

Fertilization, but Not Post-Implantation Development, Can Occur in the Absence of Sperm and Oocyte Beta1 Integrin in Mice

 , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Efficiency of the Cre-Mediated Deletion of the Itgb1 Gene in Sperm and Oocytes
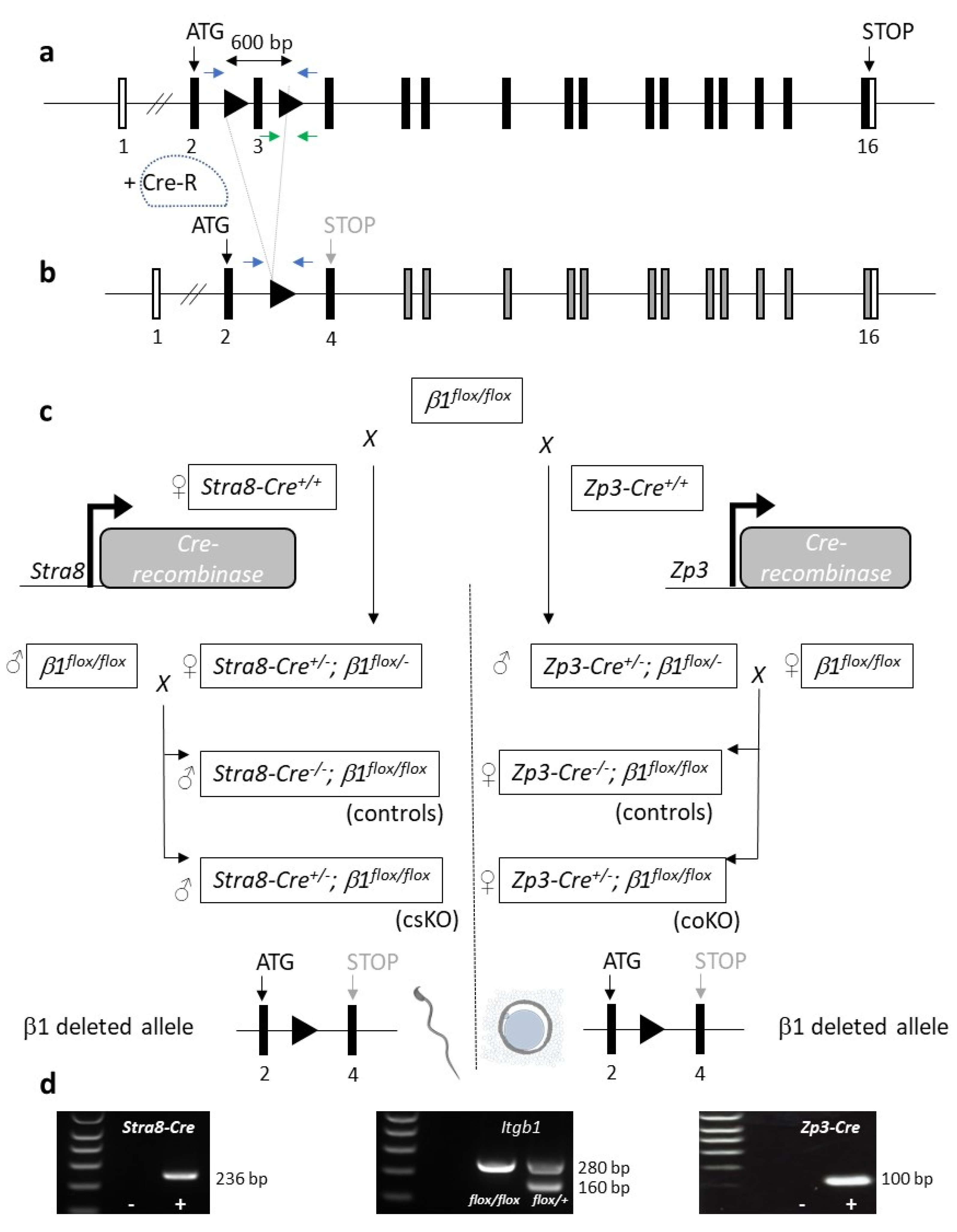
2.1.1. Diagram of the Mating Used to Generate Conditional Knockouts on Sperm and Oocyte
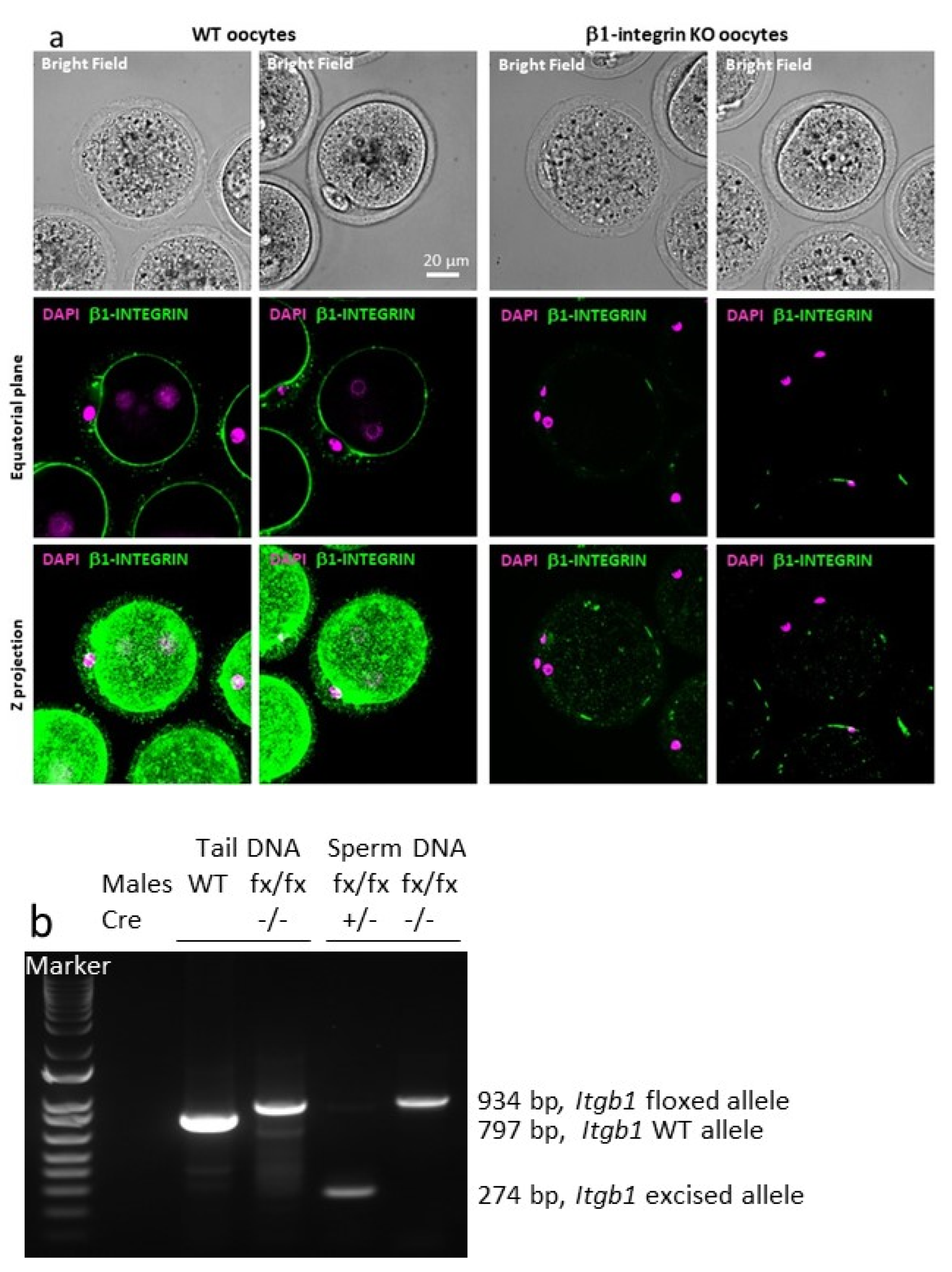
2.1.2. Efficiency of the Cre-Mediated Deletion of the Itgb1 Gene in Oocytes and Sperm
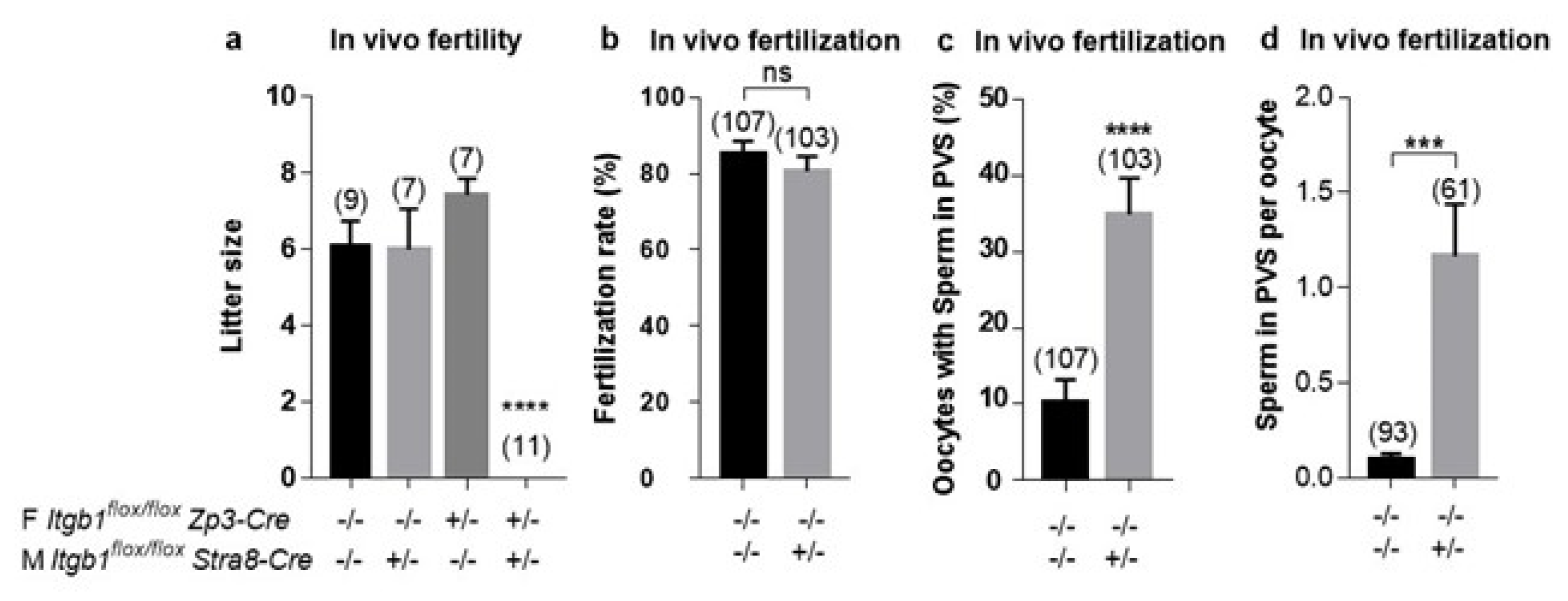
2.2. In Vivo Evaluation of the Fertilizing Ability of Sperm from Stra8-Cre +/− Itgb1 flox/flox Males
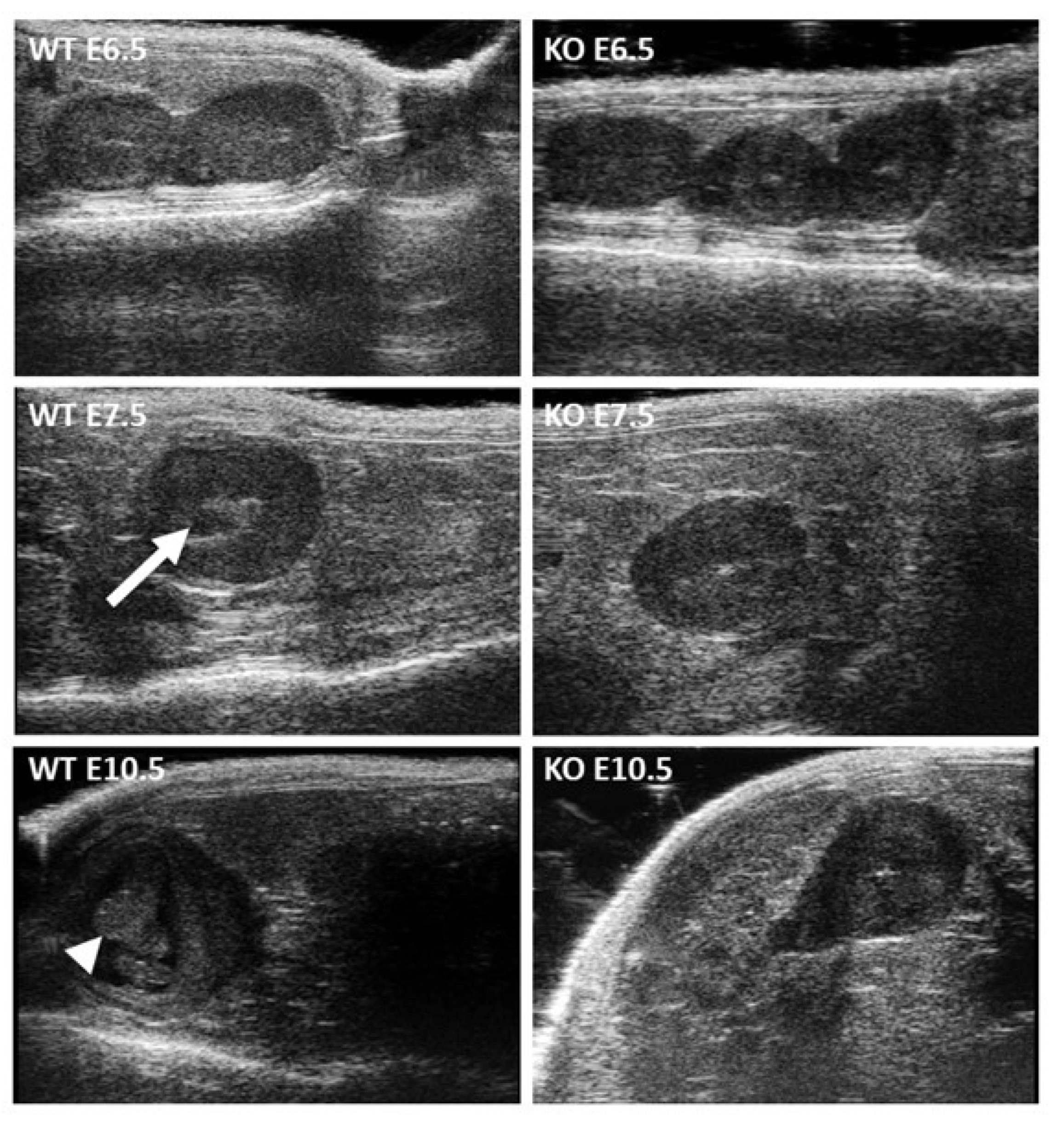
2.3. Peri-Implantation Lethality of Embryos Resulting from Crosses of Stra8-Cre +/− Itgb1 flox/flox Males and Zp3-Cre +/− Itgb1 flox/flox Females
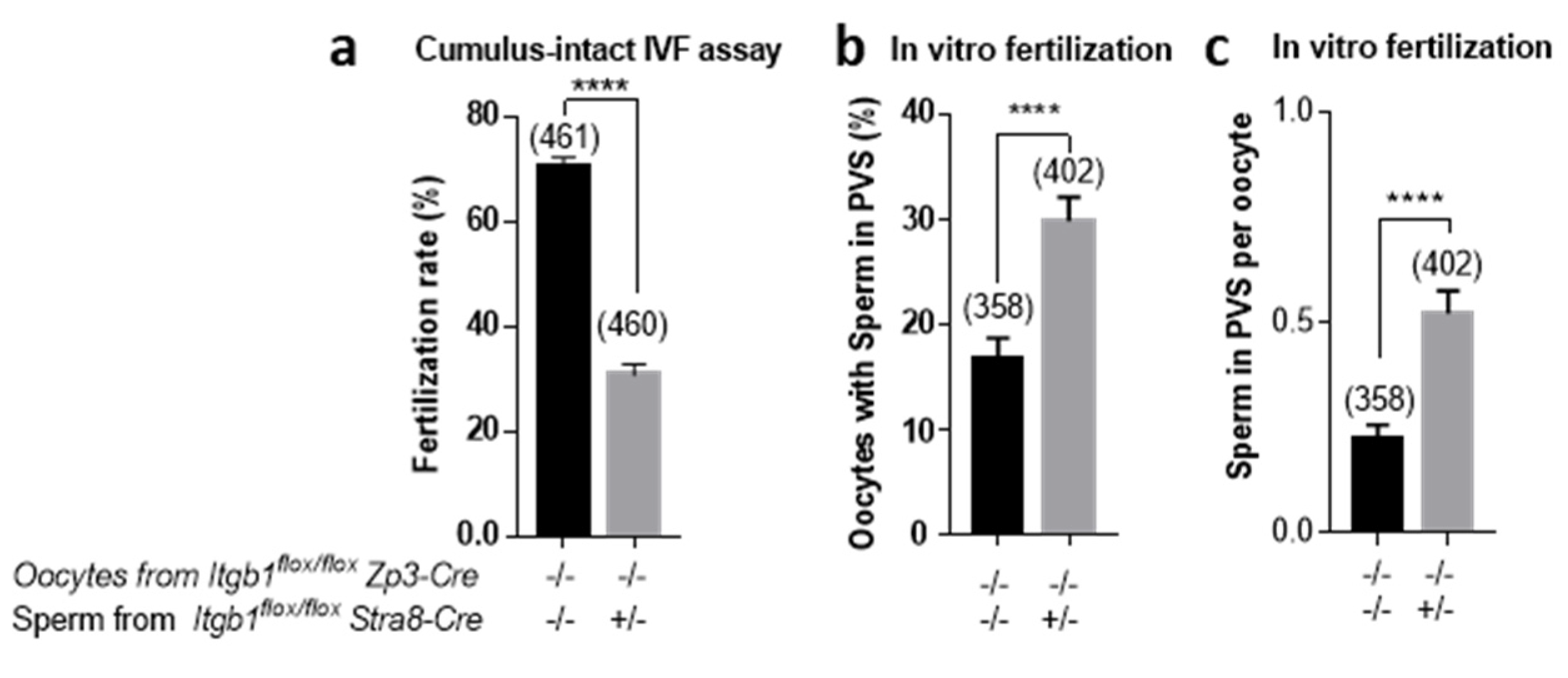
2.4. In Vitro Evaluation of the Fertilizing Ability of Stra8-Cre +/− Itgb1 flox/flox Sperm
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Generation of Oocyte and Sperm Itgb1 Conditional Knockout Mice
4.3. Immunofluorescence of Mouse Embryos
4.4. PCR on Mouse Tail and Sperm DNA
4.5. In Vivo Mating, Gamete Preparation and in vitro Fertilization
4.6. High Frequency Ultrasonography Implantation and Survival Assessment
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kaji, K.; Oda, S.; Shikano, T.; Ohnuki, T.; Uematsu, Y.; Sakagami, J.; Tada, N.; Miyazaki, S.; Kudo, A. The gamete fusion process is defective in eggs of Cd9-deficient mice. Nat. Genet. 2000, 24, 279–282. [Google Scholar] [CrossRef] [PubMed]
- Le Naour, F.; Rubinstein, E.; Jasmin, C.; Prenant, M.; Boucheix, C. Severely reduced female fertility in CD9-deficient mice. Science 2000, 287, 319–321. [Google Scholar] [CrossRef] [PubMed]
- Miyado, K.; Yamada, G.; Yamada, S.; Hasuwa, H.; Nakamura, Y.; Ryu, F.; Suzuki, K.; Kosai, K.; Inoue, K.; Ogura, A.; et al. Requirement of CD9 on the egg plasma membrane for fertilization. Science 2000, 287, 321–324. [Google Scholar] [CrossRef] [PubMed]
- Inoue, N.; Ikawa, M.; Isotani, A.; Okabe, M. The immunoglobulin superfamily protein Izumo is required for sperm to fuse with eggs. Nature 2005, 434, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, E.; Doe, B.; Goulding, D.; Wright, G.J. Juno is the egg Izumo receptor and is essential for mammalian fertilization. Nature 2014, 508, 483–487. [Google Scholar] [CrossRef] [Green Version]
- Barbaux, S.; Ialy-Radio, C.; Chalbi, M.; Dybal, E.; Homps-Legrand, M.; Do Cruzeiro, M.; Vaiman, D.; Wolf, J.P.; Ziyyat, A. Sperm SPACA6 protein is required for mammalian Sperm-Egg Adhesion/Fusion. Sci. Rep. 2020, 10, 5335. [Google Scholar] [CrossRef] [Green Version]
- Fujihara, Y.; Lu, Y.; Noda, T.; Oji, A.; Larasati, T.; Kojima-Kita, K.; Yu, Z.; Matzuk, R.M.; Matzuk, M.M.; Ikawa, M. Spermatozoa lacking Fertilization Influencing Membrane Protein (FIMP) fail to fuse with oocytes in mice. Proc. Natl. Acad. Sci. USA 2020, 117, 9393–9400. [Google Scholar] [CrossRef] [Green Version]
- Lamas-Toranzo, I.; Hamze, J.G.; Bianchi, E.; Fernandez-Fuertes, B.; Perez-Cerezales, S.; Laguna-Barraza, R.; Fernandez-Gonzalez, R.; Lonergan, P.; Gutierrez-Adan, A.; Wright, G.J.; et al. TMEM95 is a sperm membrane protein essential for mammalian fertilization. elife 2020, 9, e53913. [Google Scholar] [CrossRef]
- Noda, T.; Lu, Y.; Fujihara, Y.; Oura, S.; Koyano, T.; Kobayashi, S.; Matzuk, M.M.; Ikawa, M. Sperm proteins SOF1, TMEM95, and SPACA6 are required for sperm-oocyte fusion in mice. Proc. Natl. Acad. Sci. USA 2020, 117, 11493–11502. [Google Scholar] [CrossRef]
- Siu, K.K.; Serrão, V.H.B.; Ziyyat, A.; Lee, J.E. The cell biology of fertilization: Gamete attachment and fusion. J. Cell Biol. 2021, 220, 2146. [Google Scholar] [CrossRef]
- Vondrakova, J.; Frolikova, M.; Ded, L.; Cerny, J.; Postlerova, P.; Palenikova, V.; Simonik, O.; Nahacka, Z.; Basus, K.; Valaskova, E.; et al. MAIA, Fc receptor-like 3, supersedes JUNO as IZUMO1 receptor during human fertilization. Sci. Adv. 2022, 8, eabn0047. [Google Scholar] [CrossRef]
- Almeida, E.A.; Huovila, A.P.; Sutherland, A.E.; Stephens, L.E.; Calarco, P.G.; Shaw, L.M.; Mercurio, A.M.; Sonnenberg, A.; Primakoff, P.; Myles, D.G.; et al. Mouse egg integrin alpha 6 beta 1 functions as a sperm receptor. Cell 1995, 81, 1095–1104. [Google Scholar] [CrossRef] [Green Version]
- Miller, B.J.; Georges-Labouesse, E.; Primakoff, P.; Myles, D.G. Normal fertilization occurs with eggs lacking the integrin alpha6beta1 and is CD9-dependent. J. Cell Biol. 2000, 149, 1289–1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Z.Y.; Brakebusch, C.; Fassler, R.; Kreidberg, J.A.; Primakoff, P.; Myles, D.G. None of the integrins known to be present on the mouse egg or to be ADAM receptors are essential for sperm-egg binding and fusion. Dev. Biol. 2003, 254, 226–237. [Google Scholar] [CrossRef] [Green Version]
- Schwander, M.; Leu, M.; Stumm, M.; Dorchies, O.M.; Ruegg, U.T.; Schittny, J.; Muller, U. Beta1 integrins regulate myoblast fusion and sarcomere assembly. Dev. Cell 2003, 4, 673–685. [Google Scholar] [CrossRef] [Green Version]
- Barraud-Lange, V.; Ialy-Radio, C.; Chalas, C.; Holtzmann, I.; Wolf, J.P.; Barbaux, S.; Ziyyat, A. Partial Sperm beta1 Integrin Subunit Deletion Proves its Involvement in Mouse Gamete Adhesion/Fusion. Int. J. Mol. Sci. 2020, 21, 8494. [Google Scholar] [CrossRef]
- Sadate-Ngatchou, P.I.; Payne, C.J.; Dearth, A.T.; Braun, R.E. Cre recombinase activity specific to postnatal, premeiotic male germ cells in transgenic mice. Genesis 2008, 46, 738–742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raghavan, S.; Bauer, C.; Mundschau, G.; Li, Q.; Fuchs, E. Conditional ablation of beta1 integrin in skin. Severe defects in epidermal proliferation, basement membrane formation, and hair follicle invagination. J. Cell Biol. 2000, 150, 1149–1160. [Google Scholar] [CrossRef] [Green Version]
- Stephens, L.E.; Sutherland, A.E.; Klimanskaya, I.V.; Andrieux, A.; Meneses, J.; Pedersen, R.A.; Damsky, C.H. Deletion of beta 1 integrins in mice results in inner cell mass failure and peri-implantation lethality. Genes Dev. 1995, 9, 1883–1895. [Google Scholar] [CrossRef] [Green Version]
- Fassler, R.; Meyer, M. Consequences of lack of beta 1 integrin gene expression in mice. Genes Dev. 1995, 9, 1896–1908. [Google Scholar] [CrossRef]
- Burkart, A.D.; Xiong, B.; Baibakov, B.; Jimenez-Movilla, M.; Dean, J. Ovastacin, a cortical granule protease, cleaves ZP2 in the zona pellucida to prevent polyspermy. J. Cell Biol. 2012, 197, 37–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Vries, W.N.; Binns, L.T.; Fancher, K.S.; Dean, J.; Moore, R.; Kemler, R.; Knowles, B.B. Expression of Cre recombinase in mouse oocytes: A means to study maternal effect genes. Genesis 2000, 26, 110–112. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mating | Implanted Embryos at E6.5 | Implanted Embryos at E10.5 | Birth | Survival Rate |
|---|---|---|---|---|
| ♀Ctrl × ♂♂Ctrl | 5 | 5 | 5 | 100% |
| 5 | 5 | 5 | ||
| 5 | 5 | 5 | ||
| 8 | 8 | 8 | ||
| ♀coKO × ♂♂csKO | 9 | 0 | 0 | 0% |
| 7 | 0 | 0 | ||
| 5 | 0 | 0 | ||
| 7 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mimouni, N.E.H.; Ialy-Radio, C.; Denizot, A.-L.; Lagoutte, I.; Frolikova, M.; Komrskova, K.; Barbaux, S.; Ziyyat, A. Fertilization, but Not Post-Implantation Development, Can Occur in the Absence of Sperm and Oocyte Beta1 Integrin in Mice. Int. J. Mol. Sci. 2022, 23, 13812. https://doi.org/10.3390/ijms232213812
Mimouni NEH, Ialy-Radio C, Denizot A-L, Lagoutte I, Frolikova M, Komrskova K, Barbaux S, Ziyyat A. Fertilization, but Not Post-Implantation Development, Can Occur in the Absence of Sperm and Oocyte Beta1 Integrin in Mice. International Journal of Molecular Sciences. 2022; 23(22):13812. https://doi.org/10.3390/ijms232213812
Chicago/Turabian StyleMimouni, Nour El Houda, Côme Ialy-Radio, Anne-Lyse Denizot, Isabelle Lagoutte, Michaela Frolikova, Katerina Komrskova, Sandrine Barbaux, and Ahmed Ziyyat. 2022. "Fertilization, but Not Post-Implantation Development, Can Occur in the Absence of Sperm and Oocyte Beta1 Integrin in Mice" International Journal of Molecular Sciences 23, no. 22: 13812. https://doi.org/10.3390/ijms232213812
APA StyleMimouni, N. E. H., Ialy-Radio, C., Denizot, A. -L., Lagoutte, I., Frolikova, M., Komrskova, K., Barbaux, S., & Ziyyat, A. (2022). Fertilization, but Not Post-Implantation Development, Can Occur in the Absence of Sperm and Oocyte Beta1 Integrin in Mice. International Journal of Molecular Sciences, 23(22), 13812. https://doi.org/10.3390/ijms232213812