

LIF–IGF Axis Contributes to the Proliferation of Neural Progenitor Cells in Developing Rat Cerebrum
 , , ,
, , ,  , , and
, , and
Abstract
:1. Introduction
2. Results
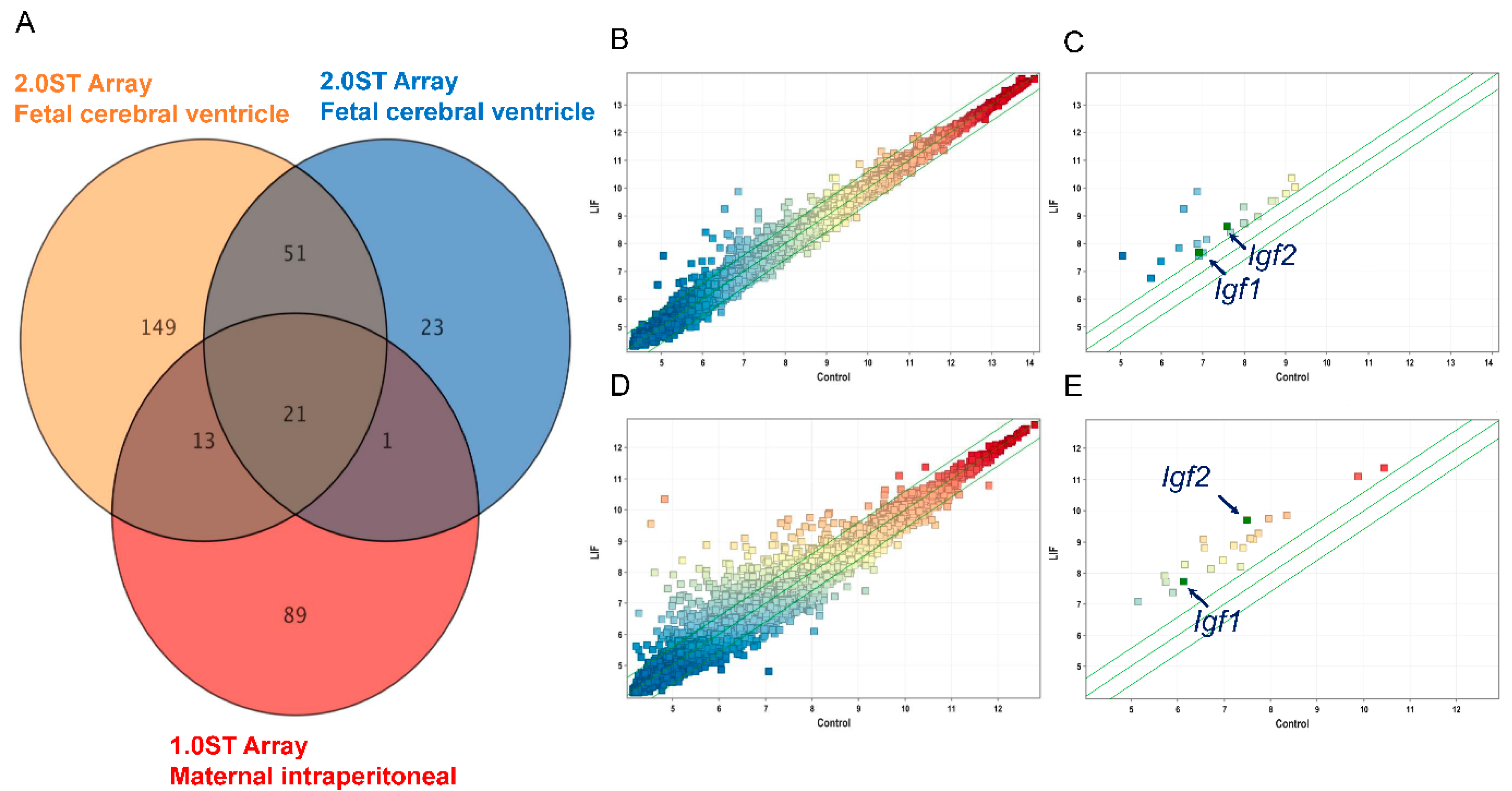
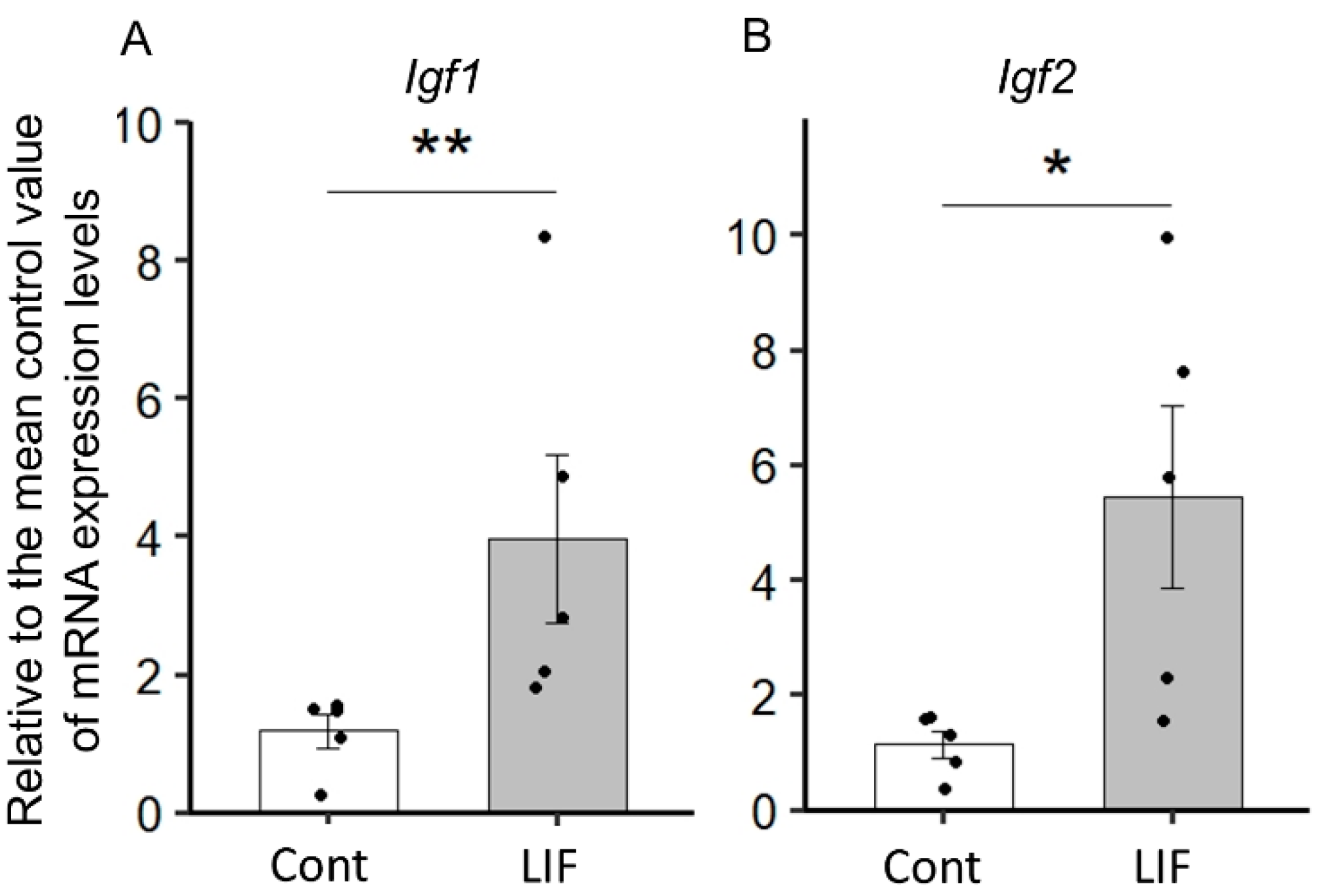
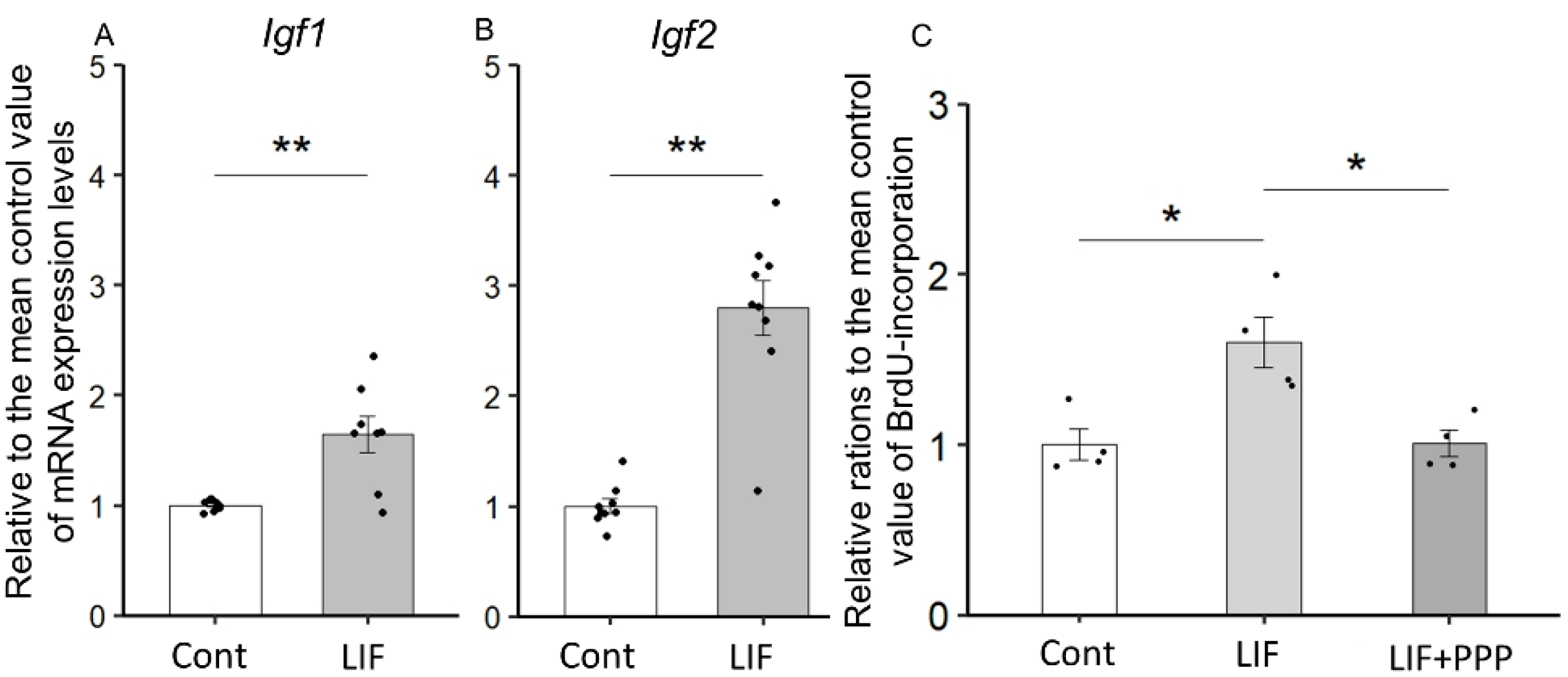
2.1. LIF Upregulates the Expression of Igf1 and Igf2 in the Dorsal Cerebrum
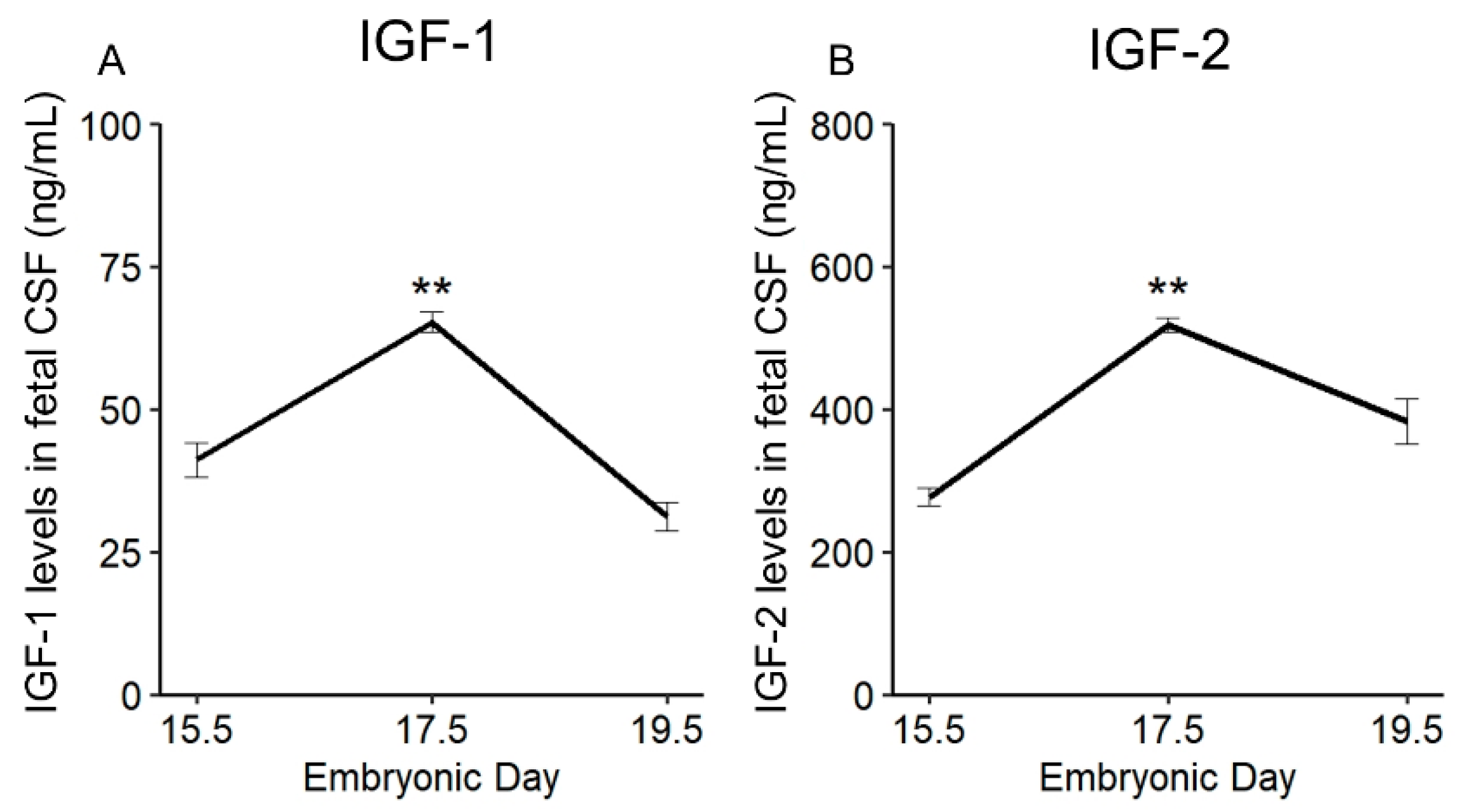
2.2. Physiological Change in IGF-1 and IGF-2 Levels in Fetal CSF
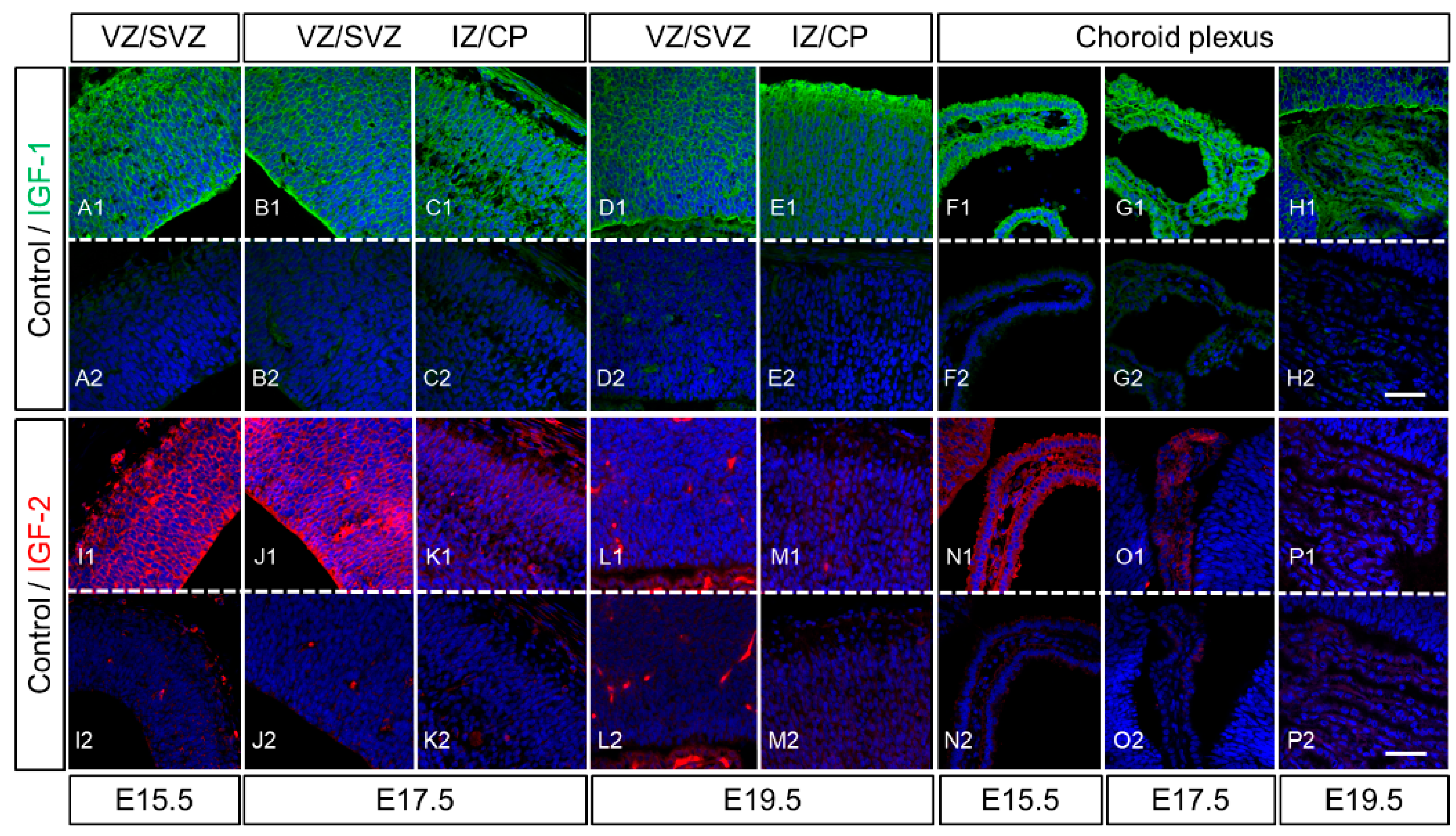
2.3. Expression of IGF-1 and IGF-2 in the Cerebrum of Rat Fetuses
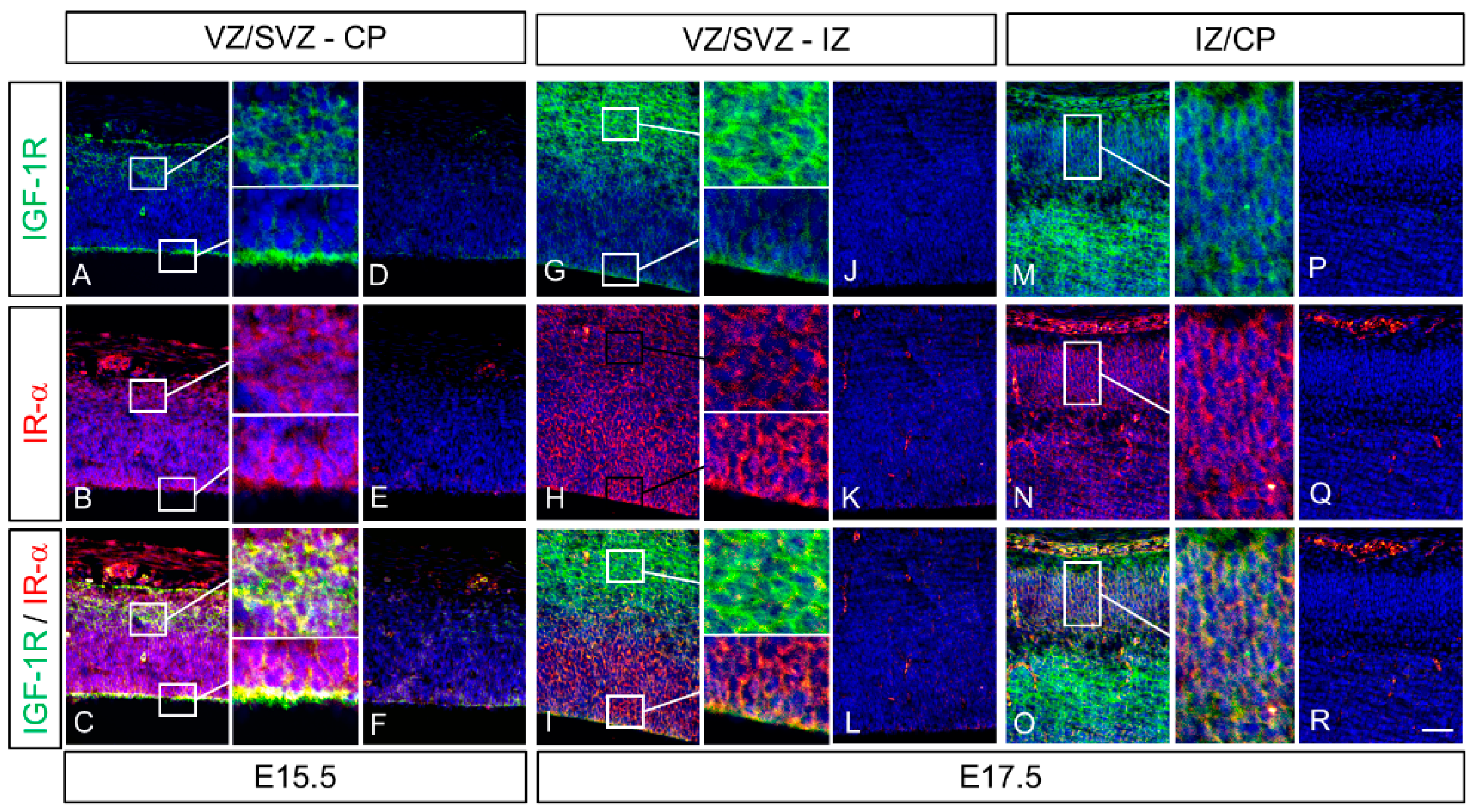
2.4. Colocalization of IGF-1 Receptor (IGF-1R) and Insulin Receptor α (IR-α) in the Cerebrum of Rat Fetuses
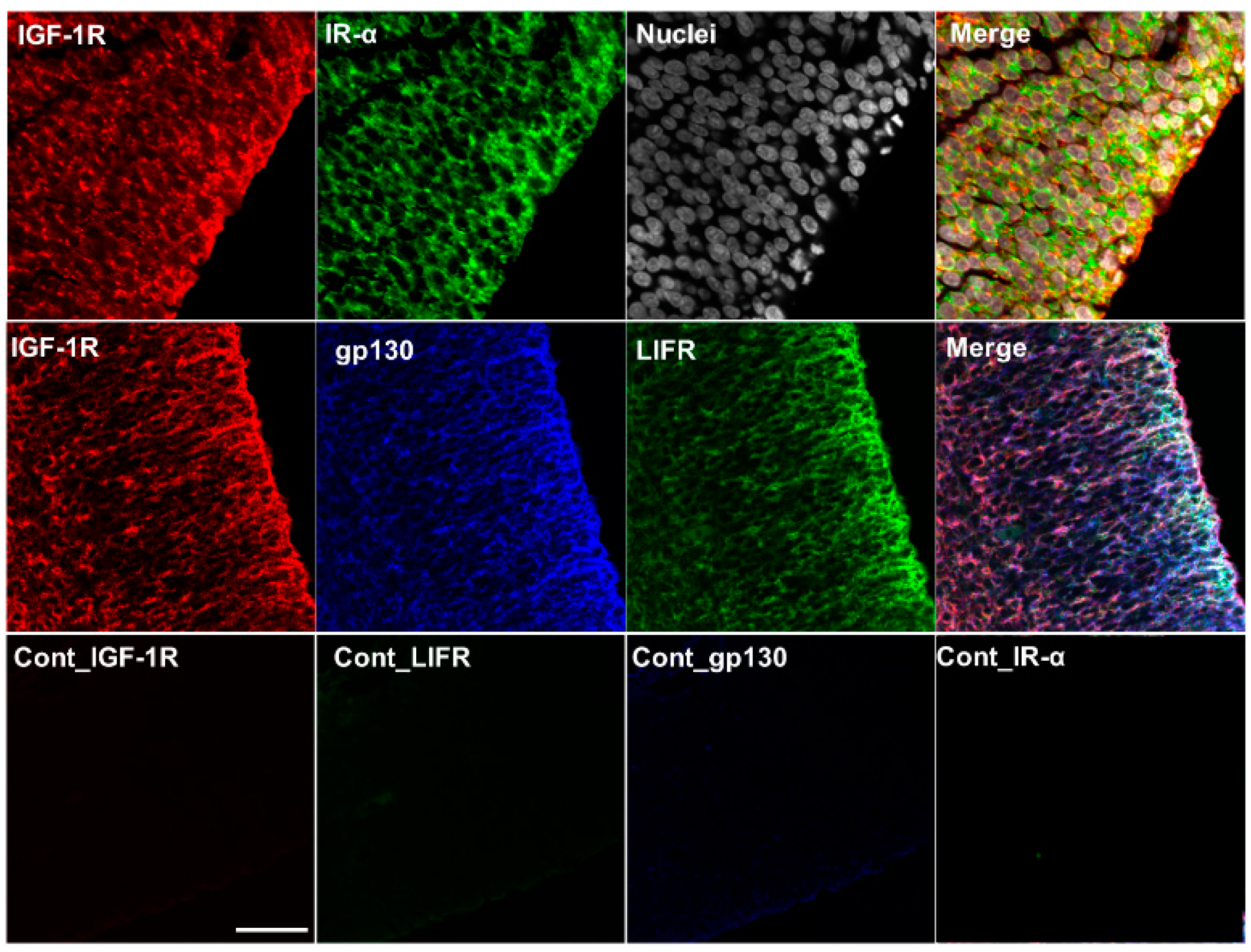
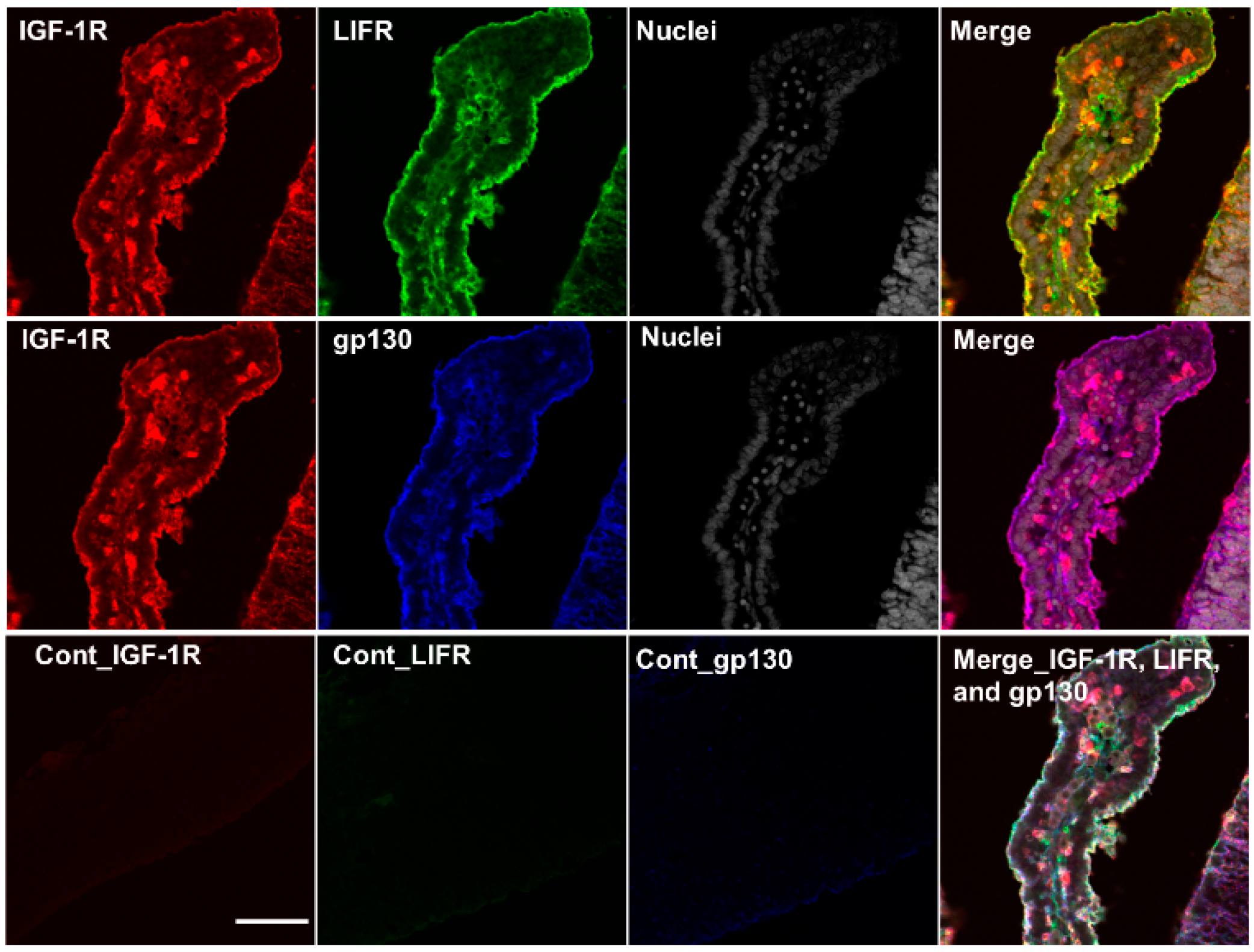
2.5. Colocalization of IGF-1R, IR-α, LIF Receptor (LIFR), and gp130 in the Cerebrum and Choroid Plexus of Rat Fetuses
2.6. LIF-Induced Proliferation of NPCs Is Dependent on IGF-1 or IGF-2
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Induction of IGF-1 and IGF-2 in the Fetal Cerebrum by LIF
4.3. Sample Preparation of Fetal CSF and Serum
4.4. RNA Preparation
4.5. DNA Microarray and Real-Time Quantitative PCR
4.6. ELISA
4.7. Fluorescence Immunohistochemistry
4.8. Neurosphere Assay
4.9. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bauer, S.; Patterson, P.H. Leukemia inhibitory factor promotes neural stem cell self-renewal in the adult brain. J. Neurosci. 2006, 26, 12089–12099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakashima, K.; Wiese, S.; Yanagisawa, M.; Arakawa, H.; Kimura, N.; Hisatsune, T.; Yoshida, K.; Kishimoto, T.; Sendtner, M.; Taga, T. Developmental requirement of gp130 signaling in neuronal survival and astrocyte differentiation. J. Neurosci. 1999, 19, 5429–5434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimazaki, T.; Shingo, T.; Weiss, S. The Ciliary Neurotrophic Factor/Leukemia Inhibitory Factor/gp130 Receptor Complex Operates in the Maintenance of Mammalian Forebrain Neural Stem Cells. J. Neurosci. 2001, 21, 7642–7653. [Google Scholar] [CrossRef]
- Pitman, M.; Emery, B.; Binder, M.; Wang, S.; Butzkueven, H.; Kilpatrick, T.J. LIF receptor signaling modulates neural stem cell renewal. Mol. Cell. Neurosci. 2004, 27, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Bonaguidi, M.A.; McGuire, T.; Hu, M.; Kan, L.; Samanta, J.; Kessler, J.A. LIF and BMP signaling generate separate and discrete types of GFAP-expressing cells. Development 2005, 132, 5503–5514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinrich, P.C.; Behrmann, I.; Müller-Newen, G.; Schaper, F.; Graeve, L. Interleukin-6-type cytokine signalling through the gp130/Jak/STAT pathway. Biochem. J. 1998, 334, 297–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taga, T.; Kishimoto, T. gp130 and the interleukin-6 family of cytokines. Annu. Rev. Immunol. 1997, 15, 797–819. [Google Scholar] [CrossRef]
- Chojnacki, A.; Shimazaki, T.; Gregg, C.; Weinmaster, G.; Weiss, S. Glycoprotein 130 signaling regulates Notch1 expression and activation in the self-renewal of mammalian forebrain neural stem cells. J. Neurosci. 2003, 23, 1730–1741. [Google Scholar] [CrossRef] [Green Version]
- Kamakura, S.; Oishi, K.; Yoshimatsu, T.; Nakafuku, M.; Masuyama, N.; Gotoh, Y. Hes binding to STAT3 mediates crosstalk between Notch and JAK-STAT signalling. Nat. Cell Biol. 2004, 6, 547–554. [Google Scholar] [CrossRef]
- Yoshimatsu, T.; Kawaguchi, D.; Oishi, K.; Takeda, K.; Akira, S.; Masuyama, N.; Gotoh, Y. Non-cell-autonomous action of STAT3 in maintenance of neural precursor cells in the mouse neocortex. Development 2006, 133, 2553–2563. [Google Scholar] [CrossRef]
- Hatta, T.; Moriyama, K.; Nakashima, K.; Taga, T.; Otani, H. The Role of gp130 in Cerebral Cortical Development: In Vivo Functional Analysis in a Mouse Exo Utero System. J. Neurosci. 2002, 22, 5516–5524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gregg, C.; Weiss, S. CNTF/LIF/gp130 receptor complex signaling maintains a VZ precursor differentiation gradient in the developing ventral forebrain. Development 2005, 132, 565–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatta, T.; Matsumoto, A.; Ono, A.; Udagawa, J.; Nimura, M.; Hashimoto, R.; Otani, H. Quantitative analyses of leukemia inhibitory factor in the cerebrospinal fluid in mouse embryos. Neuroreport 2006, 17, 1863–1866. [Google Scholar] [CrossRef] [PubMed]
- Simamura, E.; Shimada, H.; Higashi, N.; Uchishiba, M.; Otani, H.; Hatta, T. Maternal Leukemia Inhibitory Factor (LIF) promotes fetal neurogenesis via a LIF-ACTH-LIF signaling relay pathway. Endocrinology 2010, 151, 1853–1862. [Google Scholar] [CrossRef] [PubMed]
- Simamura, E.; Shimada, H.; Shoji, H.; Otani, H.; Hatta, T. Effects of melanocortins on fetal development. Congenit. Anom. 2011, 51, 47–54. [Google Scholar] [CrossRef]
- Tsukada, T.; Simamura, E.; Shimada, H.; Arai, T.; Higashi, N.; Akai, T.; Iizuka, H.; Hatta, T. The suppression of maternal-fetal leukemia inhibitory factor signal relay pathway by maternal immune activation impairs brain development in mice. PLoS ONE 2015, 10, e0129011. [Google Scholar] [CrossRef]
- Wang, H.; Tsukada, T.; Shimada, H.; Sakata-Haga, H.; Iida, Y.; Zhang, S.; Shoji, H.; Hatta, T. Leukemia inhibitory factor induces corticotropin-releasing hormone in mouse trophoblast stem cells. Biochem. Biophys. Res. Commun. 2020, 522, 81–87. [Google Scholar] [CrossRef]
- Wang, H.; Sakata-Haga, H.; Masuta, H.; Tomosugi, M.; Tsukada, T.; Shimada, H.; Sakai, D.; Shoji, H.; Hatta, T. Leukemia Inhibitory Factor Induces Proopiomelanocortin via CRH/CRHR Pathway in Mouse Trophoblast. Front. Cell Dev. Biol. 2021, 9, 1783. [Google Scholar] [CrossRef]
- Tsukada, T.; Shimada, H.; Sakata-Haga, H.; Iizuka, H.; Hatta, T. Molecular mechanisms underlying the models of neurodevelopmental disorders in maternal immune activation relevant to the placenta. Congenit. Anom. 2019, 59, 81–87. [Google Scholar] [CrossRef]
- Mairet-Coello, G.; Tury, A.; DiCicco-Bloom, E. Insulin-like growth factor-1 promotes G(1)/S cell cycle progression through bidirectional regulation of cyclins and cyclin-dependent kinase inhibitors via the phosphatidylinositol 3-kinase/Akt pathway in developing rat cerebral cortex. J. Neurosci. 2009, 29, 775–788. [Google Scholar] [CrossRef] [Green Version]
- Lehtinen, M.K.; Zappaterra, M.W.; Chen, X.; Yang, Y.J.; Hill, A.D.; Lun, M.; Maynard, T.; Gonzalez, D.; Kim, S.; Ye, P.; et al. The Cerebrospinal Fluid Provides a Proliferative Niche for Neural Progenitor Cells. Neuron 2011, 69, 893–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salehi, Z.; Mashayekhi, F.; Naji, M.; Pandamooz, S. Insulin-like growth factor-1 and insulin-like growth factor binding proteins in cerebrospinal fluid during the development of mouse embryos. J. Clin. Neurosci. 2009, 16, 950–953. [Google Scholar] [CrossRef] [PubMed]
- Thongkittidilok, C.; Tharasanit, T.; Sananmuang, T.; Buarpung, S.; Techakumphu, M. Insulin-like growth factor-1 (IGF-1) enhances developmental competence of cat embryos cultured singly by modulating the expression of its receptor (IGF-1R) and reducing developmental block. Growth Horm. IGF Res. 2014, 24, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Hakuno, F.; Takahashi, S. IGF1 receptor signaling pathways. J. Mol. Endocrinol. 2018, 61, T69–T86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez, A.M.; Torres-Alemán, I. The many faces of insulin-like peptide signalling in the brain. Nat. Rev. Neurosci. 2012, 13, 225–239. [Google Scholar] [CrossRef] [PubMed]
- O’Kusky, J.; Ye, P. Neurodevelopmental effects of insulin-like growth factor signaling. Front. Neuroendocrinol. 2012, 33, 230–251. [Google Scholar] [CrossRef] [Green Version]
- Dyer, A.H.; Vahdatpour, C.; Sanfeliu, A.; Tropea, D. The role of Insulin-Like Growth Factor 1 (IGF-1) in brain development, maturation and neuroplasticity. Neuroscience 2016, 325, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Lewitt, M.S.; Boyd, G.W. The Role of Insulin-Like Growth Factors and Insulin-Like Growth Factor–Binding Proteins in the Nervous System. Biochem. Insights 2019, 12, 117862641984217. [Google Scholar] [CrossRef] [Green Version]
- Constancia, M.; Hemberger, M.; Hughes, J.; Dean, W.; Ferguson-Smith, A.; Fundele, R.; Stewart, F.; Kelsey, G.; Fowden, A.; Sibley, C.; et al. Placental-specific IGF-II is a major modulator of placental and fetal growth. Nature 2002, 417, 945–948. [Google Scholar] [CrossRef]
- DeChiara, T.M.; Robertson, E.J.; Efstratiadis, A. Parental imprinting of the mouse insulin-like growth factor II gene. Cell 1991, 64, 849–859. [Google Scholar] [CrossRef]
- LeRoith, D.; Roberts, C.T., Jr. The insulin-like growth factor system and cancer. Cancer Lett. 2003, 195, 127–137. [Google Scholar] [CrossRef] [Green Version]
- Woods, K.A.; Camacho-Hübner, C.; Savage, M.O.; Clark, A.J. Intrauterine Growth Retardation and Postnatal Growth Failure Associated with Deletion of the Insulin-Like Growth Factor I Gene. N. Engl. J. Med. 1996, 335, 1363–1367. [Google Scholar] [CrossRef] [PubMed]
- Powell-Braxton, L.; Hollingshead, P.; Warburton, C.; Dowd, M.; Pitts-Meek, S.; Dalton, D.; Gillett, N.; Stewart, T.A. IGF-I is required for normal embryonic growth in mice. Genes Dev. 1993, 7, 2609–2617. [Google Scholar] [CrossRef] [PubMed]
- Baker, J.; Liu, J.P.; Robertson, E.J.; Efstratiadis, A. Role of insulin-like growth factors in embryonic and postnatal growth. Cell 1993, 75, 73–82. [Google Scholar] [CrossRef]
- Frasca, F.; Pandini, G.; Scalia, P.; Sciacca, L.; Mineo, R.; Costantino, A.; Goldfine, I.D.; Belfiore, A.; Vigneri, R. Insulin Receptor Isoform A, a Newly Recognized, High-Affinity Insulin-like Growth Factor II Receptor in Fetal and Cancer Cells. Mol. Cell. Biol. 1999, 19, 3278–3288. [Google Scholar] [CrossRef] [Green Version]
- Belfiore, A.; Frasca, F.; Pandini, G.; Sciacca, L.; Vigneri, R. Insulin Receptor Isoforms and Insulin Receptor/Insulin-Like Growth Factor Receptor Hybrids in Physiology and Disease. Endocr. Rev. 2009, 30, 586–623. [Google Scholar] [CrossRef] [Green Version]
- Mazitschek, R.; Giannis, A. Inhibitors of angiogenesis and cancer-related receptor tyrosine kinases. Curr. Opin. Chem. Biol. 2004, 8, 432–441. [Google Scholar] [CrossRef]
- Otaegi, G.; Yusta-Boyo, M.J.; Vergaño-Vera, E.; Méndez-Gómez, H.R.; Carrera, A.C.; Abad, J.L.; González, M.; de la Rosa, E.J.; Vicario-Abejón, C.; de Pablo, F. Modulation of the PI 3-kinase-Akt signalling pathway by IGF-I and PTEN regulates the differentiation of neural stem/precursor cells. J. Cell Sci. 2006, 119, 2739–2748. [Google Scholar] [CrossRef] [Green Version]
- Hurtado-Chong, A.; Yusta-Boyo, M.J.; Vergaño-Vera, E.; Bulfone, A.; De Pablo, F.; Vicario-Abejón, C. IGF-I promotes neuronal migration and positioning in the olfactory bulb and the exit of neuroblasts from the subventricular zone. Eur. J. Neurosci. 2009, 30, 742–755. [Google Scholar] [CrossRef]
- Gilbert, J.; Man, H.Y. Fundamental Elements in Autism: From Neurogenesis and Neurite Growth to Synaptic Plasticity. Front. Cell. Neurosci. 2017, 11, 359. [Google Scholar] [CrossRef]
- Brown, A.S. Epidemiologic Studies of Exposure to Prenatal Infection and Risk of Schizophrenia and Autism. Dev. Neurobiol. 2012, 72, 1272–1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.L.; Hsiao, E.Y.; Yan, Z.; Mazmanian, S.K.; Patterson, P.H. The placental interleukin-6 signaling controls fetal brain development and behavior. Brain Behav. Immun. 2017, 62, 11–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kreitz, S.; Zambon, A.; Ronovsky, M.; Budinsky, L.; Helbich, T.H.; Sideromenos, S.; Ivan, C.; Konerth, L.; Wank, I.; Berger, A.; et al. Maternal immune activation during pregnancy impacts on brain structure and function in the adult offspring. Brain. Behav. Immun 2020, 83, 56–67. [Google Scholar] [CrossRef] [PubMed]
- Barker, D.J.P.; Osmond, C.; Winter, P.D.; Margetts, B.; Simmonds, S.J. Weight in infancy and death from ischaemic heart disease. Lancet 1989, 2, 577–580. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Gene Expression |
|---|---|
| (LIF/Control) | |
| Igf2 | 3.901060583 |
| Dcn | 3.608841573 |
| Egfl6 | 3.442054291 |
| Pdgfra | 3.115182075 |
| Gpc3 | 2.970747151 |
| Dab2 | 2.795991831 |
| Bgn | 2.752705096 |
| Postn | 2.728830926 |
| Ddr2 | 2.680990235 |
| Serpinf1 | 2.672903839 |
| Lum | 2.62812436 |
| Anxa6 | 2.566506249 |
| Ogn | 2.512830911 |
| Ror1 | 2.450680042 |
| Anxa2 | 2.421598147 |
| Igf1 | 2.401063962 |
| H19|Mir675 | 2.28898765 |
| Ctsc | 2.224476544 |
| Lgals1 | 2.174736479 |
| S100a11 | 1.98069301 |
| Colec12 | 1.892572836 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takata, S.; Sakata-Haga, H.; Shimada, H.; Tsukada, T.; Sakai, D.; Shoji, H.; Tomosugi, M.; Nakamura, Y.; Ishigaki, Y.; Iizuka, H.; et al. LIF–IGF Axis Contributes to the Proliferation of Neural Progenitor Cells in Developing Rat Cerebrum. Int. J. Mol. Sci. 2022, 23, 13199. https://doi.org/10.3390/ijms232113199
Takata S, Sakata-Haga H, Shimada H, Tsukada T, Sakai D, Shoji H, Tomosugi M, Nakamura Y, Ishigaki Y, Iizuka H, et al. LIF–IGF Axis Contributes to the Proliferation of Neural Progenitor Cells in Developing Rat Cerebrum. International Journal of Molecular Sciences. 2022; 23(21):13199. https://doi.org/10.3390/ijms232113199
Chicago/Turabian StyleTakata, Sho, Hiromi Sakata-Haga, Hiroki Shimada, Tsuyoshi Tsukada, Daisuke Sakai, Hiroki Shoji, Mitsuhiro Tomosugi, Yuka Nakamura, Yasuhito Ishigaki, Hideaki Iizuka, and et al. 2022. "LIF–IGF Axis Contributes to the Proliferation of Neural Progenitor Cells in Developing Rat Cerebrum" International Journal of Molecular Sciences 23, no. 21: 13199. https://doi.org/10.3390/ijms232113199
APA StyleTakata, S., Sakata-Haga, H., Shimada, H., Tsukada, T., Sakai, D., Shoji, H., Tomosugi, M., Nakamura, Y., Ishigaki, Y., Iizuka, H., Hayashi, Y., & Hatta, T. (2022). LIF–IGF Axis Contributes to the Proliferation of Neural Progenitor Cells in Developing Rat Cerebrum. International Journal of Molecular Sciences, 23(21), 13199. https://doi.org/10.3390/ijms232113199