
Altered Expression of Genes Associated with Major Neurotransmitter Systems in the Reward-Related Brain Regions of Mice with Positive Fighting Experience

Abstract
:1. Introduction
2. Results
2.1. Agonistic Behavior
2.2. Neurotranscriptomic Data
2.2.1. DEGs in the VTA of Highly Aggressive Mice
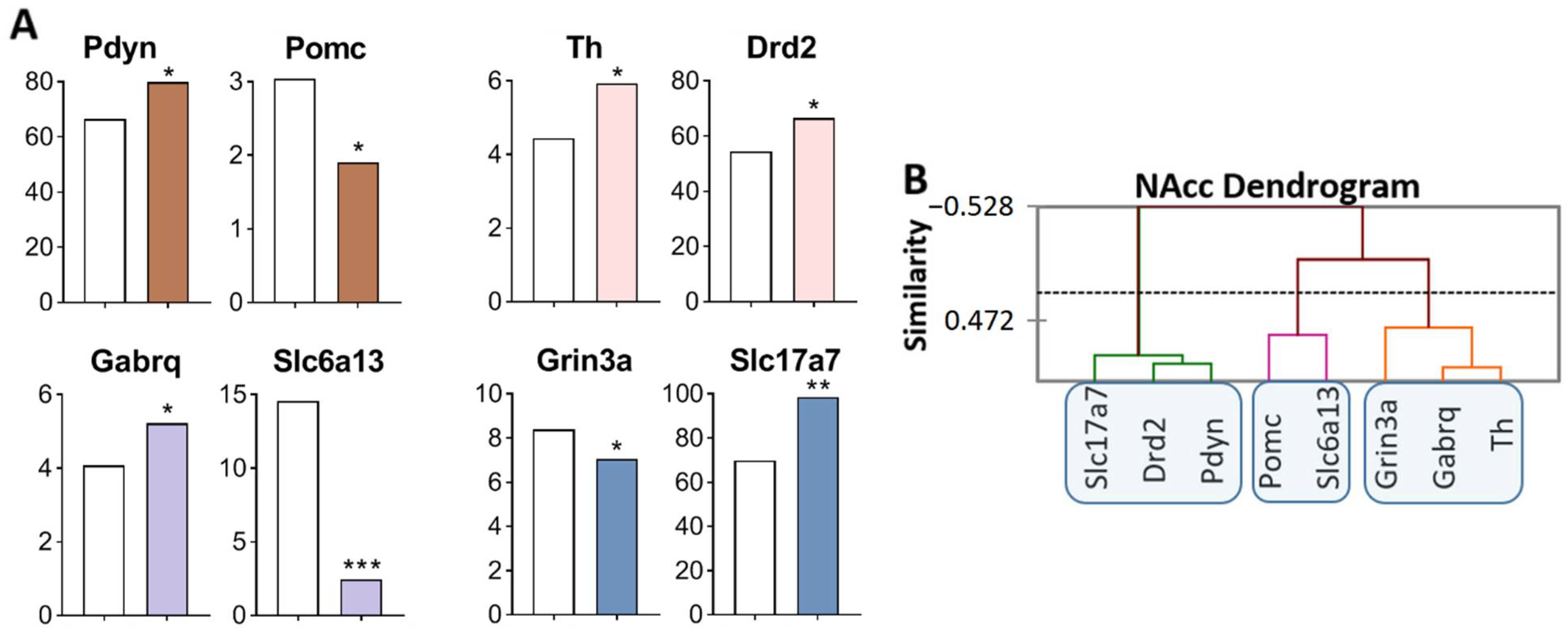
2.2.2. DEGs in the NAcc of Highly Aggressive Mice
2.2.3. DEGs in the STR of Highly Aggressive Mice
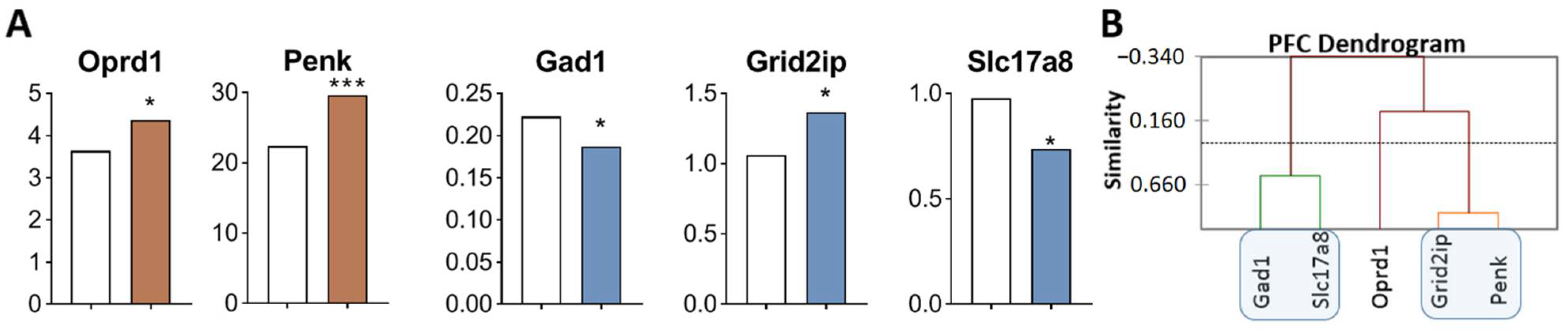
2.2.4. DEGs in the PFC of Highly Aggressive Mice
2.2.5. DEGs in the MRNs of Highly Aggressive Mice
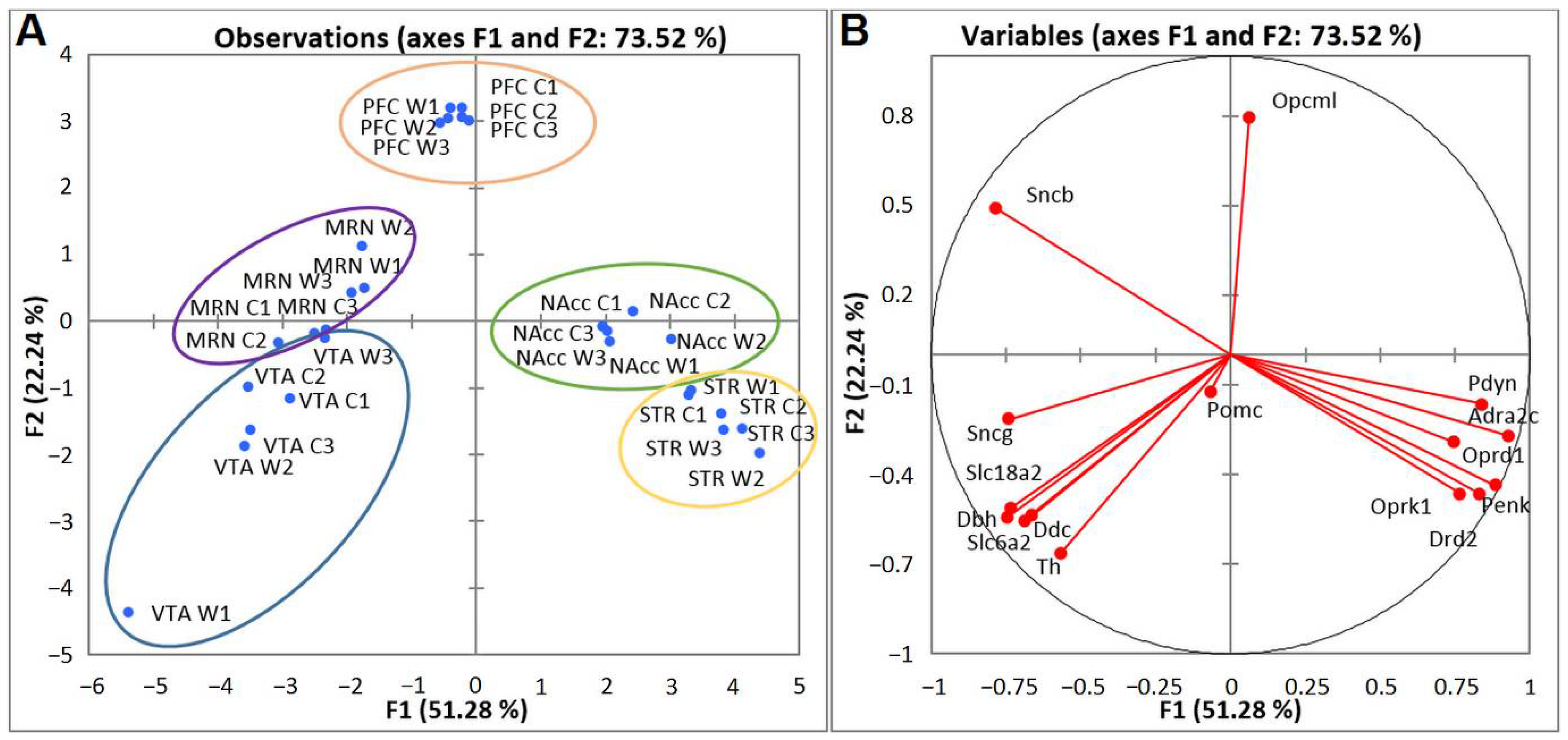
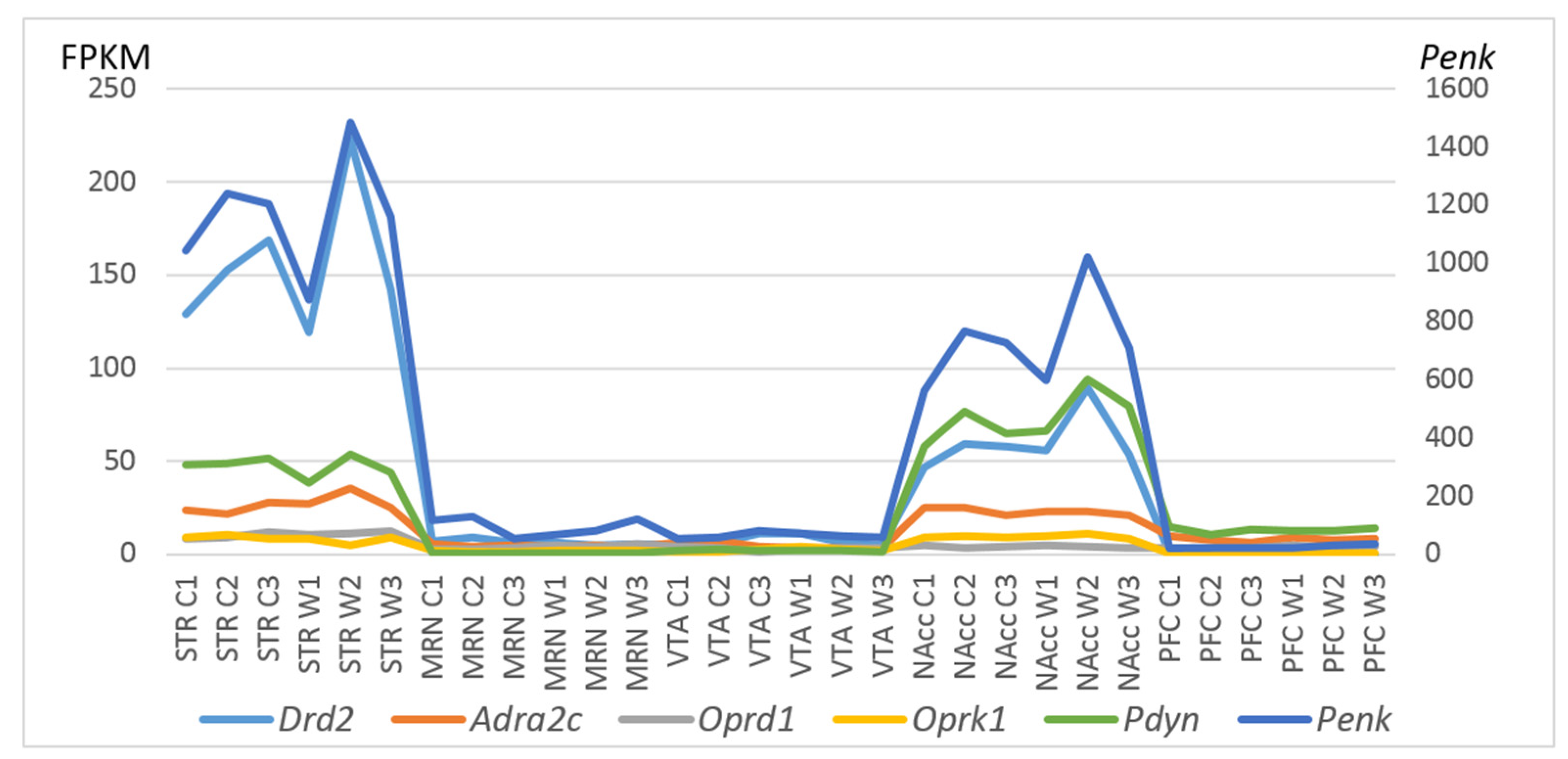
2.3. PCA Based on CAergic and Opioidergic DEGs’ Expression Profiles in Brain Regions of Highly Aggressive Mice
3. Discussion
4. Methods and Materials
4.1. Animals
4.2. The Sensory Contact Model for the Development of Pathological Aggression in Mice
4.3. Analysis of the Winners’ Behavior during Agonistic Confrontation
4.4. High-Throughput RNA Sequencing (RNA-Seq)
4.5. Brain Regions That Are Responsible for the Regulation of Aggression and Development of Addiction during Reward
4.6. The Genes That Were Analyzed in Different Brain Regions
4.7. Statistical Analysis
5. Conclusions
6. Limitations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 5-HT | serotonin |
| AHC | agglomerative hierarchical clustering |
| C | control |
| W | winners |
| CAergic | catecholaminergic |
| FPKM | fragments per kilobase of transcript per million mapped reads |
| GABA | γ-aminobutyric acid |
| MRNs | midbrain raphe nuclei |
| MSNs | medium spiny neurons |
| NAcc | nucleus accumbens |
| PCA | principal component analysis |
| PFC | prefrontal cortex |
| STR | dorsal striatum |
| VTA | ventral tegmental area |
References
- Miczek, K.A.; Faccidomo, S.P.; Fish, E.W.; DeBold, J.F. Neurochemistry and molecular neurobiology of aggressive behavior. In Handbook of Neurochemistry and Molecular Neurobiology: Behavioral Neurochemistry, Neuroendocrinology and Molecular Neurobiology; Lajtha, A., Blaustein, J.D., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 285–336. [Google Scholar]
- De Almeida, R.M.; Ferrari, P.F.; Parmigiani, S.; Miczek, K.A. Escalated aggressive behavior: Dopamine, serotonin and GABA. Eur. J. Pharmacol. 2005, 526, 51–64. [Google Scholar] [CrossRef] [PubMed]
- Narvaes, R.; de Almeida, R.M.M. Aggressive behavior and three neurotransmitters: Dopamine, GABA, and serotonin—A review of the last 10 years. Psychol. Neurosci. 2014, 7, 601–607. [Google Scholar] [CrossRef]
- Takahashi, A.; Lee, R.X.; Iwasato, T.; Itohara, S.; Arima, H.; Bettler, B.; Miczek, K.A.; Koide, T. Glutamate input in the dorsal raphe nucleus as a determinant of escalated aggression in male mice. J. Neurosci. 2015, 35, 6452–6463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haller, J. Studies into abnormal aggression in humans and rodents: Methodological and translational aspects. Neurosci. Biobehav. Rev. 2017, 76, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Aleyasin, H.; Flanigan, M.E.; Russo, S.J. Neurocircuitry of aggression and aggression seeking behavior: Nose poking into brain circuitry controlling aggression. Curr. Opin. Neurobiol. 2018, 49, 184–191. [Google Scholar] [CrossRef] [PubMed]
- de Boer, S.F. Animal models of excessive aggression: Implications for human aggression and violence. Curr. Opin. Psychol. 2018, 19, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Tordjman, S.; Carlier, M.; Cohen, D.; Cesselin, F.; Bourgoin, S.; Colas-Linhart, N.; Petiet, A.; Perez-Diaz, F.; Hamon, M.; Roubertoux, P.L. Aggression and the three opioid families (endorphins, enkephalins, and dynorphins) in mice. Behav. Genet. 2003, 33, 529–536. [Google Scholar] [CrossRef]
- Benton, D.; Brain, P.F. The role of opioid mechanisms in social interaction and attachment. In Endorphins, Opiates and Behavioural Processes; Rodgers, R.J., Cooper, S.J., Eds.; John Wiley and Sons Ltd: London, UK, 1988; pp. 217–235. [Google Scholar]
- De Boer, S.F.; Buwalda, B.; Koolhaas, J.M. Untangling the neurobiology of coping styles in rodents: Towards neural mechanisms underlying individual differences in disease susceptibility. Neurosci. Biobehav. Rev. 2017, 74, 401–422. [Google Scholar] [CrossRef]
- Glick, A.R. The role of serotonin in impulsive aggression, suicide, and homicide in adolescents and adults: A literature review. Int J. Adolesc. Med. Health 2015, 27, 143–150. [Google Scholar] [CrossRef]
- Kulikov, A.V.; Osipova, D.V.; Naumenko, V.S.; Terenina, E.; Mormede, P.; Popova, N.K. A pharmacological evidence of positive association between mouse intermale aggression and brain serotonin metabolism. Behav. Brain Res. 2012, 233, 113–119. [Google Scholar] [CrossRef]
- Montoya, E.R.; Terburg, D.; Bos, P.A.; van Honk, J. Testosterone, cortisol, and serotonin as key regulators of social aggression: A review and theoretical perspective. Motiv. Emot. 2012, 36, 65–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popova, N.K.; Naumenko, E.V.; Kolpakov, B.G. Serotonin and behavior. Novosib. Nauka SB RAS 1978, 4, 12. [Google Scholar]
- Pavlov, K.A.; Chistiakov, D.A.; Chekhonin, V.P. Genetic determinants of aggression and impulsivity in humans. J. Appl. Genet. 2012, 53, 61–82. [Google Scholar] [CrossRef]
- Takahashi, A.; Quadros, I.M.; de Almeida, R.M.; Miczek, K.A. Brain serotonin receptors and transporters: Initiation vs. termination of escalated aggression. Psychopharmacology 2011, 213, 183–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kudriavtseva, N.N.; Bakshtanovskaia, I.V. The neurochemical control of aggression and submission. Zh. Vyssh. Nerv. Deiat. Im. I P Pavlova 1991, 41, 459–466. [Google Scholar] [PubMed]
- Kudryavtseva, N.N.; Lipina, T.V.; Koryakina, L.A. Effects of haloperidol on communicative and aggressive behavior in male mice with different experiences of aggression. Pharmacol. Biochem. Behav. 1999, 63, 229–236. [Google Scholar] [CrossRef]
- Bondar, N.P.; Kudryavtseva, N.N. The effects of the D1 receptor antagonist SCH-23390 on individual and aggressive behavior in male mice with different experience of aggression. Neurosci. Behav. Physiol. 2005, 35, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Lipina, T.V.; Avgustinovich, D.F.; Koriakina, L.A.; Alekseenko, O.V.; Kudriavtseva, N.N. Differences in the effects of naltrexone on the communicative and aggressive behavi.iors of subjects with different experiences of social conquests. Eksp. Klin. Farmakol. 1998, 61, 13–18. [Google Scholar]
- Kudriavtseva, N.N.; Dolgov, V.V.; Avgustinovich, D.F.; Alekseenko, O.V.; Lipina, T.V.; Koriakina, L.A. Modifying effect of the repeated experience of agonistic confrontations on effect of naltrexone in male mice. Ross. Fiziol. Zh. Im. I. M. Sechenova 2001, 87, 227–238. [Google Scholar]
- Bondar, N.P.; Smagin, D.A.; Kudryavtseva, N.N. Effects of single and chronic naltrexone treatment on agonistic behavior of male mice with repeated experience of aggression. Psychopharmacol. Biol. Narcol. 2011, 11, 2688–2700. [Google Scholar]
- Kudryavtseva, N.N. Straub tail, the deprivation effect and addiction to aggression. In Motivation of Health Behavior; O’Neal, P.W., Ed.; NOVA Science Publishers: New York, NY, USA, 2007; pp. 97–110. [Google Scholar]
- Kudryavtseva, N.N.; Gerrits, M.A.; Avgustinovich, D.F.; Tenditnik, M.V.; van Ree, J.M. Modulation of anxiety-related behaviors by mu- and kappa-opioid receptor agonists depends on the social status of mice. Peptides 2004, 25, 1355–1363. [Google Scholar] [CrossRef] [PubMed]
- Filipenko, M.L.; Alekseyenko, O.V.; Beilina, A.G.; Kamynina, T.P.; Kudryavtseva, N.N. Increase of tyrosine hydroxylase and dopamine transporter mRNA levels in ventral tegmental area of male mice under influence of repeated aggression experience. Brain Res. Mol. Brain Res. 2001, 96, 77–81. [Google Scholar] [CrossRef]
- Bondar, N.P.; Boyarskikh, U.A.; Kovalenko, I.L.; Filipenko, M.L.; Kudryavtseva, N.N. Molecular implications of repeated aggression: Th, Dat1, Snca and Bdnf gene expression in the VTA of victorious male mice. PLoS ONE 2009, 4, e4190. [Google Scholar] [CrossRef]
- Filipenko, M.L.; Beylina, A.G.; Alekseyenko, O.V.; Timofeeva, O.A.; Avgustinovich, D.F.; Kudryavtseva, N.N. Association between brain COMT gene expression and aggressive experience in daily agonistic confrontations in male mice. In Stress: Neural, Endocrine and Molecular Studies; McCarty, R., Aguilera, G., Sabban, E., Kvetnyansky, R., Eds.; Taylor & Francis: New York, NY, USA; London, UK, 2002; pp. 157–161. [Google Scholar]
- Kudryavtseva, N.N. Psychopathology of repeated aggression: A neurobiological aspect. In Perspectives on the Psychology of Aggression; Morgan, J.P., Ed.; NOVA Science Publishers, Inc.: New York, NY, USA, 2006; pp. 35–64. [Google Scholar]
- Kudryavtseva, N.N. Positive fighting experience, addiction-like state, and relapse: Retrospective analysis of experimental studies. Aggress. Viol. Behav. 2020, 52, 101403. [Google Scholar] [CrossRef]
- Kudryavtseva, N.N.; Kovalenko, I.L.; Smagin, D.A.; Galyamina, A.G.; Babenko, V.N. Abnormal social behaviors and dysfunction of autism-related genes associated with daily agonistic interactions in mice. In Molecular-Genetic and Statistical Techniques for Behavioral and Neural Research; Gerlai, R.T., Ed.; Academic Press: San Diego, CA, USA, 2018; pp. 309–344. [Google Scholar]
- Kudryavtseva, N.N.; Smagin, D.A.; Bondar, N.P. Modeling fighting deprivation effect in mouse repeated aggression paradigm. Prog. Neuropsychopharmacol. Biol. Psychiatry 2011, 35, 1472–1478. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.K.; Hassanein, N.M.A.; Ahmed, H.M.S. Psychopharmacological assessment of the sensory contact model as a possible model of mania. J. Glob. Biosci. 2016, 5, 3725–3741. [Google Scholar]
- Covington, H.E., 3rd; Newman, E.L.; Leonard, M.Z.; Miczek, K.A. Translational models of adaptive and excessive fighting: An emerging role for neural circuits in pathological aggression. F1000Research 2019, 8, 963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golden, S.A.; Shaham, Y. Aggression addiction and relapse: A new frontier in psychiatry. Neuropsychopharmacology 2018, 43, 224–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golden, S.A.; Jin, M.; Heins, C.; Venniro, M.; Michaelides, M.; Shaham, Y. Nucleus accumbens Drd1-expressing neurons control aggression self-administration and aggression seeking in mice. J. Neurosci. 2019, 39, 2482–2496. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Arias, M.; Navarrete, F.; Daza-Losada, M.; Navarro, D.; Aguilar, M.A.; Berbel, P.; Minarro, J.; Manzanares, J. CB1 cannabinoid receptor-mediated aggressive behavior. Neuropharmacology 2013, 75, 172–180. [Google Scholar] [CrossRef]
- Miczek, K.A.; Takahashi, A.; Gobrogge, K.L.; Hwa, L.S.; de Almeida, R.M. Escalated aggression in animal models: Shedding new light on mesocorticolimbic circuits. Curr. Opin. Behav. Sci. 2015, 3, 90–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enoch, M.A.; Rosser, A.A.; Zhou, Z.; Mash, D.C.; Yuan, Q.; Goldman, D. Expression of glutamatergic genes in healthy humans across 16 brain regions; altered expression in the hippocampus after chronic exposure to alcohol or cocaine. Genes Brain Behav. 2014, 13, 758–768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moeller, S.J.; London, E.D.; Northoff, G. Neuroimaging markers of glutamatergic and GABAergic systems in drug addiction: Relationships to resting-state functional connectivity. Neurosci. Biobehav. Rev. 2016, 61, 35–52. [Google Scholar] [CrossRef] [Green Version]
- Cooper, S.J. Interaction between endogenous opioids and dopamine: Implications for reward and aversion. In The Mesolimbic Dopamine System: From Motivation to Action; Willner, P., Scheel-Kruger, J., Eds.; John Wiley and Sons Ltd: London, UK, 1991; pp. 331–366. [Google Scholar]
- Van Ree, J.M.; Gerrits, M.A.; Vanderschuren, L.J. Opioids, reward and addiction: An encounter of biology, psychology, and medicine. Pharmacol. Rev. 1999, 51, 341–396. [Google Scholar]
- Volkow, N.D.; Morales, M. The brain on drugs: From reward to addiction. Cell 2015, 162, 712–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kudryavtseva, N.N. The sensory contact model for the study of aggressive and submissive behaviors in male mice. Aggress. Behav. 1991, 17, 285–291. [Google Scholar] [CrossRef]
- Kudryavtseva, N.N.; Smagin, D.A.; Kovalenko, I.L.; Vishnivetskaya, G.B. Repeated positive fighting experience in male inbred mice. Nat. Protoc. 2014, 9, 2705–2717. [Google Scholar] [CrossRef]
- Redina, O.; Babenko, V.; Smagin, D.; Kovalenko, I.; Galyamina, A.; Efimov, V.; Kudryavtseva, N. Gene expression changes in the ventral tegmental area of male mice with alternative social behavior experience in chronic agonistic interactions. Int. J. Mol. Sci. 2020, 21, 6599. [Google Scholar] [CrossRef]
- Babenko, V.N.; Smagin, D.A.; Kovalenko, I.L.; Galyamina, A.G.; Kudryavtseva, N.N. Differentially expressed genes of the Slc6a family as markers of altered brain neurotransmitter system function in pathological states in mice. Neurosci. Behav. Physiol. 2020, 50, 199–209. [Google Scholar] [CrossRef]
- Babenko, V.N.; Galyamina, A.G.; Rogozin, I.B.; Smagin, D.A.; Kudryavtseva, N.N. Dopamine response gene pathways in dorsal striatum MSNs from a gene expression viewpoint: cAMP-mediated gene networks. BMC Neurosci. 2020, 21, 12. [Google Scholar] [CrossRef] [Green Version]
- Redina, O.E.; Babenko, V.N.; Smagin, D.A.; Kovalenko, I.L.; Galyamina, A.G.; Kudryavtseva, N.N. Correlation of expression changes between genes controlling 5-HT synthesis and genes Crh and Trh in the midbrain raphe nuclei of chronically aggressive and defeated male mice. Genes 2021, 12, 1811. [Google Scholar] [CrossRef] [PubMed]
- Randesi, M.; van den Brink, W.; Levran, O.; Blanken, P.; van Ree, J.M.; Ott, J.; Kreek, M.J. VMAT2 gene (SLC18A2) variants associated with a greater risk for developing opioid dependence. Pharmacogenomics 2019, 20, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Kudryavtseva, N.N.; Smagin, D.A.; Kovalenko, I.L.; Galyamina, A.G.; Vishnivetskaya, G.B.; Babenko, V.N.; Orlov, Y.L. Serotonergic genes in the development of anxiety/depression-like state and pathology of aggressive behavior in male mice: RNA-seq data. Mol. Biol. 2017, 51, 288–300. [Google Scholar] [CrossRef]
- Lagerspetz, K.M.J. Studies on the aggressive behavior of mice. Ann. Acad. Sci. Fenn. B 1964, 131, 1–131. [Google Scholar]
- Scott, J.P. Agonistic behavior of mice and rats: A review. Am. Zool. 1966, 6, 683–701. [Google Scholar] [CrossRef] [Green Version]
- Scott, J.P. Theoretical issues concerning the origin and causes of fighting. In The Physiology of Aggression and Defeat; Eleftheriou, B.E., Scott, J.P., Eds.; Plenum New-York: New York, NY, USA, 1971; pp. 11–42. [Google Scholar]
- Brain, P.F.; Kamal, K.B.H. Effects of prior social experience on individual aggressiveness in laboratory rodents. Rassegna Psicol. 1989, 6, 37–43. [Google Scholar]
- Ramirez, J.M. Aggression: Causes and functions. Hiroshima For. Psychol. 1996, 17, 21–37. [Google Scholar]
- Fish, E.W.; de Bold, J.F.; Miczek, K.A. Aggressive behavior as a reinforcer in mice: Activation by allopregnanolone. Psychopharmacology 2002, 163, 459–466. [Google Scholar] [CrossRef]
- Kudryavtseva, N.N. Lorenz was right! Or does aggressive energy accumulate? Russ. J. Genet. 2004, 40, 656–662. [Google Scholar] [CrossRef]
- Robinson, T.E.; Berridge, K.C. Addiction. Ann. Rev. Psychol. 2003, 54, 25–53. [Google Scholar] [CrossRef]
- Smagin, D.A.; Boyarskikh, U.A.; Bondar, N.P.; Filipenko, M.L.; Kudryavtseva, N.N. Reduction of serotonergic gene expression in the midbrain raphe nuclei under positive fighting experience. Adv. Biosci. Biotechnol. 2013, 4, 36–44. [Google Scholar] [CrossRef] [Green Version]
- Flatscher-Bader, T.; Zuvela, N.; Landis, N.; Wilce, P.A. Smoking and alcoholism target genes associated with plasticity and glutamate transmission in the human ventral tegmental area. Hum. Mol. Genet. 2008, 17, 38–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farris, S.P.; Harris, R.A.; Ponomarev, I. Epigenetic modulation of brain gene networks for cocaine and alcohol abuse. Front. Neurosci. 2015, 9, 176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, B.L.; Oscar-Berman, M.; Kaplan, G.B. Opioid-induced structural and functional plasticity of medium-spiny neurons in the nucleus accumbens. Neurosci. Biobehav. Rev. 2021, 120, 417–430. [Google Scholar] [CrossRef]
- Wang, C.W.; Ma, M.; Lu, W.G.; Luo, R.Q. Association between prodynorphin gene polymorphisms and opioid dependence susceptibility: A meta-analysis. BMC Psychiatry 2019, 19, 281. [Google Scholar] [CrossRef] [Green Version]
- Bazov, I.; Sarkisyan, D.; Kononenko, O.; Watanabe, H.; Yakovleva, T.; Hansson, A.C.; Sommer, W.H.; Spanagel, R.; Bakalkin, G. Dynorphin and kappa-opioid receptor dysregulation in the dopaminergic reward system of human alcoholics. Mol. Neurobiol. 2018, 55, 7049–7061. [Google Scholar] [CrossRef] [Green Version]
- Yuanyuan, J.; Rui, S.; Hua, T.; Jingjing, C.; Cuola, D.; Yuhui, S.; Shuguang, W. Genetic association analyses and meta-analysis of dynorphin-kappa opioid system potential functional variants with heroin dependence. Neurosci. Lett. 2018, 685, 75–82. [Google Scholar] [CrossRef]
- Hikida, T.; Yao, S.; Macpherson, T.; Fukakusa, A.; Morita, M.; Kimura, H.; Hirai, K.; Ando, T.; Toyoshiba, H.; Sawa, A. Nucleus accumbens pathways control cell-specific gene expression in the medial prefrontal cortex. Sci. Rep. 2020, 10, 1838. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, J.A.; Emory, E. Executive function and the frontal lobes: A meta-analytic review. Neuropsychol. Rev. 2006, 16, 17–42. [Google Scholar] [CrossRef]
- Steketee, J.D. Neurotransmitter systems of the medial prefrontal cortex: Potential role in sensitization to psychostimulants. Brain Res. Brain Res. Rev. 2003, 41, 203–228. [Google Scholar] [CrossRef]
- Del Arco, A.; Mora, F. Neurotransmitters and prefrontal cortex-limbic system interactions: Implications for plasticity and psychiatric disorders. J. Neural. Transm. 2009, 116, 941–952. [Google Scholar] [CrossRef] [PubMed]
- Morales, M.; Margolis, E.B. Ventral tegmental area: Cellular heterogeneity, connectivity and behaviour. Nat. Rev. Neurosci. 2017, 18, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Sheen, W.; Morales, M. Glutamatergic neurons are present in the rat ventral tegmental area. Eur. J. Neurosci. 2007, 25, 106–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smagin, D.A.; Galyamina, A.G.; Kovalenko, I.L.; Babenko, V.N.; Tamkovich, N.V.; Borisov, S.A.; Tolstikova, T.G.; Kudryavtseva, N.N. Altered expression of neurotransmitter genes in the dorsal striatum of male mice with psychomotor disturbances. Zh. Vyssh. Nerv. Deiat. Im. I. P. Pavlova 2018, 68, 227–249. [Google Scholar]
- Jiao, D.; Liu, Y.; Li, X.; Liu, J.; Zhao, M. The role of the GABA system in amphetamine-type stimulant use disorders. Front. Cell. Neurosci. 2015, 9, 162. [Google Scholar] [CrossRef] [Green Version]
- Crist, R.C.; Reiner, B.C.; Berrettini, W.H. A review of opioid addiction genetics. Curr. Opin. Psychol. 2019, 27, 31–35. [Google Scholar] [CrossRef]
- Trapnell, C.; Hendrickson, D.G.; Sauvageau, M.; Goff, L.; Rinn, J.L.; Pachter, L. Differential analysis of gene regulation at transcript resolution with RNA-seq. Nat. Biotechnol. 2013, 31, 46–53. [Google Scholar] [CrossRef]
- Margolis, E.B.; Lock, H.; Hjelmstad, G.O.; Fields, H.L. The ventral tegmental area revisited: Is there an electrophysiological marker for dopaminergic neurons? J. Physiol. 2006, 577, 907–924. [Google Scholar] [CrossRef]
- Sesack, S.R.; Grace, A.A. Cortico-basal ganglia reward network: Microcircuitry. Neuropsychopharmacology 2010, 35, 27–47. [Google Scholar] [CrossRef] [Green Version]
- Trutti, A.C.; Mulder, M.J.; Hommel, B.; Forstmann, B.U. Functional neuroanatomical review of the ventral tegmental area. NeuroImage 2019, 191, 258–268. [Google Scholar] [CrossRef]
- Walsh, J.J.; Han, M.H. The heterogeneity of ventral tegmental area neurons: Projection functions in a mood-related context. Neuroscience 2014, 282, 101–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, R.; Adermark, L. Dopaminergic regulation of striatal interneurons in reward and addiction: Focus on alcohol. Neural. Plast. 2015, 2015, 814567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nair-Roberts, R.G.; Chatelain-Badie, S.D.; Benson, E.; White-Cooper, H.; Bolam, J.P.; Ungless, M.A. Stereological estimates of dopaminergic, GABAergic and glutamatergic neurons in the ventral tegmental area, substantia nigra and retrorubral field in the rat. Neuroscience 2008, 152, 1024–1031. [Google Scholar] [CrossRef] [Green Version]
- Robison, A.J.; Nestler, E.J. Transcriptional and epigenetic mechanisms of addiction. Nat. Rev. Neurosci. 2011, 12, 623–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calipari, E.S.; Bagot, R.C.; Purushothaman, I.; Davidson, T.J.; Yorgason, J.T.; Pena, C.J.; Walker, D.M.; Pirpinias, S.T.; Guise, K.G.; Ramakrishnan, C.; et al. In vivo imaging identifies temporal signature of D1 and D2 medium spiny neurons in cocaine reward. Proc. Natl. Acad. Sci. USA 2016, 113, 2726–2731. [Google Scholar] [CrossRef] [Green Version]
- Meredith, G.E.; Pennartz, C.M.; Groenewegen, H.J. The cellular framework for chemical signalling in the nucleus accumbens. Prog. Brain Res. 1993, 99, 3–24. [Google Scholar] [CrossRef]
- Olsen, C.M. Natural rewards, neuroplasticity, and non-drug addictions. Neuropharmacology 2011, 61, 1109–1122. [Google Scholar] [CrossRef] [Green Version]
- Luis, C.; Cannella, N.; Spanagel, R.; Kohr, G. Persistent strengthening of the prefrontal cortex—nucleus accumbens pathway during incubation of cocaine-seeking behavior. Neurobiol. Learn. Mem. 2017, 138, 281–290. [Google Scholar] [CrossRef]
- Yager, L.M.; Garcia, A.F.; Wunsch, A.M.; Ferguson, S.M. The ins and outs of the striatum: Role in drug addiction. Neuroscience 2015, 301, 529–541. [Google Scholar] [CrossRef] [Green Version]
- Babenko, V.; Redina, O.; Smagin, D.; Kovalenko, I.; Galyamina, A.; Babenko, R.; Kudryavtseva, N. Dorsal striatum transcriptome profile profound shift in repeated aggression mouse model converged to networks of 12 transcription factors after fighting deprivation. Genes 2021, 13, 21. [Google Scholar] [CrossRef]
- Azmitia, E.C.; Segal, M. An autoradiographic analysis of the differential ascending projections of the dorsal and median raphe nuclei in the rat. J. Comp. Neurol. 1978, 179, 641–667. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.H.; Ikemoto, S. The midbrain raphe nuclei mediate primary reinforcement via GABA(A) receptors. Eur. J. Neurosci. 2007, 25, 735–743. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Highly Aggressive Mice | Aggressive Mice | ||||||
|---|---|---|---|---|---|---|---|
| Attacking behavior | p | ||||||
| Latency, t | 17.50 | ± | 4.52 | 16.18 | ± | 3.25 | |
| Number, n | 7.67 | ± | 2.01 | 7.09 | ± | 0.71 | |
| Total time, t | 146.83 | ± | 14.82 | 65.00 | ± | 7.29 | *** |
| Average time, t/n | 24.24 | ± | 4.87 | 9.98 | ± | 1.33 | ** |
| Diggings, n | 20.33 | ± | 1.26 | 24.46 | ± | 1.45 | * |
| Diggings, t | 64.17 | ± | 6.97 | 78.82 | ± | 7.71 | |
| Hostile behavior | 213.00 | ± | 10.41 | 155.36 | ± | 8.43 | ** |
| Number of mice | 6 | 11 | |||||
| VTA # | |
| Opioidergic system | Oprk1Pomc |
| CAergic systems | Th+ Ddc Slc6a2 Slc6a3 + Snca +Sncb Sncg Adra2c |
| Glutamatergic system | Grid2ip Grm4 Slc17a7 Slc17a8 |
| GABAergic system | Gabra1 Gabrg2 |
| Serotonergic system * | Tph2 Ddc Slc6a4 |
| NAcc | |
| Opioidergic system | Pomc, Pdyn |
| Dopaminergic systems | Th Drd2 |
| Glutamatergic system | Grin3a Slc17a7 |
| GABAergic system | GabrqSlc6a13 |
| Serotonergic system * | Htr2a Htr4 |
| STR # | |
| Opioidergic system | Opcml Oprk1 |
| Dopaminergic systems | Th+ Drd4 + |
| Glutamatergic system | Slc17a7 |
| GABAergic system | Gabra2 Gabra3 Gabrb2 Gabrg2 Gabrg3 |
| Serotonergic system * | - |
| PFC | |
| Opioidergic system | Oprd1 Penk |
| Dopaminergic systems | - |
| Glutamatergic system | Grid2ipGad1 Slc17a8 |
| GABAergic system | - |
| Serotonergic system * | - |
| MRNs # | |
| Opioidergic system | Oprd1 |
| CAergic systems | Th Comt+ Ddc Dbh Drd2 Slc18a2 Sncg |
| Glutamatergic system | Grin3a Grik4 Grm2 Gad1 Gad2 Slc17a7 |
| GABAergic system | Gabra1 Gabra3Gabra4 Gabrb2 Gabrg1 Gabrg2 |
| Serotonergic system * | Tph2 Ddc Slc6a4 Slc18a2 Htr2a Htr3a Htr5b |
| KEGG Term | FDR | No. | Gene List |
|---|---|---|---|
| Nicotine addiction | 1.44 × 10−18 | 12 | Gabra1, Gabra2, Gabra3, Gabra4, Gabrb2, Gabrg1, Gabrg2, Gabrg3, Gabrq, Slc17a7, Slc17a8, Grin3a |
| Retrograde endocannabinoid signaling | 1.21 × 10−10 | 11 | Gabra1, Gabra2, Gabra3, Gabra4, Gabrb2, Gabrg1, Gabrg2, Gabrg3, Gabrq, Slc17a7, Slc17a8 |
| Morphine addiction | 1.71 × 10−9 | 9 | Gabra1, Gabra2, Gabra3, Gabra4, Gabrb2, Gabrg1, Gabrg2, Gabrg3, Gabrq |
| Cocaine addiction | 3.54 × 10−8 | 7 | Th, Ddc, Drd2, Pdyn, Grm2, Grin3a, Slc18a2 |
| Amphetamine addiction | 2.38 × 10−4 | 5 | Th, Ddc, Pdyn, Slc18a2, Grin3a |
| Alcoholism | 0.001 | 6 | Th, Ddc, Drd2, Pdyn, Slc18a2, Grin3a |
| Synaptic vesicle cycle | 3.25 × 10−4 | 5 | Slc6a2, Slc6a13, Slc17a7, Slc17a8, Slc18a2 |
| Taste transduction | 5.85 × 10−4 | 5 | Gabra1, Gabra2, Gabra3, Gabra4, Grm4 |
| Glutamatergic synapse | 1.12 × 10−4 | 6 | Slc17a7, Slc17a8, Grm2, Grm4, Grin3a, Grik4 |
| GABAergic synapse | 1.37 × 10−14 | 12 | Gabra1, Gabra2, Gabra3, Gabra4, Gabrb2, Gabrg1, Gabrg2, Gabrg3, Gabrq, Slc6a13, Gad1, Gad2 |
| Neuroactive ligand-receptor interaction | 1.44 × 10−18 | 20 | Drd2, Oprd1, Oprk1, Penk, Pdyn, Pomc, Adra2c, Gabra1, Gabra2, Gabra3, Gabra4, Gabrb2, Gabrg1, Gabrg2, Gabrg3, Gabrq, Grm2, Grm4, Grik4, Grin3a |
| Tyrosine metabolism | 0.018 | 3 | Th, Ddc, Dbh |
| Dopaminergic synapse | 0.020 | 4 | Th, Ddc, Drd2, Slc18a2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smagin, D.A.; Galyamina, A.G.; Kovalenko, I.L.; Kudryavtseva, N.N. Altered Expression of Genes Associated with Major Neurotransmitter Systems in the Reward-Related Brain Regions of Mice with Positive Fighting Experience. Int. J. Mol. Sci. 2022, 23, 13644. https://doi.org/10.3390/ijms232113644
Smagin DA, Galyamina AG, Kovalenko IL, Kudryavtseva NN. Altered Expression of Genes Associated with Major Neurotransmitter Systems in the Reward-Related Brain Regions of Mice with Positive Fighting Experience. International Journal of Molecular Sciences. 2022; 23(21):13644. https://doi.org/10.3390/ijms232113644
Chicago/Turabian StyleSmagin, Dmitry A., Anna G. Galyamina, Irina L. Kovalenko, and Natalia N. Kudryavtseva. 2022. "Altered Expression of Genes Associated with Major Neurotransmitter Systems in the Reward-Related Brain Regions of Mice with Positive Fighting Experience" International Journal of Molecular Sciences 23, no. 21: 13644. https://doi.org/10.3390/ijms232113644
APA StyleSmagin, D. A., Galyamina, A. G., Kovalenko, I. L., & Kudryavtseva, N. N. (2022). Altered Expression of Genes Associated with Major Neurotransmitter Systems in the Reward-Related Brain Regions of Mice with Positive Fighting Experience. International Journal of Molecular Sciences, 23(21), 13644. https://doi.org/10.3390/ijms232113644