
The Organogermanium Compound 3-(Trihydroxygermyl) Propanoic Acid (THGP) Suppresses Inflammasome Activation Via Complexation with ATP

Abstract
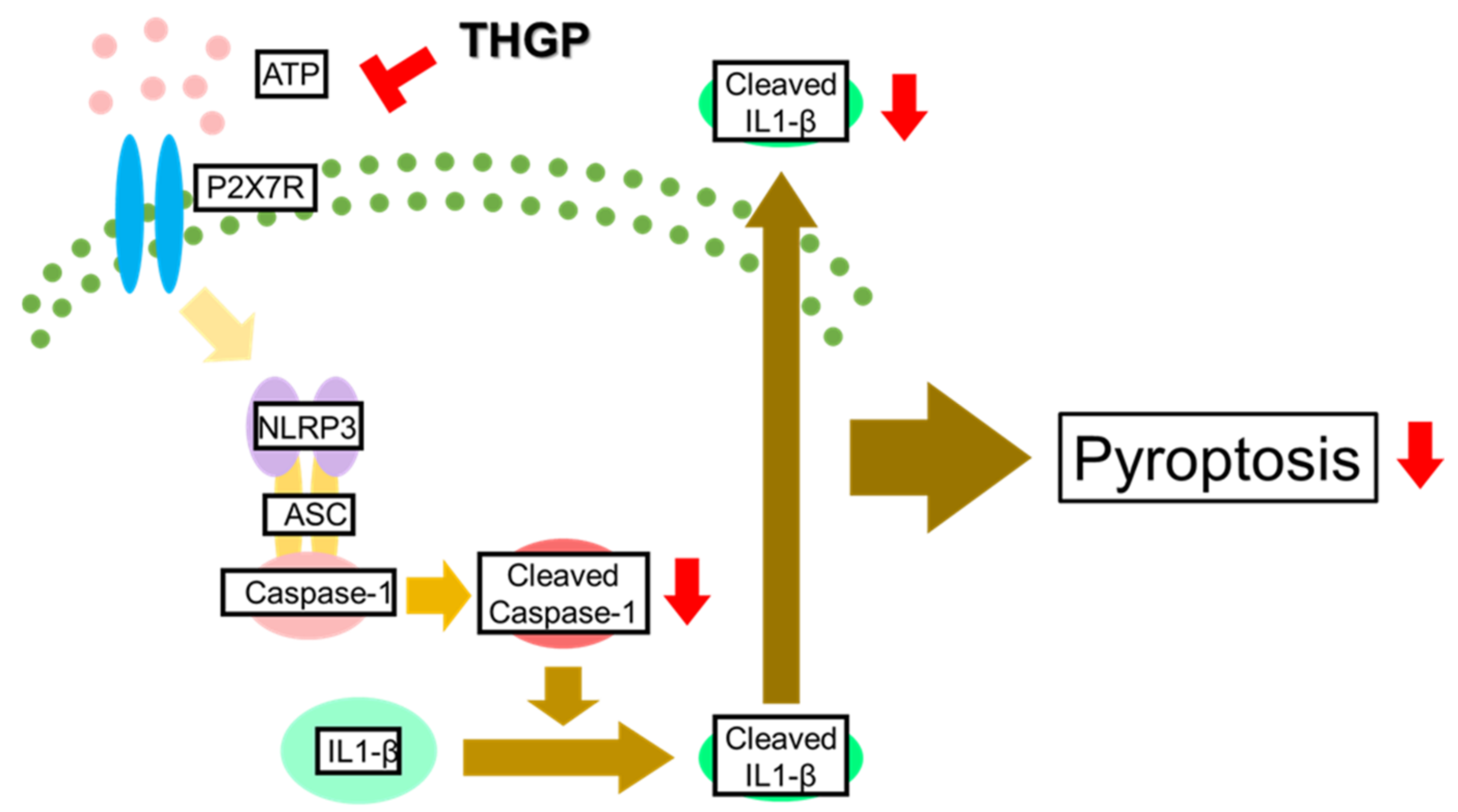
:1. Introduction
2. Results
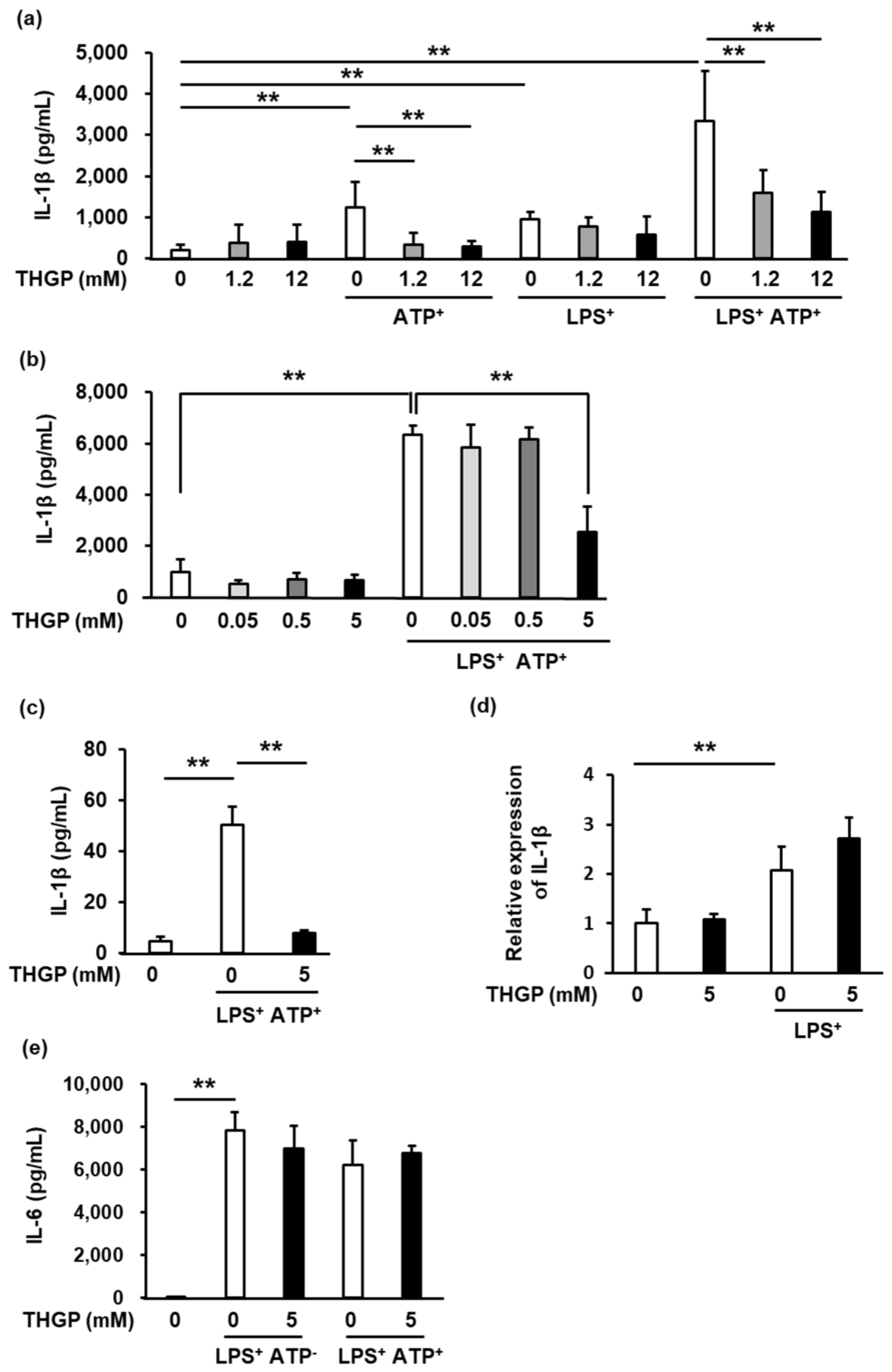
2.1. IL-1β Secretion from THP-1 Cells Was Suppressed by THGP
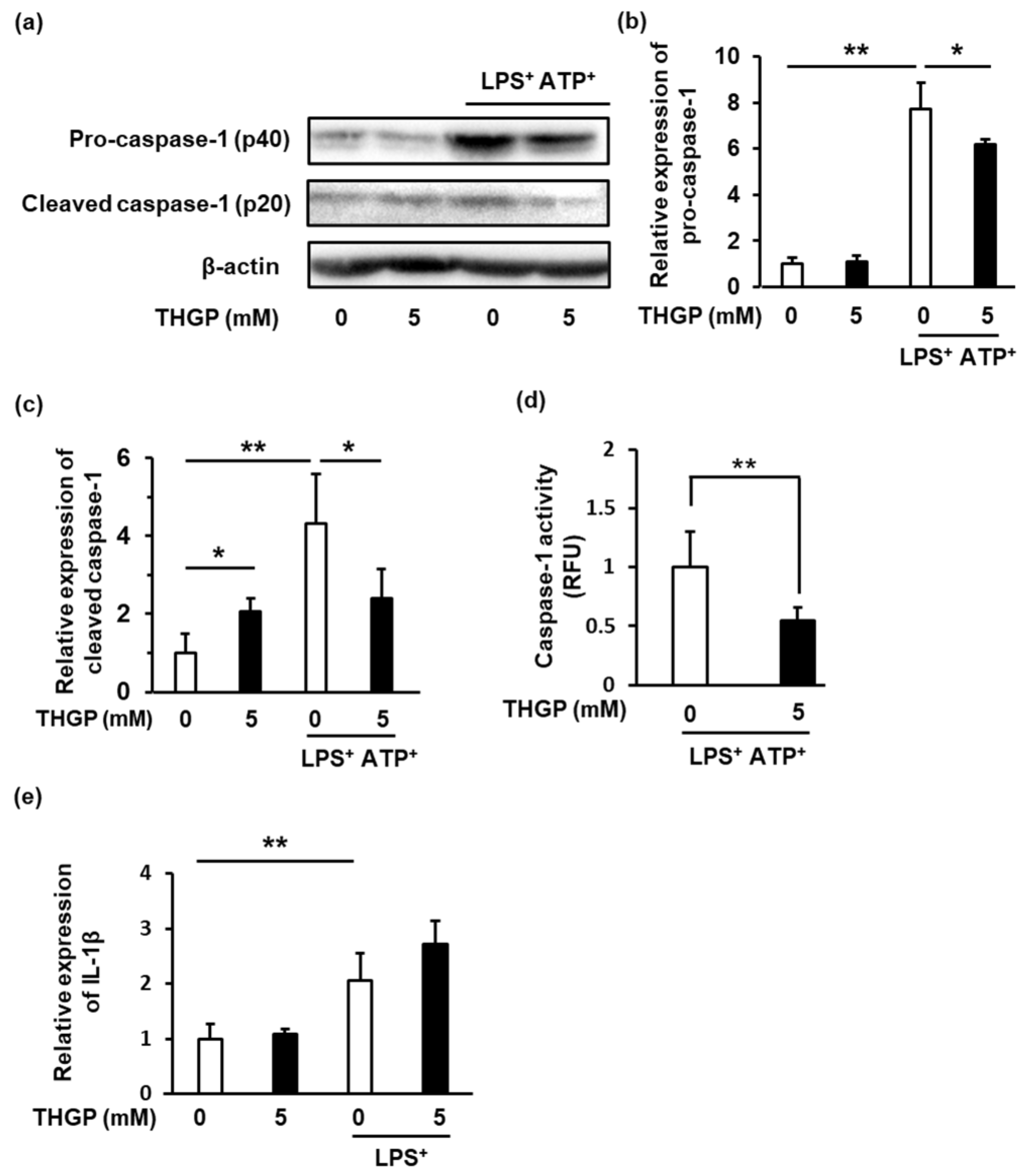
2.2. Caspase-1 Activity Was Suppressed by THGP
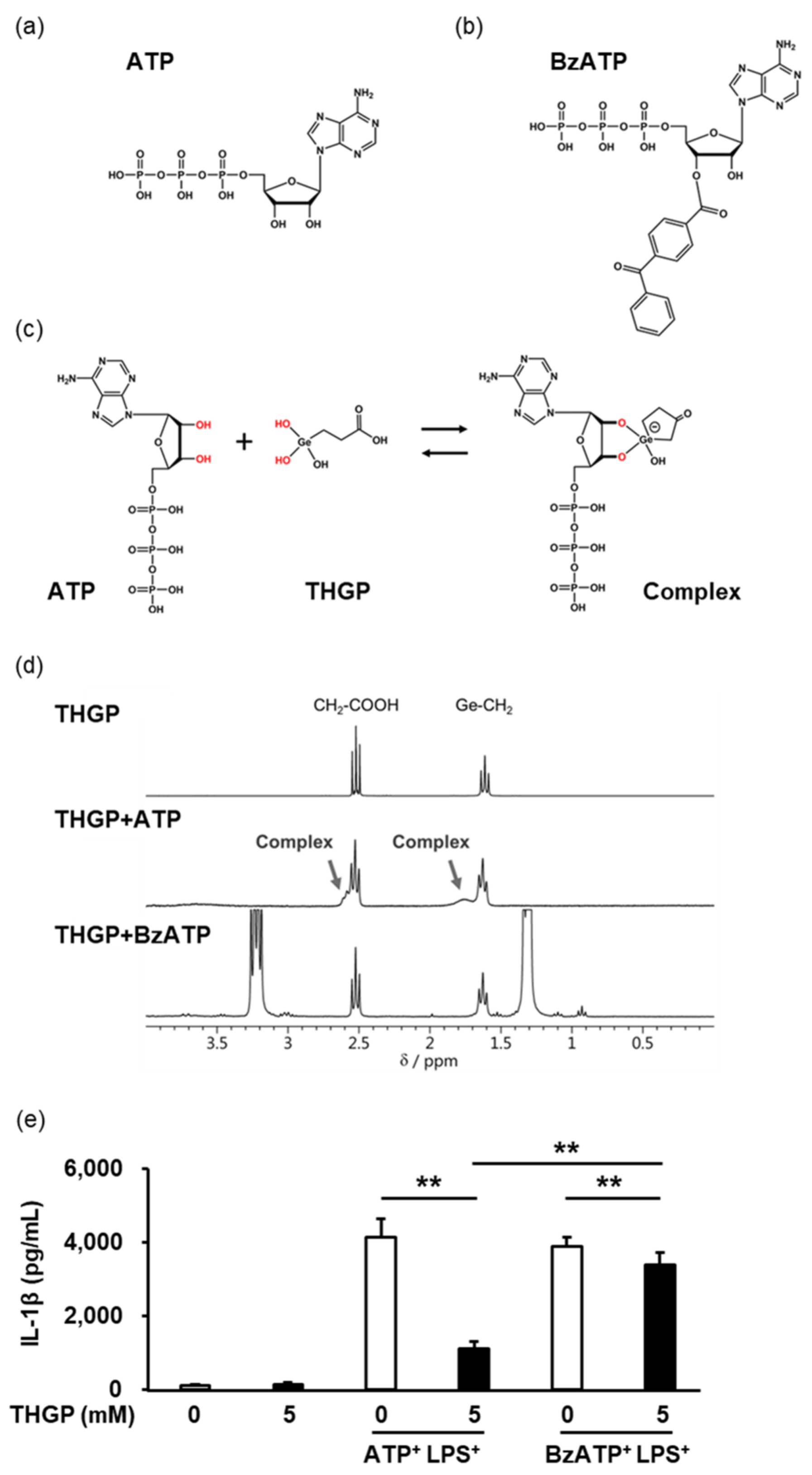
2.3. THGP Suppressed Inflammasome Activity by Forming a Complex with ATP
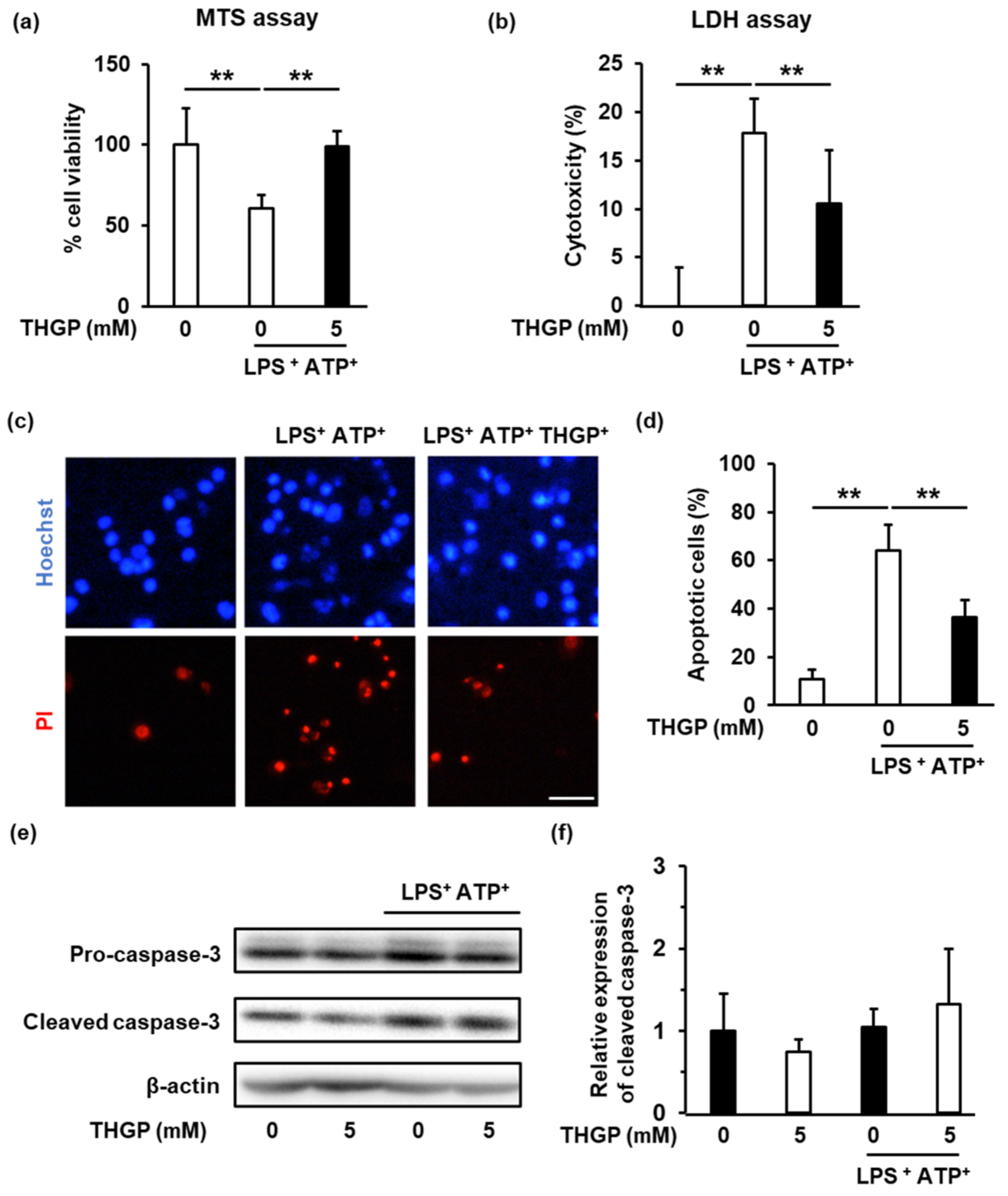
2.4. THGP Suppressed The Induction of Pyroptosis by LPS/ATP Stimulation
3. Discussion
4. Materials and Methods
4.1. Preparation of THGP
4.2. Cell Culture
4.3. ELISA
4.4. Western Blotting
4.5. Caspase-1 Activity
4.6. H-NMR
4.7. Real Time RT–PCR
4.8. Cell Viability Assay (MTS Assay)
4.9. LDH Assay
4.10. PI-Hoechst Staining
4.11. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gross, O.; Thomas, C.J.; Guarda, G.; Tschopp, J. The Inflammasome: An Integrated View. Immunol. Rev. 2011, 243, 136–151. [Google Scholar] [CrossRef] [PubMed]
- Guarda, G.; So, A. Regulation of Inflammasome Activity: Regulation of Inflammasome Activity. Immunology 2010, 130, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Kummer, J.A.; Broekhuizen, R.; Everett, H.; Agostini, L.; Kuijk, L.; Martinon, F.; van Bruggen, R.; Tschopp, J. Inflammasome Components NALP 1 and 3 Show Distinct but Separate Expression Profiles in Human Tissues Suggesting a Site-Specific Role in the Inflammatory Response. J. Histochem. Cytochem. 2006, 55, 443–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arend, W.P.; Palmer, G.; Gabay, C. IL-1, IL-18, and IL-33 Families of Cytokines. Immunol. Rev. 2008, 223, 20–38. [Google Scholar] [CrossRef] [PubMed]
- Van de Veerdonk, F.L.; Netea, M.G.; Dinarello, C.A.; Joosten, L.A.B. Inflammasome Activation and IL-1β and IL-18 Processing during Infection. Trends Immunol. 2011, 32, 110–116. [Google Scholar] [CrossRef]
- Cunha, C.; Carvalho, A.; Esposito, A.; Bistoni, F.; Romani, L. DAMP Signaling in Fungal Infections and Diseases. Front. Immunol. 2012, 3, 286. [Google Scholar] [CrossRef] [Green Version]
- Corriden, R.; Insel, P.A. Basal Release of ATP: An Autocrine-Paracrine Mechanism for Cell Regulation. Sci. Signal. 2010, 3, re1. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.Y.; Moon, J.H.; Lee, H.G.; Kim, S.U.; Lee, Y.B. ATP Released from Β-amyloid-Stimulated Microglia Induces Reactive Oxygen Species Production in an Autocrine Fashion. Exp. Mol. Med. 2007, 39, 820–827. [Google Scholar] [CrossRef]
- Martinon, F.; Burns, K.; Tschopp, J. The Inflammasome: A Molecular Platform Triggering Activation of Inflammatory Caspases and Processing of ProIL-Beta. Mol. Cell 2002, 10, 417–426. [Google Scholar] [CrossRef]
- Gabay, C.; Lamacchia, C.; Palmer, G. IL-1 Pathways in Inflammation and Human Diseases. Nat. Rev. Rheumatol. 2010, 6, 232–241. [Google Scholar] [CrossRef]
- Nakamura, T.; Takeda, T.; Tokuji, Y. The Oral Intake of Organic Germanium, Ge-132, Elevates α-Tocopherol Levels in the Plas-Ma and Modulates Hepatic Gene Expression Profiles to Promote Immune Activation in Mice. Int. J. Vitam. Nutr. Res. 2014, 84, 183–195. [Google Scholar] [CrossRef]
- Nakamura, T.; Nagura, T.; Akiba, M.; Sato, K.; Tokuji, Y.; Ohnishi, M.; Osada, K. Promotive Effects of the Dietary Organic Germanium Poly-Trans-[(2-Carboxyethyl) Germasesquioxane] (Ge-132) on the Secretion and Antioxidative Activity of Bile in Rodents. J. Health Sci. 2010, 56, 72–80. [Google Scholar] [CrossRef] [Green Version]
- Dozono, H.; Ikeda, K.; Onishi, T. Effectiveness of Ge-132 to Relieve Pain and Smooth Home Care Administration for the Terminal Cancer Patient. Gan Kagaku Ryoho. Cancer Chemother. 1996, 23, 291–295. [Google Scholar]
- Shimada, Y.; Sato, K.; Takeda, T.; Tokuji, Y. The Organogermanium Compound Ge-132 Interacts with Nucleic Acid Components and Inhibits the Catalysis of Adenosine Substrate by Adenosine Deaminase. Biol. Trace Elem. Res. 2018, 181, 164–172. [Google Scholar] [CrossRef]
- Shimada, Y.; Sato, K.; Masaki, M.; Nakamura, T.; Tokuji, Y. Quantitative Assessment of the Interactions between the Organogermanium Compound and Saccharides Using an NMR Reporter Molecule. Carbohydr. Res. 2021, 499, 108199. [Google Scholar] [CrossRef]
- Shimada, Y.; Sato, K.; Tokuji, Y.; Nakamura, T. Nuclear Magnetic Resonance Studies of the Interactions between the Organic Germanium Compound Ge-132 and Saccharides. Carbohydr. Res. 2015, 407, 10–15. [Google Scholar] [CrossRef]
- Nakamura, T.; Shimada, Y.; Takeda, T.; Sato, K.; Akiba, M.; Fukaya, H. Organogermanium Compound, Ge-132, Forms Complexes with Adrenaline, ATP and Other Physiological Cis-Diol Compounds. Future Med. Chem. 2015, 7, 1233–1246. [Google Scholar] [CrossRef]
- Miao, E.A.; Rajan, J.V.; Aderem, A. Caspase-1-Induced Pyroptotic Cell Death: Caspase-1-Induced Pyroptotic Cell Death. Immunol. Rev. 2011, 243, 206–214. [Google Scholar] [CrossRef] [Green Version]
- Fusco, R.; Siracusa, R.; Genovese, T.; Cuzzocrea, S.; Paola, R.D. Focus on the Role of NLRP3 Inflammasome in Diseases. Int. J. Mol. Sci. 2020, 21, 4223. [Google Scholar] [CrossRef]
- Groslambert, M.; Py, B.F. Spotlight on the NLRP3 Inflammasome Pathway. J. Inflamm. Res. 2018, 11, 359–374. [Google Scholar] [CrossRef] [Green Version]
- Tsutsui, M.; Kakimoto, N.; Axtell, D.D.; Oikawa, H.; Asai, K. Crystal Structure of “Carboxyethylgermanium Sesquioxide”. J. Am. Chem. Soc. 1976, 98, 8287–8289. [Google Scholar] [CrossRef]
- Gombault, A.; Baron, L.; Couillin, I. ATP Release and Purinergic Signaling in NLRP3 Inflammasome Activation. Front. Immunol. 2013, 3, 414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldwin, A.G.; Brough, D.; Freeman, S. Inhibiting the Inflammasome: A Chemical Perspective. J. Med. Chem. 2016, 59, 1691–1710. [Google Scholar] [CrossRef] [PubMed]
- Shao, B.-Z.; Xu, Z.-Q.; Han, B.-Z.; Su, D.-F.; Liu, C. NLRP3 Inflammasome and Its Inhibitors: A Review. Front. Pharmacol. 2015, 6, 262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Teng, G.; Zhou, H.; Dong, C. Germanium Reduces Inflammatory Damage in Mammary Glands During Lipopolysaccharide-Induced Mastitis in Mice. Biol. Trace Elem. Res. 2020, 198, 617–626. [Google Scholar] [CrossRef] [PubMed]
- Azumi, J.; Takeda, T.; Shimada, Y.; Aso, H.; Nakamura, T. The Organogermanium Compound THGP Suppresses Melanin Synthesis via Complex Formation with L-DOPA on Mushroom Tyrosinase and in B16 4A5 Melanoma Cells. Int. J. Mol. Sci. 2019, 20, 4785. [Google Scholar] [CrossRef] [Green Version]
- Belhadjer, Z.; Méot, M.; Bajolle, F.; Khraiche, D.; Legendre, A.; Abakka, S.; Auriau, J.; Grimaud, M.; Oualha, M.; Beghetti, M.; et al. Acute Heart Failure in Multisystem Inflammatory Syndrome in Children in the Context of Global SARS-CoV-2 Pandemic. Circulation 2020, 142, 429–436. [Google Scholar] [CrossRef]
- Ren, Y.; Shu, T.; Wu, D.; Mu, J.; Wang, C.; Huang, M.; Han, Y.; Zhang, X.-Y.; Zhou, W.; Qiu, Y.; et al. The ORF3a Protein of SARS-CoV-2 Induces Apoptosis in Cells. Cell. Mol. Immunol. 2020, 17, 881–883. [Google Scholar] [CrossRef]
- Siu, K.; Yuen, K.; Castano-Rodriguez, C.; Ye, Z.; Yeung, M.; Fung, S.; Yuan, S.; Chan, C.; Yuen, K.; Enjuanes, L.; et al. Severe Acute Respiratory Syndrome Coronavirus ORF3a Protein Activates the NLRP3 Inflammasome by Promoting TRAF3-dependent Ubiquitination of ASC. FASEB J. 2019, 33, 8865–8877. [Google Scholar] [CrossRef]
- Zhao, N.; Di, B.; Xu, L. The NLRP3 Inflammasome and COVID-19: Activation, Pathogenesis and Therapeutic Strategies. Cytokine Growth Factor Rev. 2021, 61, 2–15. [Google Scholar] [CrossRef]
- Chang, Y.-S.; Ko, B.-H.; Ju, J.-C.; Chang, H.-H.; Huang, S.-H.; Lin, C.-W. SARS Unique Domain (SUD) of Severe Acute Respiratory Syndrome Coronavirus Induces NLRP3 Inflammasome-Dependent CXCL10-Mediated Pulmonary Inflammation. Int. J. Mol. Sci. 2020, 21, 3179. [Google Scholar] [CrossRef]
- Narokha, V.; Nizhenkovska, I.; Kuznetsova, O. Potential of Germanium-Based Compounds in Coronavirus Infection. Acta Pharm. 2022, 72, 245–258. [Google Scholar] [CrossRef]
- Tezuka, T. Book Review of “The Secret of Organogermanium―Amazing Power to Renew Life”. Adv. Enzym. Res. 2021, 9, 50–53. [Google Scholar] [CrossRef]
- Aso, H.; Suzuki, F.; Ebina, T.; Ishida, N. Antiviral Activity of Carboxyethylgermanium Sesquioxide (Ge-132) in Mice Infected with Influenza Virus. J. Biol. Response Modif. 1989, 8, 180–189. [Google Scholar]
- Baidya, S.; Nishimoto, Y.; Sato, S.; Shimada, Y.; Sakurai, N.; Nonaka, H.; Noguchi, K.; Kido, M.; Tadano, S.; Ishikawa, K.; et al. Dual Effect of Organogermanium Compound THGP on RIG-I-Mediated Viral Sensing and Viral Replication during Influenza a Virus Infection. Viruses 2021, 13, 1674. [Google Scholar] [CrossRef]
- Shigeru, A.; Miyoko, Y.; Kaori, I. Improved Rheumatoid Arthritis Case with Ge-132 Administration Evaluated by Clinically and Immunologically Using Two-Color Flow Cytometry. Jpn. J. Clin. Immunol. 1990, 13, 80–86. [Google Scholar] [CrossRef] [Green Version]
- Wakayashi, Y. Effect of Germanium-132 on Low-Density Lipoprotein Oxidation and Atherosclerosis in Kurosawa and Kusanagi Hypercholesterolemic Rabbits. Biosci. Biotechnol. Biochem. 2014, 65, 1893–1896. [Google Scholar] [CrossRef]
- Aso, H.; Suzuki, F.; Yamagughi, T.; Hayashi, Y.; Ebina, T.; Ishida, N. Induction of Interferon and Activation of NK Cells and Macrophages in Mice by Oral Administration of Ge-132, an Organic Germanium Compound. Microbiol. Immunol. 1985, 29, 65–74. [Google Scholar] [CrossRef] [Green Version]
- Munakata, T.; Arai, S.; Kuwano, K.; Furukawa, M.; Tomita, Y. Induction of Interferon Production by Natural Killer Cells by Organogermanium Compound, Gel32. J. Interferon Res. 1987, 7, 69–76. [Google Scholar] [CrossRef]
- Ngoungoure, F.P.; Owona, B.A. Withaferin A Modulates AIM2 Inflammasome and Caspase-1 Expression in THP-1 Polarized Macrophages. Exp. Cell Res. 2019, 383, 111564. [Google Scholar] [CrossRef]
- Zheng, J.; Yang, L.; Deng, Y.; Zhang, C.; Zhang, Y.; Xiong, S.; Ding, C.; Zhao, J.; Liao, C.; Gong, D. A Review of Public and Environmental Consequences of Organic Germanium. Crit. Rev. Environ. Sci. Technol. 2019, 50, 1384–1409. [Google Scholar] [CrossRef]
- Nakamura, T.; Nagura, T.; Sato, K.; Ohnishi, M. Evaluation of the Effects of Dietary Organic Germanium, Ge-132, and Raffinose Supplementation on Caecal Flora in Rats. Biosci. Microbiota Food Health 2012, 31, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Sugiya, Y.; Sakamaki, S.; Sugita, T.; Abo, Y.; Satoh, H. Subacute Oral Toxicity of Carboxyethylgermanium Sesquioxide (Ge-132) in Rats. Pharmacometrics 1986, 6, 1181–1190. [Google Scholar]
- Sugiya, Y.; Sakamaki, S.; Satoh, H. Reversibility Study in Rats after Two Months’Oral Administration of Carboxyethylgermanium Sesquioxide (Ge-132) in Rats. Pharmacometrics 1986, 6, 1191–1200. [Google Scholar]
- Iwadate, K.; Yamaguchi, Y.; Sasaki, M.; Nakatani, M.; Doi, Y.; Imai, N.; Tamano, S.; Nishihori, Y. Carcinogenicity Study of Poly-Trans-[(2-Carboxyethyl)Germasesquioxane] (Ge-132) in F344 Rats. Fundam. Toxicol. Sci. 2018, 5, 127–140. [Google Scholar] [CrossRef] [Green Version]
- Doi, Y.; Imai, N.; Suguro, M.; Numano, T.; Furukawa, F. No Carcinogenicity of Poly-Trans-[(2-Carboxyethyl) Germasesquioxane] (Ge-132): 26-Week Feeding Study Using RasH2 Mice. Fundam. Toxicol. Sci. 2017, 4, 137–150. [Google Scholar] [CrossRef]
- Miyao, K.; Onishi, T.; Asai, K.; Tomizawa, S.; Suzuki, F. Toxicology and Phase I Studies on a Novel Organogermanium Compound, Ge-132. Curr. Chemother Infec. Dis. 1979, 2, 1527–1529. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forward | Reverse | |
|---|---|---|
| Il-1β | TTACAGTGGCAATGAGGATGAC | GTCGGAGATTCGTAGCTGGAT |
| Rps18 | TCAGCCTCTTCTCCTTCCTG | GGCTACAGGCTTGTCACTCG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azumi, J.; Shimada, Y.; Takeda, T.; Aso, H.; Nakamura, T. The Organogermanium Compound 3-(Trihydroxygermyl) Propanoic Acid (THGP) Suppresses Inflammasome Activation Via Complexation with ATP. Int. J. Mol. Sci. 2022, 23, 13364. https://doi.org/10.3390/ijms232113364
Azumi J, Shimada Y, Takeda T, Aso H, Nakamura T. The Organogermanium Compound 3-(Trihydroxygermyl) Propanoic Acid (THGP) Suppresses Inflammasome Activation Via Complexation with ATP. International Journal of Molecular Sciences. 2022; 23(21):13364. https://doi.org/10.3390/ijms232113364
Chicago/Turabian StyleAzumi, Junya, Yasuhiro Shimada, Tomoya Takeda, Hisashi Aso, and Takashi Nakamura. 2022. "The Organogermanium Compound 3-(Trihydroxygermyl) Propanoic Acid (THGP) Suppresses Inflammasome Activation Via Complexation with ATP" International Journal of Molecular Sciences 23, no. 21: 13364. https://doi.org/10.3390/ijms232113364
APA StyleAzumi, J., Shimada, Y., Takeda, T., Aso, H., & Nakamura, T. (2022). The Organogermanium Compound 3-(Trihydroxygermyl) Propanoic Acid (THGP) Suppresses Inflammasome Activation Via Complexation with ATP. International Journal of Molecular Sciences, 23(21), 13364. https://doi.org/10.3390/ijms232113364