
Topical Administration of a Novel Acetylated Tetrapeptide Suppresses Vascular Permeability and Immune Responses and Alleviates Atopic Dermatitis in a Murine Model


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
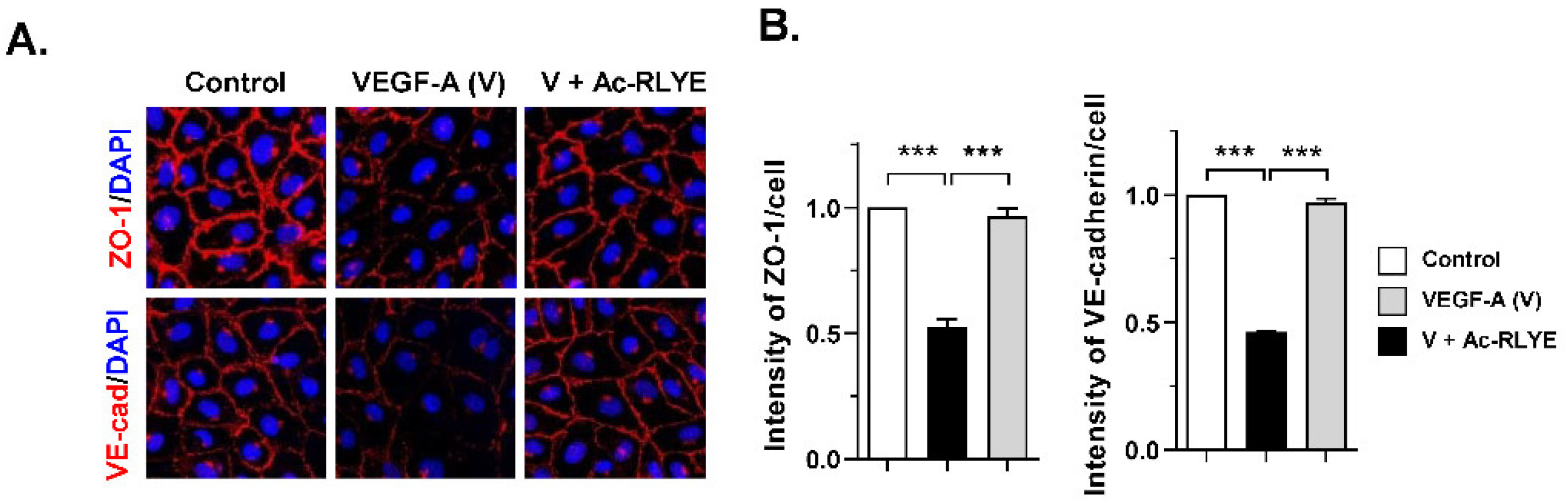
2.1. Ac-RLYE Blocks Vascular Endothelial Growth Factor-Induced Permeability in HUVECs
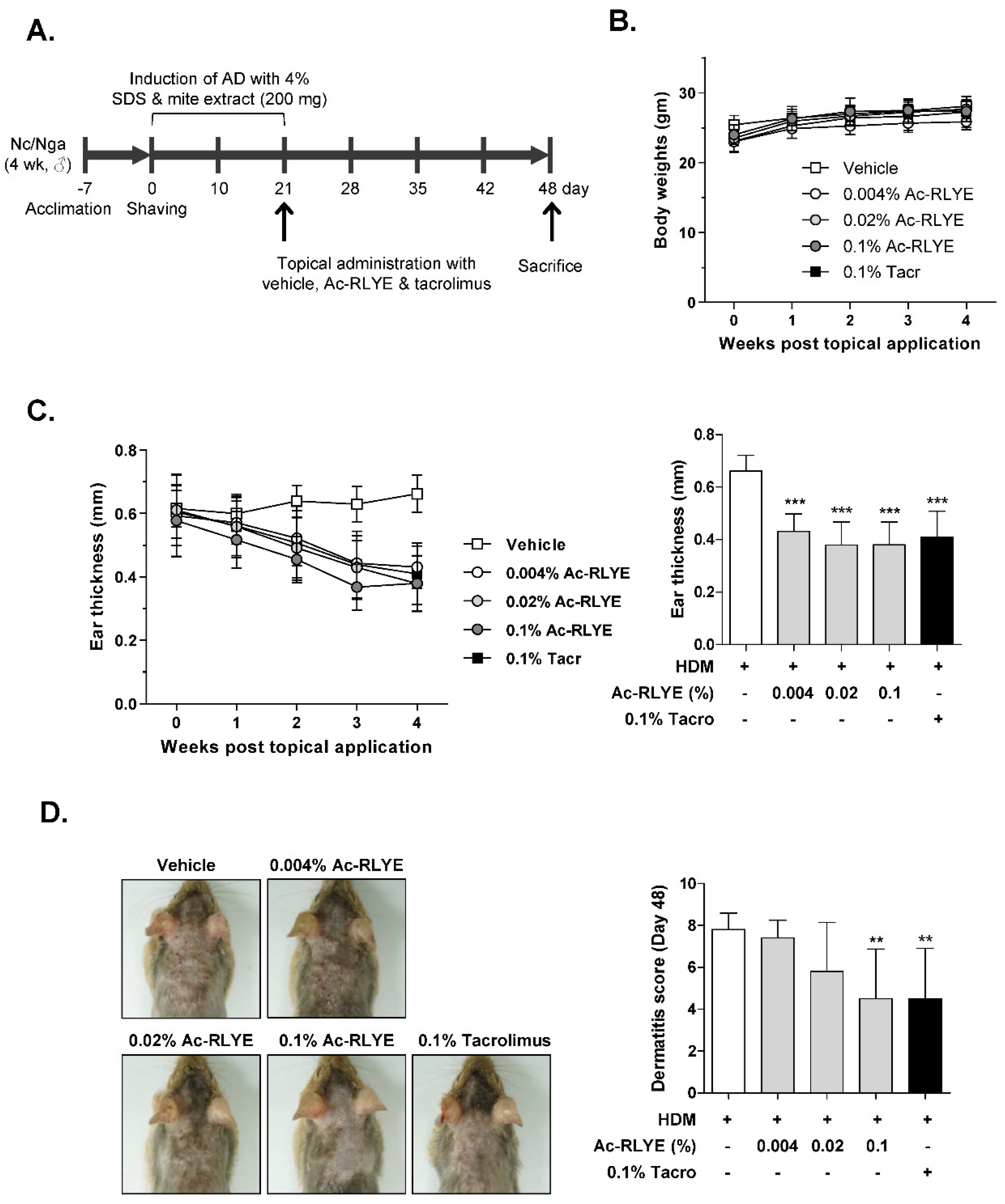
2.2. Ac-RLYE Alleviated AD Symptoms in HDM-Challenged NC/Nga Mice
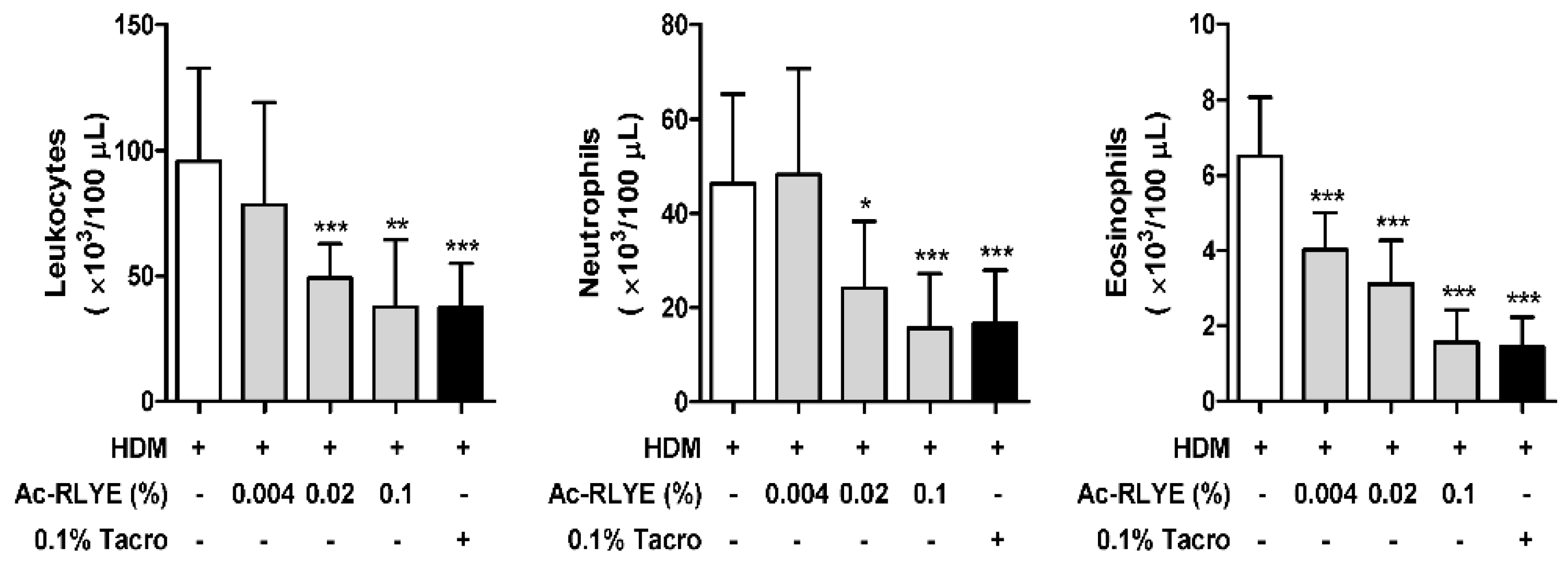
2.3. Ac-RLYE Lowered the Levels of White Blood Cells (WBCs) in the Blood of HDM-Challenged NC/Nga Mice
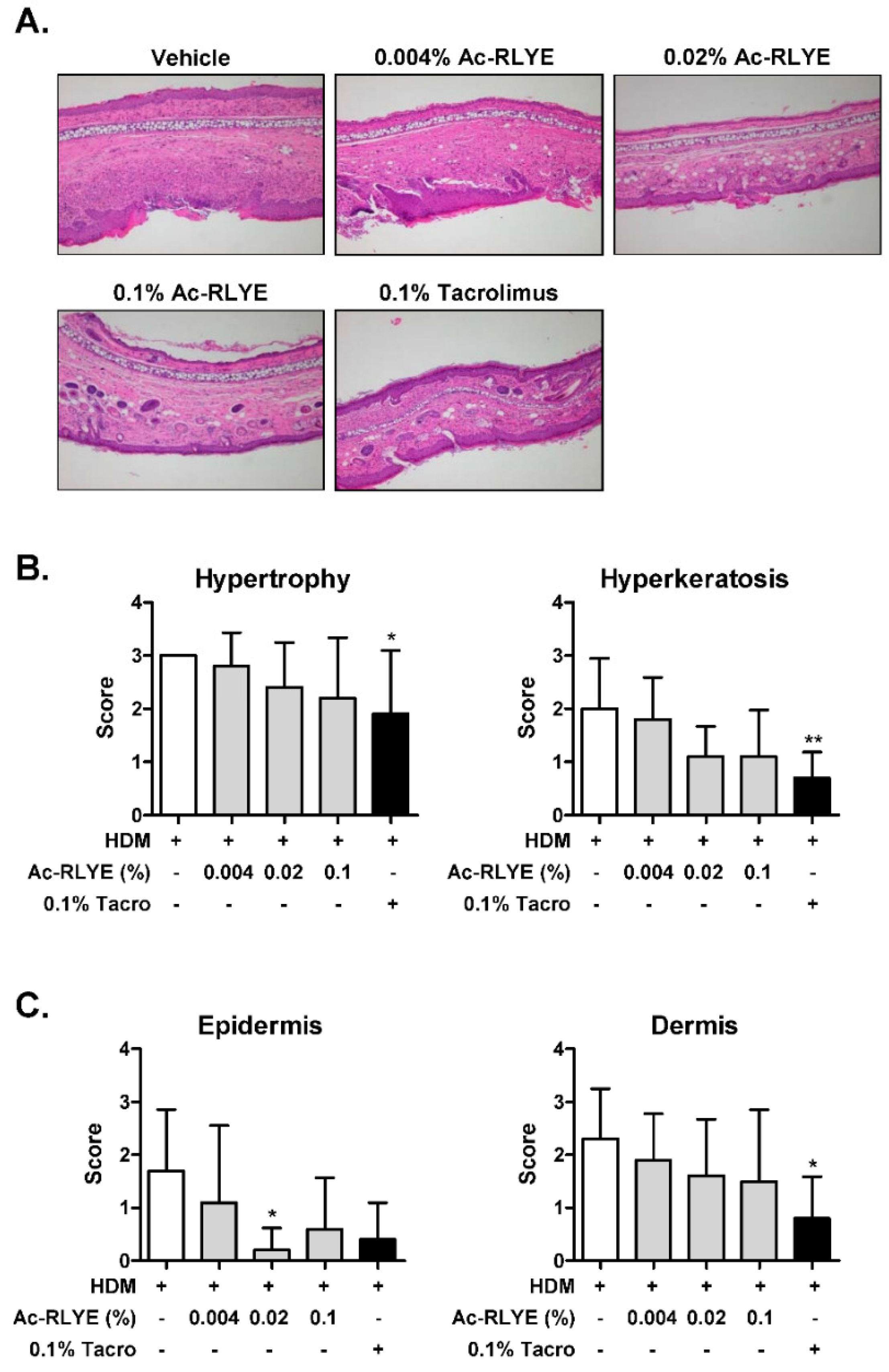
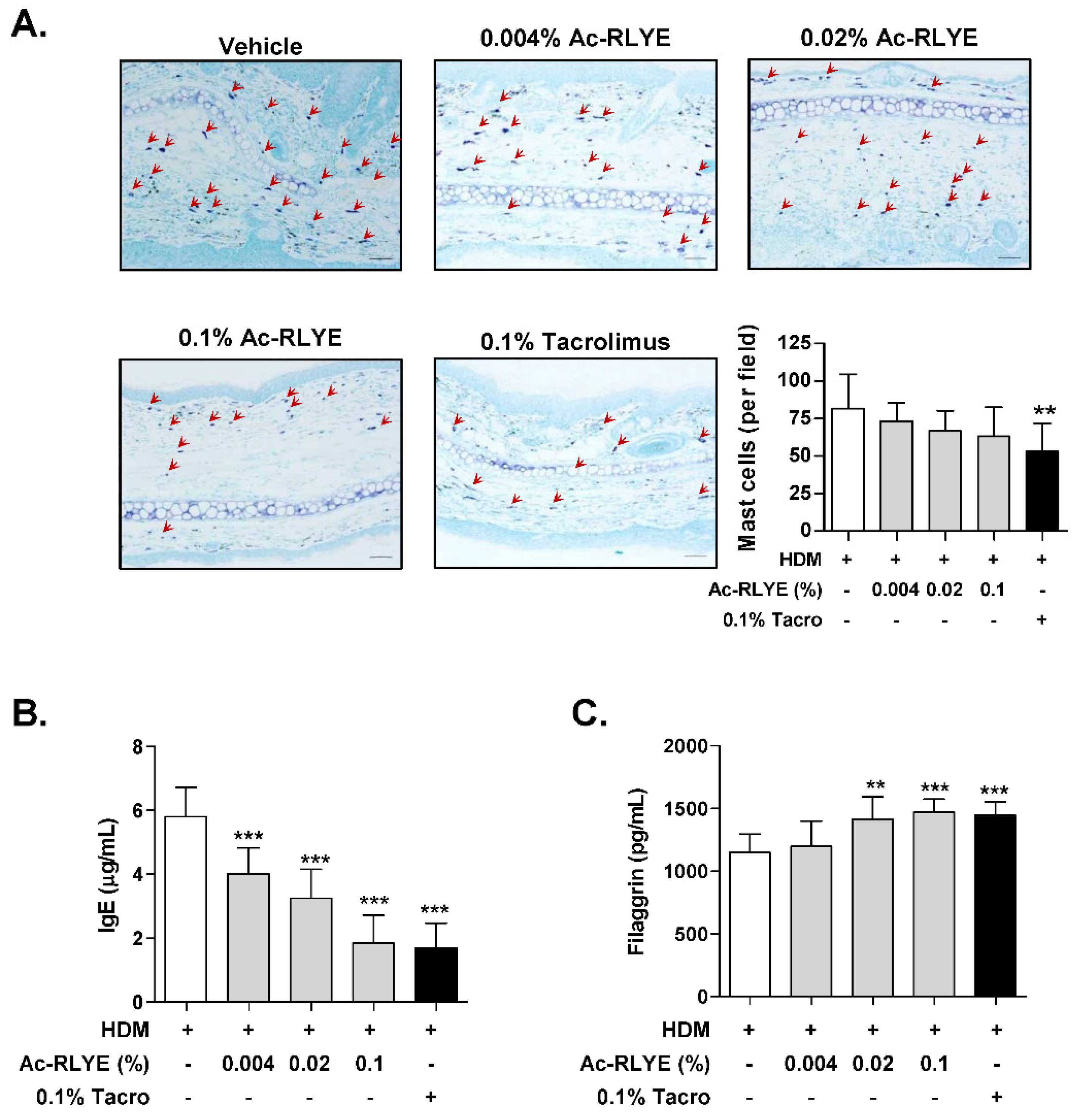
2.4. Ac-RLYE Reduced Mast Cell Infiltration and Plasma IgE
2.5. Ac-RLYE Restored Skin Barrier Function-Related Protein Level
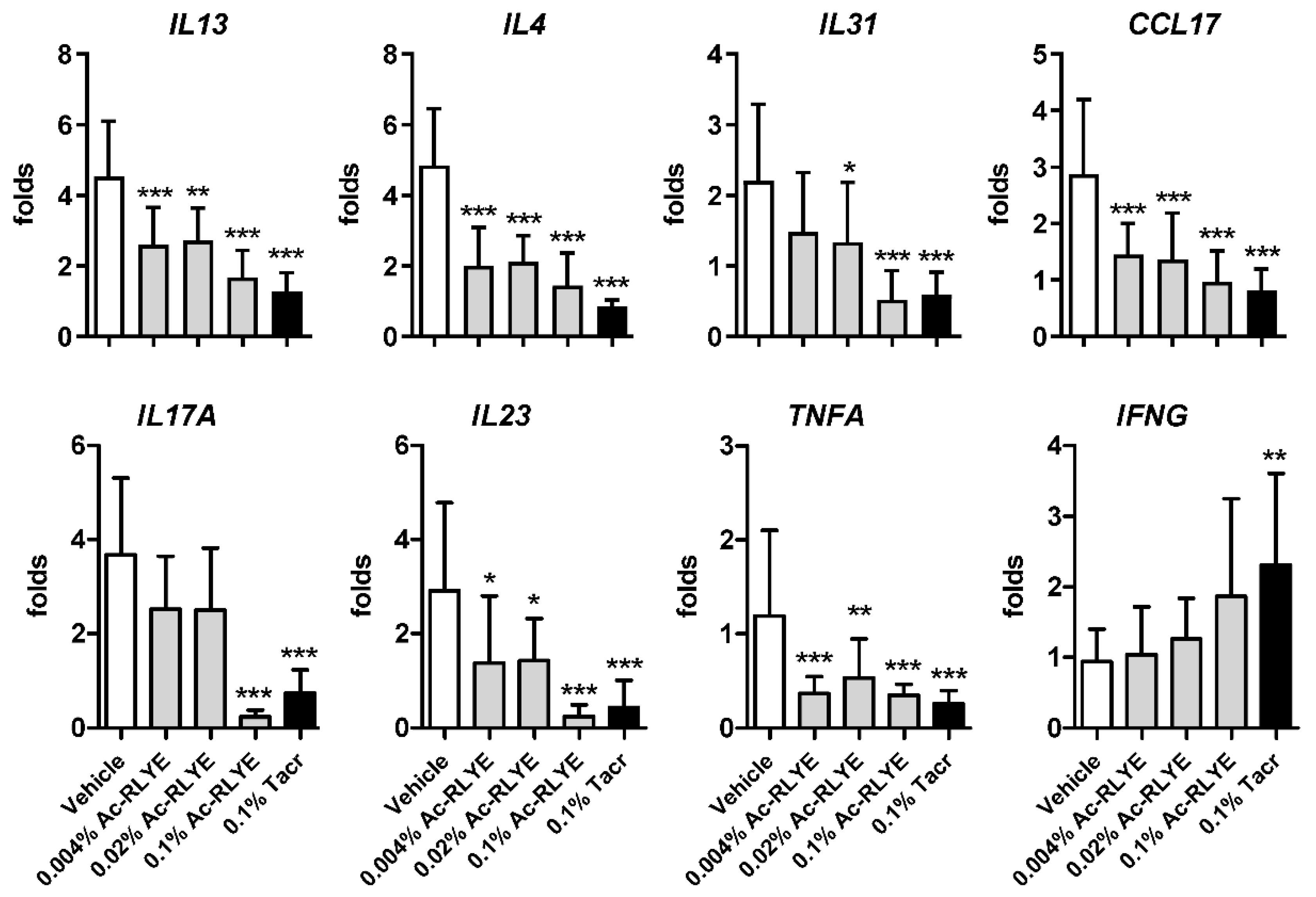
2.6. Ac-RLYE Suppressed HDM-Induced Th1 and Th2 Cytokine Expression
3. Discussion
4. Reagents and Materials
4.1. Reagents
4.2. Cells
4.3. Immunofluorescence Staining of HUVECs
4.4. Animals and Ethics Approval
4.5. Induction of AD-like Skin Lesions and Topical Application
4.6. Measurement of Dermatitis Severity and Ear Thickness
4.7. Histological Analysis
4.8. Analysis of Mouse Blood
4.9. Measurement of Skin Barrier Protein Filaggrin (FLG) and Plasma IgE
4.10. Real-Time Quantitative RT-PCR
4.11. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barbarot, S.; Auziere, S.; Gadkari, A.; Girolomoni, G.; Puig, L.; Simpson, E.L.; Margolis, D.J.; de Bruin-Weller, M.; Eckert, L. Epidemiology of atopic dermatitis in adults: Results from an international survey. Allergy 2018, 73, 1284–1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flohr, C.; Mann, J. New insights into the epidemiology of childhood atopic dermatitis. Allergy 2014, 69, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Galli, E.; Cinicola, B.; Carello, R.; Caimmi, S.; Brindisi, G.; De Castro, G.; Zicari, A.M.; Tosca, M.A.; Manti, S.; Martelli, A.; et al. Atopic dermatitis. Acta Biomed. 2020, 91, e2020011. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.W.; Le Couter, J.; Strauss, E.C.; Ferrara, N. Vascular endothelial growth factor a in intraocular vascular disease. Ophthalmology 2013, 120, 106–114. [Google Scholar] [CrossRef]
- Varricchi, G.; Granata, F.; Loffredo, S.; Genovese, A.; Marone, G. Angiogenesis and lymphangiogenesis in inflammatory skin disorders. J. Am. Acad. Dermatol. 2015, 73, 144–153. [Google Scholar] [CrossRef]
- Ferrara, N.; Gerber, H.P.; LeCouter, J. The biology of VEGF and its receptors. Nat. Med. 2003, 9, 669–676. [Google Scholar] [CrossRef]
- Sawano, A.; Iwai, S.; Sakurai, Y.; Ito, M.; Shitara, K.; Nakahata, T.; Shibuya, M. Flt-1, vascular endothelial growth factor receptor 1, is a novel cell surface marker for the lineage of monocyte-macrophages in humans. Blood 2001, 97, 785–791. [Google Scholar] [CrossRef] [Green Version]
- Kim, I.; Moon, S.O.; Kim, S.H.; Kim, H.J.; Koh, Y.S.; Koh, G.Y. Vascular endothelial growth factor expression of intercellular adhesion molecule 1 (ICAM-1), vascular cell adhesion molecule 1 (VCAM-1), and E-selectin through nuclear factor-kappa B activation in endothelial cells. J. Biol. Chem. 2001, 276, 7614–7620. [Google Scholar] [CrossRef] [Green Version]
- Baek, Y.Y.; Lee, D.K.; Kim, J.; Kim, J.H.; Park, W.; Kim, T.; Han, S.; Jeoung, D.; You, J.C.; Lee, H.; et al. Arg-Leu-Tyr-Glu tetrapeptide inhibits tumor progression by suppressing angiogenesis and vascular permeability via VEGF receptor-2 antagonism. Oncotarget 2017, 8, 11763–11777. [Google Scholar] [CrossRef] [Green Version]
- Baek, Y.Y.; Lee, D.K.; So, J.H.; Kim, C.H.; Jeoung, D.; Lee, H.; Choe, J.; Won, M.H.; Ha, K.S.; Kwon, Y.G.; et al. The tetrapeptide Arg-Leu-Tyr-Glu inhibits VEGF-induced angiogenesis. Biochem. Biophys. Res. Commun. 2015, 463, 532–537. [Google Scholar] [CrossRef]
- Yun, J.A.; Kim, J.; Baek, Y.Y.; Park, W.; Park, M.; Kim, S.; Kim, T.; Choi, S.; Jeoung, D.; Lee, H.; et al. N-Terminal Modification of the Tetrapeptide Arg-Leu-Tyr-Glu, a Vascular Endothelial Growth Factor Receptor-2 (VEGFR-2) Antagonist, Improves Antitumor Activity by Increasing its Stability against Serum Peptidases. Mol. Pharmacol. 2019, 96, 692–701. [Google Scholar] [CrossRef] [Green Version]
- Koo, H.C.; Baek, Y.Y.; Choi, J.S.; Kim, Y.M.; Sung, B.; Kim, M.J.; Kim, J.G.; You, J.C. Therapeutic Efficacy of a Novel Acetylated Tetrapeptide in Animal Models of Age-Related Macular Degeneration. Int. J. Mol. Sci. 2021, 22, 3893. [Google Scholar] [CrossRef]
- Park, W.; Baek, Y.Y.; Kim, J.; Jo, D.H.; Choi, S.; Kim, J.H.; Kim, T.; Kim, S.; Park, M.; Kim, J.Y.; et al. Arg-Leu-Tyr-Glu Suppresses Retinal Endothelial Permeability and Choroidal Neovascularization by Inhibiting the VEGF Receptor 2 Signaling Pathway. Biomol. Ther. 2019, 27, 474–483. [Google Scholar] [CrossRef]
- Koczy-Baron, E.; Jochem, J.; Kasperska-Zajac, A. Increased plasma concentration of vascular endothelial growth factor in patients with atopic dermatitis and its relation to disease severity and platelet activation. Inflamm. Res. 2012, 61, 1405–1409. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Matsuo, H.; Morita, E. Increased production of vascular endothelial growth factor in the lesions of atopic dermatitis. Arch. Dermatol. Res. 2006, 297, 425–429. [Google Scholar] [CrossRef]
- Amarbayasgalan, T.; Takahashi, H.; Dekio, I.; Morita, E. Content of vascular endothelial growth factor in stratum corneum well correlates to local severity of acute inflammation in patients with atopic dermatitis. Int. Arch. Allergy Immunol. 2012, 157, 251–258. [Google Scholar] [CrossRef]
- Heo, M.J.; Lee, C.; Choi, S.Y.; Choi, Y.M.; An, I.S.; Bae, S.; An, S.; Jung, J.H. Nintedanib ameliorates animal model of dermatitis. Sci. Rep. 2020, 10, 4493. [Google Scholar] [CrossRef] [Green Version]
- Warbrick, E.V.; Dearman, R.J.; Basketter, D.A.; Kimber, I. Analysis of interleukin 12 protein production and mRNA expression in mice exposed topically to chemical allergens. Toxicology 1999, 132, 57–66. [Google Scholar] [CrossRef]
- Lubach, D.; Bensmann, A.; Bornemann, U. Steroid-induced dermal atrophy. Investigations on discontinuous application. Dermatologica 1989, 179, 67–72. [Google Scholar] [CrossRef]
- Reitamo, S.; Rissanen, J.; Remitz, A.; Granlund, H.; Erkko, P.; Elg, P.; Autio, P.; Lauerma, A.I. Tacrolimus ointment does not affect collagen synthesis: Results of a single-center randomized trial. J. Investig. Dermatol. 1998, 111, 396–398. [Google Scholar] [CrossRef]
- Damsgaard, T.E.; Olesen, A.B.; Sorensen, F.B.; Thestrup-Pedersen, K.; Schiotz, P.O. Mast cells and atopic dermatitis. Stereological quantification of mast cells in atopic dermatitis and normal human skin. Arch. Dermatol. Res. 1997, 289, 256–260. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.; Igawa, K.; Handa, Y.; Munetsugu, T.; Satoh, T.; Yokozeki, H. Basophils and mast cells are crucial for reactions due to epicutaneous sensitization to ovalbumin. Exp. Dermatol. 2017, 26, 778–784. [Google Scholar] [CrossRef] [PubMed]
- Furue, M.; Chiba, T.; Tsuji, G.; Ulzii, D.; Kido-Nakahara, M.; Nakahara, T.; Kadono, T. Atopic dermatitis: Immune deviation, barrier dysfunction, IgE autoreactivity and new therapies. Allergol. Int. 2017, 66, 398–403. [Google Scholar] [CrossRef] [PubMed]
- Brandt, E.B.; Sivaprasad, U. Th2 Cytokines and Atopic Dermatitis. J. Clin. Cell Immunol. 2011, 2, 110. [Google Scholar] [CrossRef] [PubMed]
- Sugaya, M. The Role of Th17-Related Cytokines in Atopic Dermatitis. Int. J. Mol. Sci. 2020, 21, 1314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brockow, K.; Akin, C.; Huber, M.; Scott, L.M.; Schwartz, L.B.; Metcalfe, D.D. Levels of mast-cell growth factors in plasma and in suction skin blister fluid in adults with mastocytosis: Correlation with dermal mast-cell numbers and mast-cell tryptase. J. Allergy Clin. Immunol. 2002, 109, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Agha-Majzoub, R.; Becker, R.P.; Schraufnagel, D.E.; Chan, L.S. Angiogenesis: The major abnormality of the keratin-14 IL-4 transgenic mouse model of atopic dermatitis. Microcirculation 2005, 12, 455–476. [Google Scholar] [CrossRef]
- Shi, V.Y.; Bao, L.; Chan, L.S. Inflammation-driven dermal lymphangiogenesis in atopic dermatitis is associated with CD11b+ macrophage recruitment and VEGF-C up-regulation in the IL-4-transgenic mouse model. Microcirculation 2012, 19, 567–579. [Google Scholar] [CrossRef]
- Van Aanhold, C.C.L.; Bus, P.; Zandbergen, M.; Bos, M.; Berbee, J.F.P.; Quint, K.D.; Bruijn, J.A.; Baelde, H.J. The Vascular Endothelial Growth Factor Inhibitor Soluble FLT-1 Ameliorates Atopic Dermatitis in APOC1 Transgenic Mice. J. Investig. Dermatol. 2020, 140, 491–494.e494. [Google Scholar] [CrossRef]
- Halin, C.; Fahrngruber, H.; Meingassner, J.G.; Bold, G.; Littlewood-Evans, A.; Stuetz, A.; Detmar, M. Inhibition of chronic and acute skin inflammation by treatment with a vascular endothelial growth factor receptor tyrosine kinase inhibitor. Am. J. Pathol. 2008, 173, 265–277. [Google Scholar] [CrossRef]
- Tatari, N.; Movassagh, H.; Shan, L.; Koussih, L.; Gounni, A.S. Semaphorin 3E Inhibits House Dust Mite-Induced Angiogenesis in a Mouse Model of Allergic Asthma. Am. J. Pathol. 2019, 189, 762–772. [Google Scholar] [CrossRef]
- Lampugnani, M.G. Endothelial cell-to-cell junctions: Adhesion and signaling in physiology and pathology. Cold Spring Harb. Perspect. Med. 2012, 2, a006528. [Google Scholar] [CrossRef] [Green Version]
- Claesson-Welsh, L.; Dejana, E.; McDonald, D.M. Permeability of the Endothelial Barrier: Identifying and Reconciling Controversies. Trends Mol. Med. 2021, 27, 314–331. [Google Scholar] [CrossRef]
- Tsakok, T.; Woolf, R.; Smith, C.H.; Weidinger, S.; Flohr, C. Atopic dermatitis: The skin barrier and beyond. Br. J. Dermatol. 2019, 180, 464–474. [Google Scholar] [CrossRef]
- Jungersted, J.M.; Scheer, H.; Mempel, M.; Baurecht, H.; Cifuentes, L.; Hogh, J.K.; Hellgren, L.I.; Jemec, G.B.; Agner, T.; Weidinger, S. Stratum corneum lipids, skin barrier function and filaggrin mutations in patients with atopic eczema. Allergy 2010, 65, 911–918. [Google Scholar] [CrossRef]
- Palmer, C.N.; Irvine, A.D.; Terron-Kwiatkowski, A.; Zhao, Y.; Liao, H.; Lee, S.P.; Goudie, D.R.; Sandilands, A.; Campbell, L.E.; Smith, F.J.; et al. Common loss-of-function variants of.f the epidermal barrier protein filaggrin are a major predisposing factor for atopic dermatitis. Nat. Genet. 2006, 38, 441–446. [Google Scholar] [CrossRef]
- Esparza-Gordillo, J.; Matanovic, A.; Marenholz, I.; Bauerfeind, A.; Rohde, K.; Nemat, K.; Lee-Kirsch, M.A.; Nordenskjold, M.; Winge, M.C.; Keil, T.; et al. Maternal filaggrin mutations increase the risk of atopic dermatitis in children: An effect independent of mutation inheritance. PLoS Genet. 2015, 11, e1005076. [Google Scholar] [CrossRef] [Green Version]
- Howell, M.D.; Kim, B.E.; Gao, P.; Grant, A.V.; Boguniewicz, M.; Debenedetto, A.; Schneider, L.; Beck, L.A.; Barnes, K.C.; Leung, D.Y. Cytokine modulation of atopic dermatitis filaggrin skin expression. J. Allergy Clin. Immunol. 2007, 120, 150–155. [Google Scholar] [CrossRef] [Green Version]
- Gutowska-Owsiak, D.; Schaupp, A.L.; Salimi, M.; Selvakumar, T.A.; McPherson, T.; Taylor, S.; Ogg, G.S. IL-17 downregulates filaggrin and affects keratinocyte expression of genes associated with cellular adhesion. Exp. Dermatol. 2012, 21, 104–110. [Google Scholar] [CrossRef]
- Meng, J.; Li, Y.; Fischer, M.J.M.; Steinhoff, M.; Chen, W.; Wang, J. Th2 Modulation of Transient Receptor Potential Channels: An Unmet Therapeutic Intervention for Atopic Dermatitis. Front. Immunol. 2021, 12, 696784. [Google Scholar] [CrossRef]
- Van de Veen, W.; Akdis, M. The use of biologics for immune modulation in allergic disease. J. Clin. Investig. 2019, 129, 1452–1462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gandhi, N.A.; Pirozzi, G.; Graham, N.M.H. Commonality of the IL-4/IL-13 pathway in atopic diseases. Expert Rev. Clin. Immunol. 2017, 13, 425–437. [Google Scholar] [CrossRef] [PubMed]
- Albanesi, C.; Scarponi, C.; Sebastiani, S.; Cavani, A.; Federici, M.; De Pita, O.; Puddu, P.; Girolomoni, G. IL-4 enhances keratinocyte expression of CXCR3 agonistic chemokines. J. Immunol. 2000, 165, 1395–1402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koga, C.; Kabashima, K.; Shiraishi, N.; Kobayashi, M.; Tokura, Y. Possible pathogenic role of Th17 cells for atopic dermatitis. J. Investig. Dermatol. 2008, 128, 2625–2630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nograles, K.E.; Zaba, L.C.; Shemer, A.; Fuentes-Duculan, J.; Cardinale, I.; Kikuchi, T.; Ramon, M.; Bergman, R.; Krueger, J.G.; Guttman-Yassky, E. IL-22-producing “T22” T cells account for upregulated IL-22 in atopic dermatitis despite reduced IL-17-producing TH17 T cells. J. Allergy Clin. Immunol. 2009, 123, 1244–1252.e1242. [Google Scholar] [CrossRef] [Green Version]
- Nakajima, S.; Kitoh, A.; Egawa, G.; Natsuaki, Y.; Nakamizo, S.; Moniaga, C.S.; Otsuka, A.; Honda, T.; Hanakawa, S.; Amano, W.; et al. IL-17A as an inducer for Th2 immune responses in murine atopic dermatitis models. J. Investig. Dermatol. 2014, 134, 2122–2130. [Google Scholar] [CrossRef] [Green Version]
- Wakita, H.; Sakamoto, T.; Tokura, Y.; Takigawa, M. E-selectin and vascular cell adhesion molecule-1 as critical adhesion molecules for infiltration of T lymphocytes and eosinophils in atopic dermatitis. J. Cutan Pathol. 1994, 21, 33–39. [Google Scholar] [CrossRef]
- Bhushan, M.; Craven, N.M.; Beck, M.H.; Chalmers, R.J. Linear porokeratosis of mibelli: Successful treatment with cryotherapy. Br. J. Dermatol. 1999, 141, 389. [Google Scholar] [CrossRef]
- Matsuda, H.; Watanabe, N.; Geba, G.P.; Sperl, J.; Tsudzuki, M.; Hiroi, J.; Matsumoto, M.; Ushio, H.; Saito, S.; Askenase, P.W.; et al. Development of atopic dermatitis-like skin lesion with IgE hyperproduction in NC/Nga mice. Int. Immunol. 1997, 9, 461–466. [Google Scholar] [CrossRef]
- Leung, D.Y.; Hirsch, R.L.; Schneider, L.; Moody, C.; Takaoka, R.; Li, S.H.; Meyerson, L.A.; Mariam, S.G.; Goldstein, G.; Hanifin, J.M. Thymopentin therapy reduces the clinical severity of atopic dermatitis. J. Allergy Clin. Immunol. 1990, 85, 927–933. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sung, B.; Baek, Y.-Y.; Kim, Y.-M.; You, J.C. Topical Administration of a Novel Acetylated Tetrapeptide Suppresses Vascular Permeability and Immune Responses and Alleviates Atopic Dermatitis in a Murine Model. Int. J. Mol. Sci. 2022, 23, 13498. https://doi.org/10.3390/ijms232113498
Sung B, Baek Y-Y, Kim Y-M, You JC. Topical Administration of a Novel Acetylated Tetrapeptide Suppresses Vascular Permeability and Immune Responses and Alleviates Atopic Dermatitis in a Murine Model. International Journal of Molecular Sciences. 2022; 23(21):13498. https://doi.org/10.3390/ijms232113498
Chicago/Turabian StyleSung, Bokyung, Yi-Yong Baek, Young-Myeong Kim, and Ji Chang You. 2022. "Topical Administration of a Novel Acetylated Tetrapeptide Suppresses Vascular Permeability and Immune Responses and Alleviates Atopic Dermatitis in a Murine Model" International Journal of Molecular Sciences 23, no. 21: 13498. https://doi.org/10.3390/ijms232113498
APA StyleSung, B., Baek, Y. -Y., Kim, Y. -M., & You, J. C. (2022). Topical Administration of a Novel Acetylated Tetrapeptide Suppresses Vascular Permeability and Immune Responses and Alleviates Atopic Dermatitis in a Murine Model. International Journal of Molecular Sciences, 23(21), 13498. https://doi.org/10.3390/ijms232113498