecular mechanisms of schisandrin B alleviates CCl4-induced liver fibrosis in rats by RNA-sequencing. Chem. -Biol. Interact. 2019, 309, 108675. [Google Scholar] [CrossRef] [PubMed]
- Campos, A.; Clemente-Blanco, A. Cell Cycle and DNA Repair Regulation in the Damage Response: Protein Phosphatases Take Over the Reins. Int. J. Mol. Sci. 2020, 21, 446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, Y.; Huang, T.; Yu, Q.; Liu, B.; Wang, J.; Wang, Z.; Huang, X. Sorafenib suppresses proliferation rate of fibroblast-like synoviocytes through the arrest of cell cycle in experimental adjuvant arthritis. J. Pharm. Pharmacol. 2021, 73, 32–39. [Google Scholar] [CrossRef]
- Brandsma, I.; Gent, D.C. Pathway choice in DNA double strand break repair: Observations of a balancing act. Genome Integr. 2012, 3, 9. [Google Scholar] [CrossRef] [Green Version]
- Featherstone, C.; Jackson, S.P. DNA double-strand break repair. Curr. Biol. 1999, 9, R759–R761. [Google Scholar] [CrossRef] [Green Version]
- Demeyer, A.; Benhelli-Mokrani, H.; Chenais, B.; Weigel, P.; Fleury, F. Inhibiting homologous recombination by targeting RAD51 protein. Biochim. Biophys. Acta Rev. Cancer 2021, 1876, 188597. [Google Scholar] [CrossRef]
- Mah, L.J.; El-Osta, A.; Karagiannis, T.C. gammaH2AX: A sensitive molecular marker of DNA damage and repair. Leukemia 2010, 24, 679–686. [Google Scholar] [CrossRef] [Green Version]
- Gildemeister, O.S.; Sage, J.M.; Knight, K.L. Cellular redistribution of Rad51 in response to DNA damage: Novel role for Rad51C. J. Biol. Chem. 2009, 284, 31945–31952. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.M.; Taniguchi, T. MicroRNAs and DNA damage response Implications for cancer therapy. Cell Cycle 2013, 12, 32–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.E.; Pan, Y.; Park, E.; Konstantinopoulos, P.; De, S.; D’Andrea, A.; Chowdhury, D. MicroRNAs down-regulate homologous recombination in the G1 phase of cycling cells to maintain genomic stability. eLife 2014, 3, e02445. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Yang, D.; Rupaimoole, R.; Pecot, C.V.; Sun, Y.; Mangala, L.S.; Li, X.; Ji, P.; Cogdell, D.; Hu, L.; et al. Augmentation of response to chemotherapy by microRNA-506 through regulation of RAD51 in serous ovarian cancers. J. Natl. Cancer Inst. 2015, 107, djv108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lodygin, D.; Tarasov, V.; Epanchintsev, A.; Berking, C.; Knyazeva, T.; Korner, H.; Knyazev, P.; Diebold, J.; Hermeking, H. Inactivation of miR−34a by aberrant CpG methylation in multiple types of cancer. Cell Cycle 2008, 7, 2591–2600. [Google Scholar] [CrossRef] [Green Version]
- Gasparini, P.; Lovat, F.; Fassan, M.; Casadei, L.; Cascione, L.; Jacob, N.K.; Carasi, S.; Palmieri, D.; Costinean, S.; Shapiro, C.L.; et al. Protective role of miR−155 in breast cancer through RAD51 targeting impairs homologous recombination after irradiation. Proc. Natl. Acad. Sci. USA 2014, 111, 4536–4541. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.; Wang, X.; McGregor, N.; Pienta, K.J.; Zhang, J. Dynamic regulation of Rad51 by E2F1 and p53 in prostate cancer cells upon drug-induced DNA damage under hypoxia. Mol. Pharmacol. 2014, 85, 866–876. [Google Scholar] [CrossRef] [Green Version]
- Stocker, M.; Le Nail, L.R.; De Belenet, H.; Wunder, J.S.; Andrulis, I.L.; Gokgoz, N.; Levy, N.; Mattei, J.C.; Olschwang, S. Inhibition of P53-mediated cell cycle control as the determinant in dedifferentiated liposarcomas development. Am. J. Cancer Res. 2021, 11, 3271–3284. [Google Scholar]
- Hine, C.M.; Li, H.; Xie, L.; Mao, Z.; Seluanov, A.; Gorbunova, V. Regulation of Rad51 promoter. Cell Cycle 2014, 13, 2038–2045. [Google Scholar] [CrossRef]
- Tessitore, A.; Cicciarelli, G.; Del Vecchio, F.; Gaggiano, A.; Verzella, D.; Fischietti, M.; Vecchiotti, D.; Capece, D.; Zazzeroni, F.; Alesse, E. MicroRNAs in the DNA Damage/Repair Network and Cancer. Int. J. Genom. 2014, 2014, 820248. [Google Scholar] [CrossRef] [Green Version]
- Li, E.; Xia, M.; Du, Y.; Long, K.; Ji, F.; Pan, F.; He, L.; Hu, Z.; Guo, Z. METTL3 promotes homologous recombination repair and modulates chemotherapeutic response in breast cancer by regulating the EGF/RAD51 axis. eLife 2022, 11, e75231. [Google Scholar] [CrossRef]
- Chen, W.T.; Tseng, H.Y.; Jiang, C.L.; Lee, C.Y.; Chi, P.; Chen, L.Y.; Lo, K.Y.; Wang, I.C.; Lin, F.J. Elp1 facilitates RAD51-mediated homologous recombination repair via translational regulation. J. Biomed. Sci. 2021, 28, 81. [Google Scholar] [CrossRef] [PubMed]
- Bae, T.; Jang, J.; Lee, H.; Song, J.; Chae, S.; Park, M.; Son, C.G.; Yoon, S.; Yoon, Y. Paeonia lactiflora root extract suppresses cancer cachexia by down-regulating muscular NF-kappaB signalling and muscle-specific E3 ubiquitin ligases in cancer-bearing mice. J. Ethnopharmacol. 2020, 246, 112222. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Annotation | |||
|---|---|---|---|---|
| Description | Fold Change | Normalized Read Count (log2) | ||
| QB/B | IB/B | |||
| RAD51 | RAD51 recombinase | 2.422 | 2.734 | 6.433 |
| ESCO2 | Establishment of sister chromatid cohesion N-acetyltransferase 2 | 2.351 | 2.691 | 7.394 |
| PARPBP | PARP1 binding protein | 2.100 | 2.532 | 6.244 |
| CDK1 | Cyclin-dependent kinase 1 | 2.061 | 2.736 | 7.082 |
| FANCD2 | Fanconi anemia, complementation group D2 | 1.975 | 2.622 | 7.983 |
| MND1 | Meiotic nuclear divisions 1 homolog | 1.954 | 2.939 | 6.457 |
| BRCA2 | Breast cancer 2, early onset | 1.948 | 2.796 | 6.681 |
| XRCC2 | X-ray repair complementing defective repair in Chinese hamster cells 2 | 1.915 | 2.405 | 6.988 |
| DTP | Denticleless E3 ubiquitin protein ligase homolog | 1.913 | 2.474 | 8.875 |
| POLQ | Polymerase (DNA directed), theta | 1.896 | 2.602 | 6.216 |
| CLSPN | claspin | 1.894 | 2.509 | 7.625 |
| TICRR | TOPBP1-interacting checkpoint and replication regulator | 1.854 | 2.213 | 6.882 |
| BRIP1 | BRCA1 interacting protein C−terminal helicase 1 | 1.839 | 2.458 | 7.377 |
| NEIL3 | Nei endonuclease VIII-like 3 | 1.829 | 2.282 | 6.896 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, M.; Jee, S.-C.; Shin, M.-K.; Han, D.-H.; Bu, K.-B.; Lee, S.-C.; Jang, B.-Y.; Sung, J.-S. Quercetin and Isorhamnetin Reduce Benzo[a]pyrene-Induced Genotoxicity by Inducing RAD51 Expression through Downregulation of miR−34a. Int. J. Mol. Sci. 2022, 23, 13125. https://doi.org/10.3390/ijms232113125
Kim M, Jee S-C, Shin M-K, Han D-H, Bu K-B, Lee S-C, Jang B-Y, Sung J-S. Quercetin and Isorhamnetin Reduce Benzo[a]pyrene-Induced Genotoxicity by Inducing RAD51 Expression through Downregulation of miR−34a. International Journal of Molecular Sciences. 2022; 23(21):13125. https://doi.org/10.3390/ijms232113125
Chicago/Turabian StyleKim, Min, Seung-Cheol Jee, Min-Kyoung Shin, Dong-Hee Han, Kyung-Bin Bu, Seung-Cheol Lee, Bo-Young Jang, and Jung-Suk Sung. 2022. "Quercetin and Isorhamnetin Reduce Benzo[a]pyrene-Induced Genotoxicity by Inducing RAD51 Expression through Downregulation of miR−34a" International Journal of Molecular Sciences 23, no. 21: 13125. https://doi.org/10.3390/ijms232113125
APA StyleKim, M., Jee, S. -C., Shin, M. -K., Han, D. -H., Bu, K. -B., Lee, S. -C., Jang, B. -Y., & Sung, J. -S. (2022). Quercetin and Isorhamnetin Reduce Benzo[a]pyrene-Induced Genotoxicity by Inducing RAD51 Expression through Downregulation of miR−34a. International Journal of Molecular Sciences, 23(21), 13125. https://doi.org/10.3390/ijms232113125