
Adaptation of Saccharomyces Species to High-Iron Conditions

 , ,
, , 
Abstract
:1. Introduction
2. Results
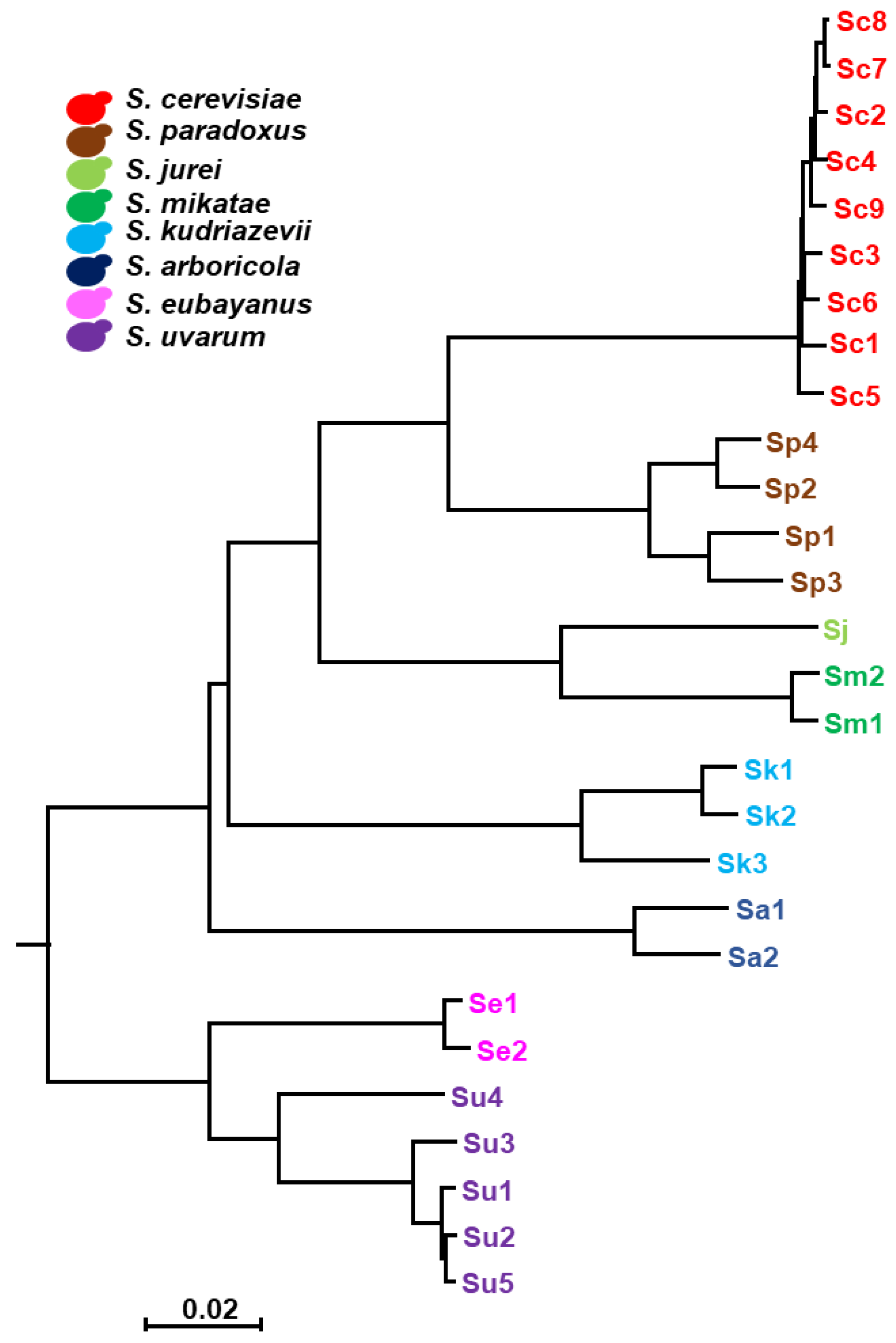
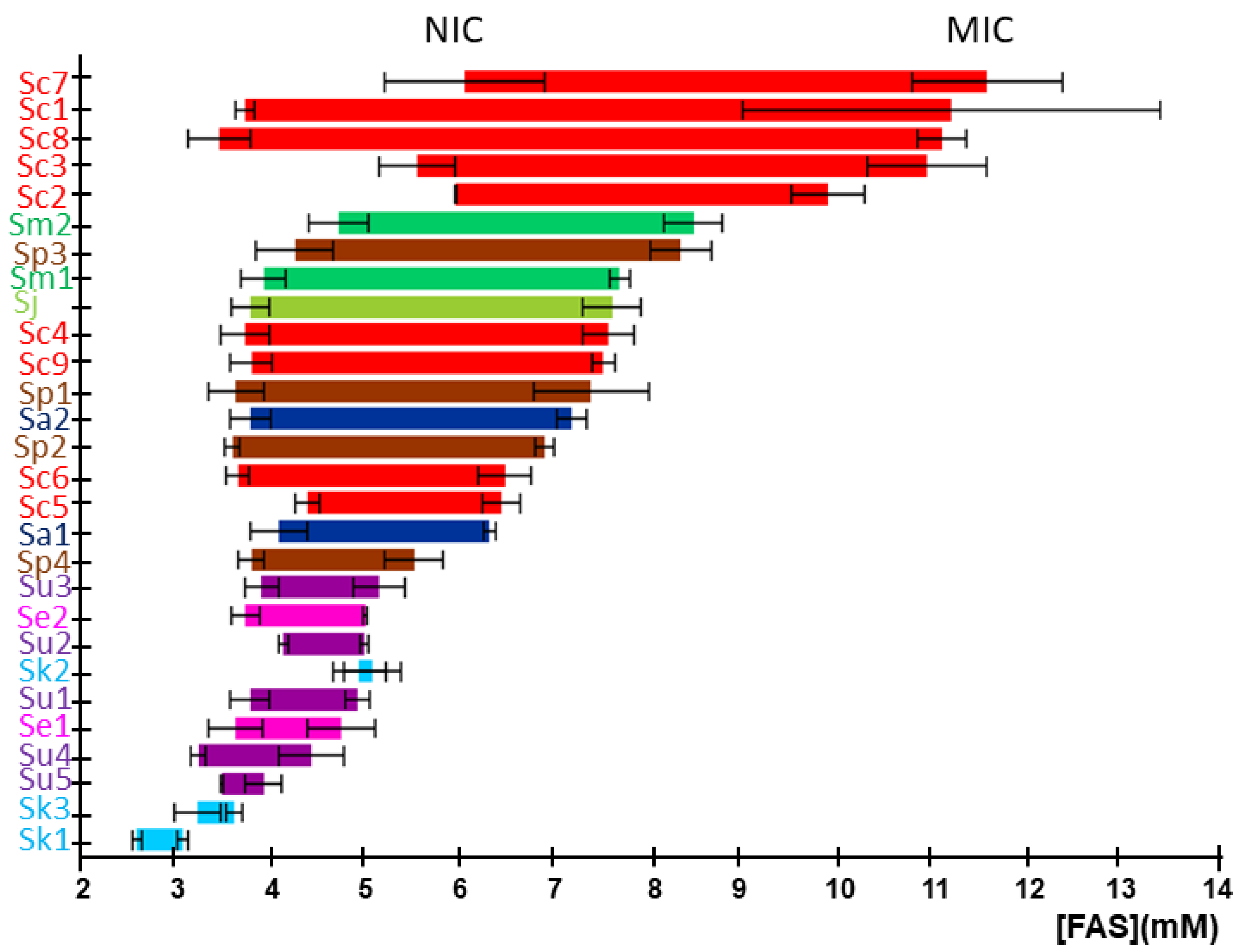
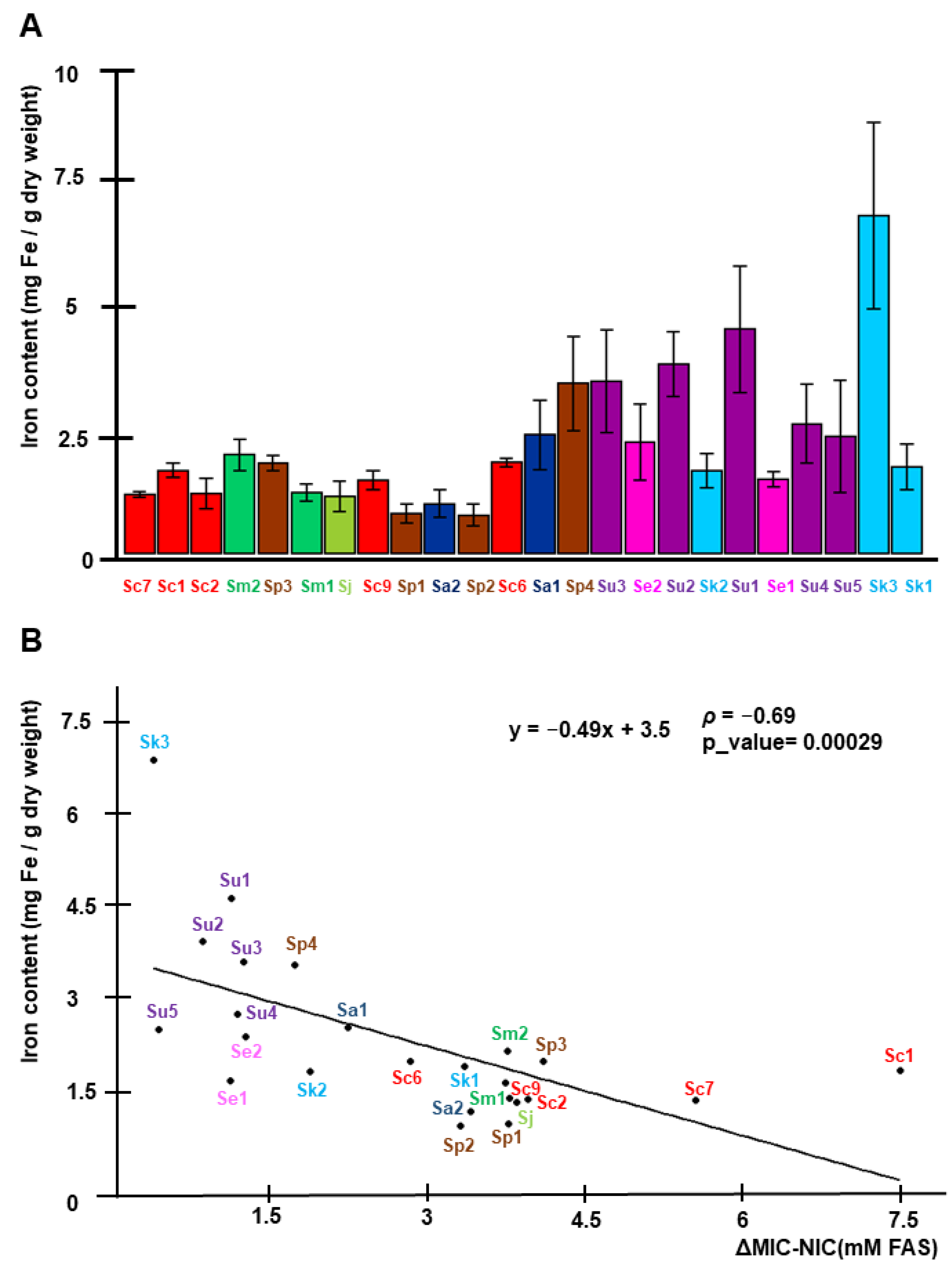
2.1. Characterization of Iron Resistance and Accumulation in the Saccharomyces Genus
2.2. Iron-Sensitive Strains Show Alterations in Cellular Redox State which Are Not Due to a Diminished Response to Oxidative Stress
2.3. Iron-Sensitive Strains Are Less Thermotolerant
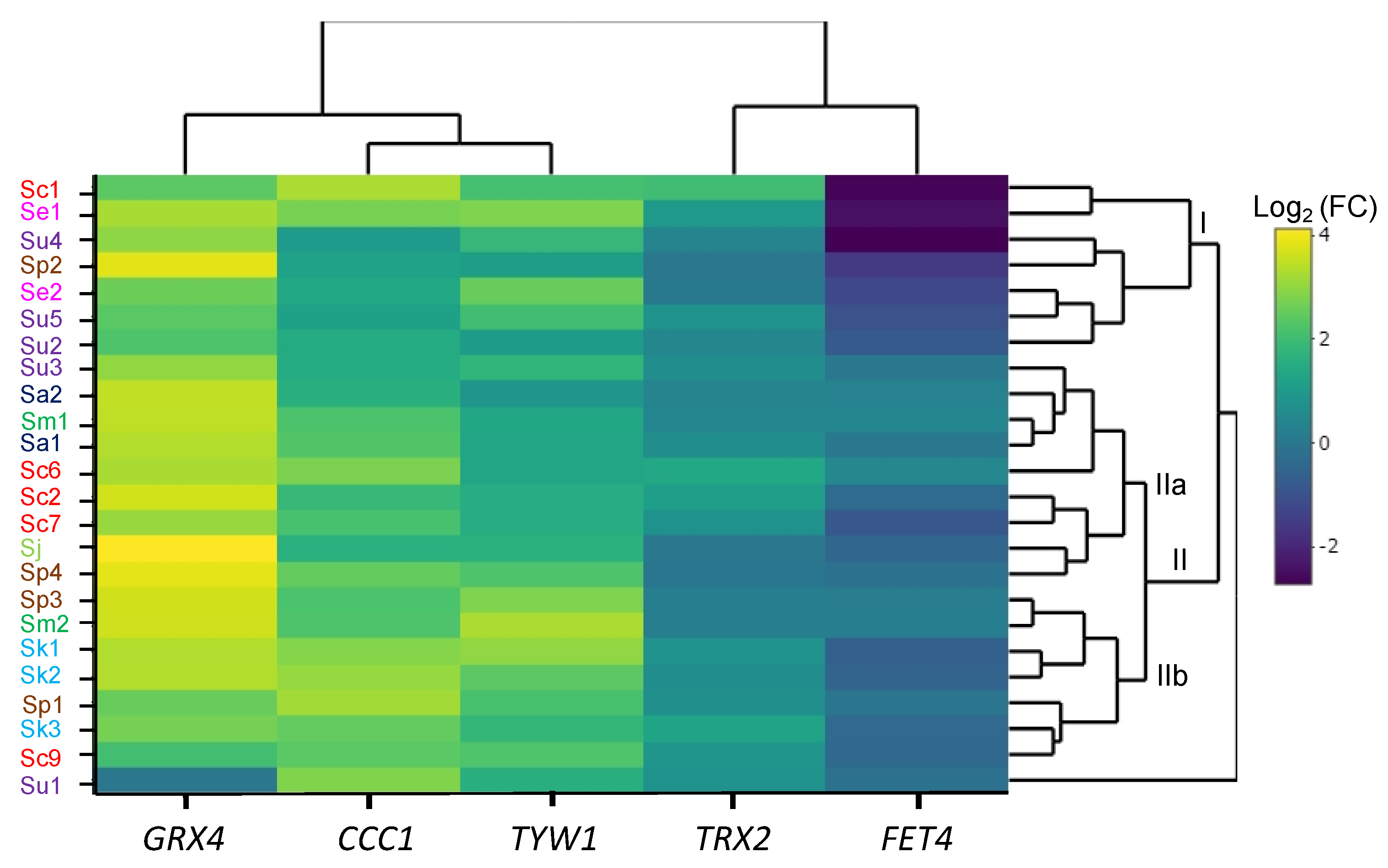
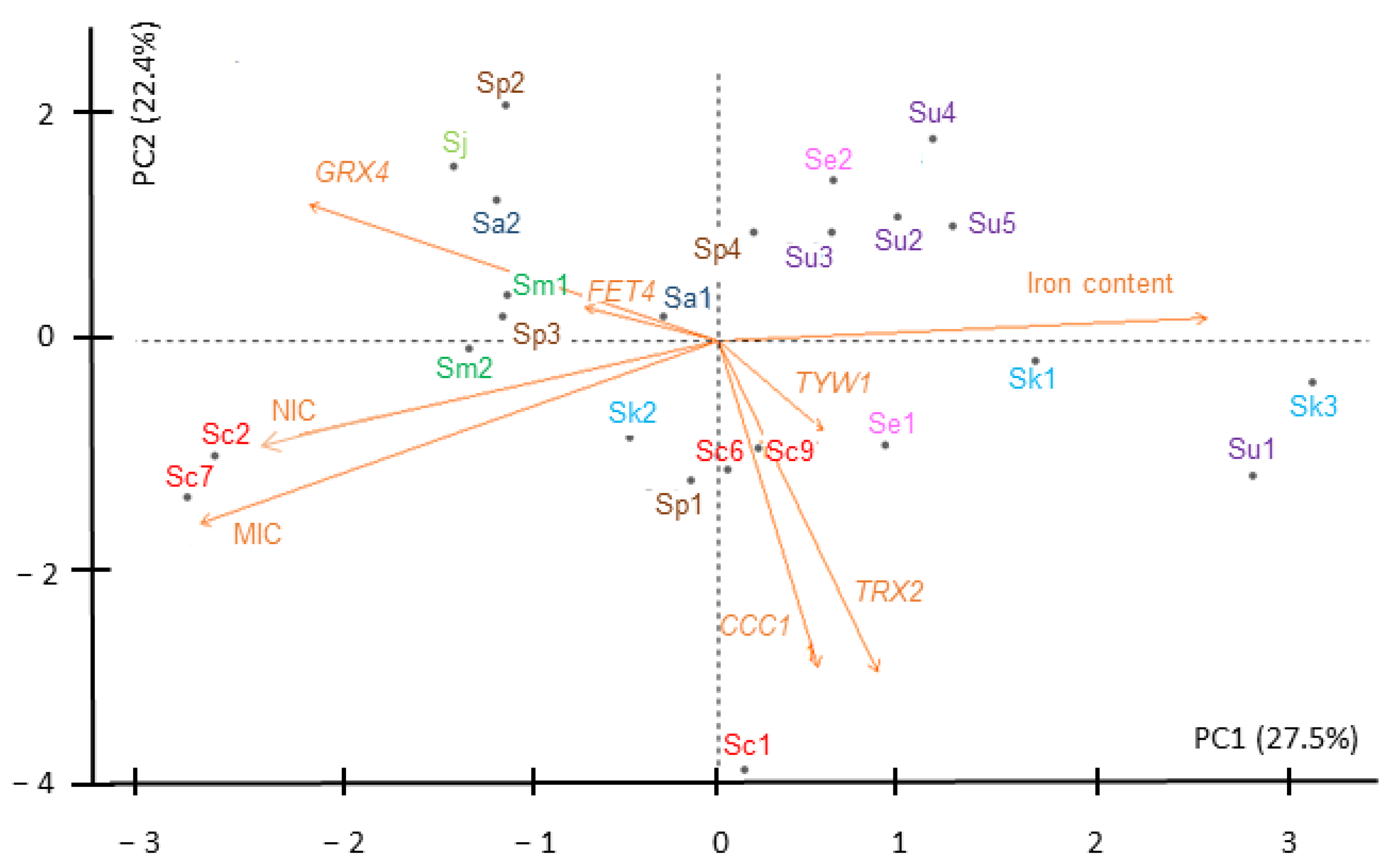
2.4. Iron-Sensitive Strains Show Altered Expression of Iron Homeostasis and Oxidative Response Genes
2.5. Iron-Sensitive Strains Have Impairments on Iron Toxicity Response
3. Discussion
4. Materials and Methods
4.1. Yeast Strains and Growth Conditions
4.2. Determination of Non-Inhibitory and Minimal Inhibitory Concentrations
4.3. Endogenous Iron Measurements
4.4. RNA Processing and Analysis
4.5. Phylogenetic Tree
4.6. Gene Expression Heatmap
4.7. Principal Component Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Puig, S.; Ramos-Alonso, L.; Romero, A.M.; Martinez-Pastor, M.T. The elemental role of iron in DNA synthesis and repair. Metallomics 2017, 9, 1483–1500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jorda, T.; Puig, S. Regulation of Ergosterol Biosynthesis in Saccharomyces cerevisiae. Genes 2020, 11, 795. [Google Scholar] [CrossRef] [PubMed]
- Romero, A.M.; Martinez-Pastor, M.T.; Puig, S. Iron in Translation: From the Beginning to the End. Microorganisms 2021, 9, 1058. [Google Scholar] [CrossRef] [PubMed]
- Sanvisens, N.; Puig, S. Causes and Consequences of Nutritional Iron Deficiency in Living Organisms; Nova Science Publishers Inc.: New York, USA, 2011. [Google Scholar]
- Martinez-Pastor, M.T.; Puig, S. Adaptation to iron deficiency in human pathogenic fungi. Biochim. Biophys. Acta Mol. Cell Res. 2020, 1867, 118797. [Google Scholar] [CrossRef]
- Martinez-Pastor, M.T.; Perea-Garcia, A.; Puig, S. Mechanisms of iron sensing and regulation in the yeast Saccharomyces cerevisiae. World J. Microbiol. Biotechnol. 2017, 33, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winterbourn, C.C. Toxicity of oxygen and hydrogen peroxide: The Fenton reaction. Toxicol. Lett. 1995, 82/83, 969–974. [Google Scholar] [CrossRef]
- Lee, Y.J.; Huang, X.; Kropat, J.; Henras, A.; Merchant, S.S.; Dickson, R.C.; Chanfreau, G.F. Sphingolipid signaling mediates iron toxicity. Cell Metab. 2012, 16, 90–96. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Cobine, P.A.; Molik, S.; Naranuntarat, A.; Lill, R.; Winge, D.R.; Culotta, V.C. The effects of mitochondrial iron homeostasis on cofactor specificity of superoxide dismutase 2. EMBO J. 2006, 25, 1775–1783. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.; Li, L.; Jia, X.; Ward, D.M.; Kaplan, J. Genetic and biochemical analysis of high iron toxicity in yeast: Iron toxicity is due to the accumulation of cytosolic iron and occurs under both aerobic and anaerobic conditions. J. Biol. Chem. 2011, 286, 3851–3862. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Jia, X.; Ward, D.M.; Kaplan, J. Yap5 protein-regulated transcription of the TYW1 gene protects yeast from high iron toxicity. J. Biol. Chem. 2011, 286, 38488–38497. [Google Scholar] [CrossRef]
- Pimentel, C.; Vicente, C.; Menezes, R.A.; Caetano, S.; Carreto, L.; Rodrigues-Pousada, C. The role of the Yap5 transcription factor in remodeling gene expression in response to Fe bioavailability. PLoS ONE 2012, 7, e37434. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Garay, C.A.; de Llanos, R.; Romero, A.M.; Martinez-Pastor, M.T.; Puig, S. Responses of Saccharomyces cerevisiae Strains from Different Origins to Elevated Iron Concentrations. Appl. Environ. Microbiol. 2016, 82, 1906–1916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.N.; Mostovoy, Y.; Hsu, T.Y.; Chang, A.H.; Brem, R.B. Divergence of iron metabolism in wild Malaysian yeast. G3 Genes Genomes Genet. 2013, 3, 2187–2194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eldarov, M.A.; Beletsky, A.V.; Tanashchuk, T.N.; Kishkovskaya, S.A.; Ravin, N.V.; Mardanov, A.V. Whole-Genome Analysis of Three Yeast Strains Used for Production of Sherry-Like Wines Revealed Genetic Traits Specific to Flor Yeasts. Front. Microbiol. 2018, 9, 965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balaban, B.G.; Yilmaz, U.; Alkim, C.; Topaloglu, A.; Kisakesen, H.I.; Holyavkin, C.; Cakar, Z.P. Evolutionary Engineering of an Iron-Resistant Saccharomyces cerevisiae Mutant and Its Physiological and Molecular Characterization. Microorganisms 2019, 8, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peris, D.; Ubbelohde, E.J.; Kuang, M.C.; Kominek, J.; Langdon, Q.K.; Adams, M.; Koshalek, J.A.; Hulfachor, A.B.; Opulente, D.A.; Hall, D.J.; et al. Macroevolutionary diversity of traits and genomes in the model yeast genus Saccharomyces. bioRxiv 2022. [Google Scholar] [CrossRef]
- Peter, J.; De Chiara, M.; Friedrich, A.; Yue, J.X.; Pflieger, D.; Bergstrom, A.; Sigwalt, A.; Barre, B.; Freel, K.; Llored, A.; et al. Genome evolution across 1,011 Saccharomyces cerevisiae isolates. Nature 2018, 556, 339–344. [Google Scholar] [CrossRef] [Green Version]
- Alsammar, H.; Delneri, D. An update on the diversity, ecology and biogeography of the Saccharomyces genus. FEMS Yeast Res. 2020, 20. [Google Scholar] [CrossRef] [Green Version]
- Salvado, Z.; Arroyo-Lopez, F.N.; Guillamon, J.M.; Salazar, G.; Querol, A.; Barrio, E. Temperature adaptation markedly determines evolution within the genus Saccharomyces. Appl. Environ. Microbiol. 2011, 77, 2292–2302. [Google Scholar] [CrossRef] [Green Version]
- Nishida, K.; Silver, P.A. Induction of biogenic magnetization and redox control by a component of the target of rapamycin complex 1 signaling pathway. PLoS Biol. 2012, 10, e1001269. [Google Scholar] [CrossRef]
- Park, J.; McCormick, S.P.; Chakrabarti, M.; Lindahl, P.A. The lack of synchronization between iron uptake and cell growth leads to iron overload in Saccharomyces cerevisiae during post-exponential growth modes. Biochemistry 2013, 52, 9413–9425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Miao, R.; Bertram, S.; Jia, X.; Ward, D.M.; Kaplan, J. A role for iron-sulfur clusters in the regulation of transcription factor Yap5-dependent high iron transcriptional responses in yeast. J. Biol. Chem. 2012, 287, 35709–35721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Ward, D.M. Iron toxicity in yeast: Transcriptional regulation of the vacuolar iron importer Ccc1. Curr. Genet. 2018, 64, 413–416. [Google Scholar] [CrossRef] [PubMed]
- Jennings, D.H. Stress Tolerance of Fungi; Taylor & Francis Group: New York, NY, USA, 1993; Volume 10. [Google Scholar]
- Li, L.; Murdock, G.; Bagley, D.; Jia, X.; Ward, D.M.; Kaplan, J. Genetic dissection of a mitochondria-vacuole signaling pathway in yeast reveals a link between chronic oxidative stress and vacuolar iron transport. J. Biol. Chem. 2010, 285, 10232–10242. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Miao, R.; Jia, X.; Ward, D.M.; Kaplan, J. Expression of the yeast cation diffusion facilitators Mmt1 and Mmt2 affects mitochondrial and cellular iron homeostasis: Evidence for mitochondrial iron export. J. Biol. Chem. 2014, 289, 17132–17141. [Google Scholar] [CrossRef] [Green Version]
- Bukhman, Y.V.; DiPiazza, N.W.; Piotrowski, J.; Shao, J.; Halstead, A.G.W.; Bui, M.D.; Xie, E.; Sato, T.K. Modeling Microbial Growth Curves with GCAT. BioEnergy Res. 2015, 8, 1022–1030. [Google Scholar] [CrossRef]
- Arroyo-Lopez, F.N.; Salvado, Z.; Tronchoni, J.; Guillamon, J.M.; Barrio, E.; Querol, A. Susceptibility and resistance to ethanol in Saccharomyces strains isolated from wild and fermentative environments. Yeast 2010, 27, 1005–1015. [Google Scholar] [CrossRef] [Green Version]
- Team’, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 20 September 2022).
- Elzhov, T.V.; Mullen, K.M.; Spiess, A.; Bolker, B. R Interface to the Levenberg-Marquardt Nonlinear Least-Squares. Algorithm Found in MINPACK, Plus Support for Bounds. 2022. R package Version 1.2-2. Available online: https://CRAN.R-project.org/package=minpack.lm (accessed on 20 September 2022).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. Available online: https://ggplot2.tidyverse.org (accessed on 20 September 2022).
- Tamarit, J.; Irazusta, V.; Moreno-Cermeno, A.; Ros, J. Colorimetric assay for the quantitation of iron in yeast. Anal. Biochem. 2006, 351, 149–151. [Google Scholar] [CrossRef]
- Sanvisens, N.; Romero, A.M.; An, X.; Zhang, C.; de Llanos, R.; Martinez-Pastor, M.T.; Bano, M.C.; Huang, M.; Puig, S. Yeast Dun1 kinase regulates ribonucleotide reductase inhibitor Sml1 in response to iron deficiency. Mol. Cell. Biol. 2014, 34, 3259–3271. [Google Scholar] [CrossRef] [Green Version]
- Jain, C.; Rodriguez, R.L.; Phillippy, A.M.; Konstantinidis, K.T.; Aluru, S. High throughput ANI analysis of 90K prokaryotic genomes reveals clear species boundaries. Nat. Commun. 2018, 9, 5114. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paradis, E.; Schliep, K. ape 5.0: An environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 2019, 35, 526–528. [Google Scholar] [CrossRef] [PubMed]
- Revell, L.J. phytools: An R package for phylogenetic comparative biology (and other things). Methods Ecol. Evol. 2012, 3, 217–223. [Google Scholar] [CrossRef]
- Yu, G.; Smith, D.K.; Zhu, H.; Guan, Y.; Lam, T.T.Y.; McInerny, G. ggtree: An r package for visualization and annotation of phylogenetic trees with their covariates and other associated data. Methods Ecol. Evol. 2016, 8, 28–36. [Google Scholar] [CrossRef]
- Galili, T.; O’Callaghan, A.; Sidi, J.; Sievert, C. heatmaply: An R package for creating interactive cluster heatmaps for online publishing. Bioinformatics 2018, 34, 1600–1602. [Google Scholar] [CrossRef] [Green Version]
- Kassambara, A.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses. 2020. R package version 1.0.7. Available online: https://CRAN.R-project.org/package=factoextra (accessed on 20 September 2022).
- Tang, Y.; Horikoshi, M.; Li, W. ggfortify: Unified Interface to Visualize Statistical Result of Popular R Packages. R J. 2016, 8, 474–485. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Species | Strain | Geographical Origin | Population * |
|---|---|---|---|---|
| Sc1 | S. cerevisiae | UWOPS03-461.4 | Asia (Malaysia) | 19. Malaysian |
| Sc2 | S. cerevisiae | CECT10131 | Europe (Spain) | 8. Mixed origin |
| Sc3 | S. cerevisiae | CECT10711 | Asia (Japan) | 25. Sake |
| Sc4 | S. cerevisiae | CECT10120 | Europe (Spain) | 8. Mixed origin |
| Sc5 | S. cerevisiae | CLIB219-2B | Europe (Russia) | 18. Far East Asia |
| Sc6 | S. cerevisiae | YPS128 | USA (Pennsylvania) | 23. North American oak |
| Sc7 | S. cerevisiae | YJM978 | Europe (Italy) | 1. Wine/European |
| Sc8 | S. cerevisiae | CECT11032 | Europe (Italy) | 1. Wine/European |
| Sc9 | S. cerevisiae | BY4743 | ||
| Sp1 | S. paradoxus | yHDPN24 | Canada (Québec) | America C |
| Sp2 | S. paradoxus | N44 | Europe (Russia) | Far East |
| Sp3 | S. paradoxus | YPS138 | USA (Pennsylvania) | America B |
| Sp4 | S. paradoxus | CBS432 | Europe (Russia) | EU |
| Sj | S. jurei | NCYC3947 | Europe (France) | EU |
| Sm1 | S. mikatae | yHAB336 | Asia (China) | Asia B |
| Sm2 | S. mikatae | IFO1815 | Asia (Japan) | Asia A |
| Sk1 | S. kudriazevii | IFO1802 | Asia (Japan) | Asia A |
| Sk2 | S. kudriazevii | ZP591 | Europe (Portugal) | EU |
| Sk3 | S. kudriazevii | IFO1803 | Asia (Japan) | Asia B |
| Sa1 | S. arboricola | CBS10644 | Asia (China) | Asia A |
| Sa2 | S. arboricola | ZP960 | Oceania (New Zealand) | Oceania |
| Se1 | S. eubayanus | yHCT69 | South America (Argentina) | Patagonia B/HOL |
| Se2 | S. eubayanus | yHRVM107 | USA (North Carolina) | Patagonia B/HOL |
| Su1 | S. uvarum | yHAB60 | Unknown | HOL/SA-A |
| Su2 | S. uvarum | CBS7001 | Europe (Spain) | HOL-EU |
| Su3 | S. uvarum | yHAB521 | South Ameria (Argentina) | SA-B |
| Su4 | S. uvarum | ZP964 | Asia (New Zealand) | Australasia |
| Su5 | S. uvarum | yHCT77 | USA (Oregon) | HOL-NA |
| Systematic Name | Common Name | Brief Description | Human Ortholog |
|---|---|---|---|
| YLR220W | CCC1 | Vacuolar Fe2+/Mn2+ transporter | - |
| YER174C | GRX4 | Glutathione-dependent oxidoreductase and glutathione S-transferase | GLRX3 |
| YPL207W | TYW1 | Iron-sulfur protein required for synthesis of wybutosine modified tRNA | TYW1A and TYW1B |
| YMR319C | FET4 | Plasma membrane low-affinity Fe2+ transporter | - |
| YGR209C | TRX2 | Cytoplasmic thioredoxin isoenzyme | TXN |
| YML007W | YAP1 | Basic leucine zipper (bZIP) transcription factor | |
| YJR104C | SOD1 | Cytosolic copper-zinc superoxide dismutase | SOD1 |
| YML028W | TSA1 | Thioredoxin peroxidase | PRDX1, PRDX2, PRDX3 and PRDX4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sorribes-Dauden, R.; Jordá, T.; Peris, D.; Martínez-Pastor, M.T.; Puig, S. Adaptation of Saccharomyces Species to High-Iron Conditions. Int. J. Mol. Sci. 2022, 23, 13965. https://doi.org/10.3390/ijms232213965
Sorribes-Dauden R, Jordá T, Peris D, Martínez-Pastor MT, Puig S. Adaptation of Saccharomyces Species to High-Iron Conditions. International Journal of Molecular Sciences. 2022; 23(22):13965. https://doi.org/10.3390/ijms232213965
Chicago/Turabian StyleSorribes-Dauden, Raquel, Tania Jordá, David Peris, María Teresa Martínez-Pastor, and Sergi Puig. 2022. "Adaptation of Saccharomyces Species to High-Iron Conditions" International Journal of Molecular Sciences 23, no. 22: 13965. https://doi.org/10.3390/ijms232213965
APA StyleSorribes-Dauden, R., Jordá, T., Peris, D., Martínez-Pastor, M. T., & Puig, S. (2022). Adaptation of Saccharomyces Species to High-Iron Conditions. International Journal of Molecular Sciences, 23(22), 13965. https://doi.org/10.3390/ijms232213965